Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
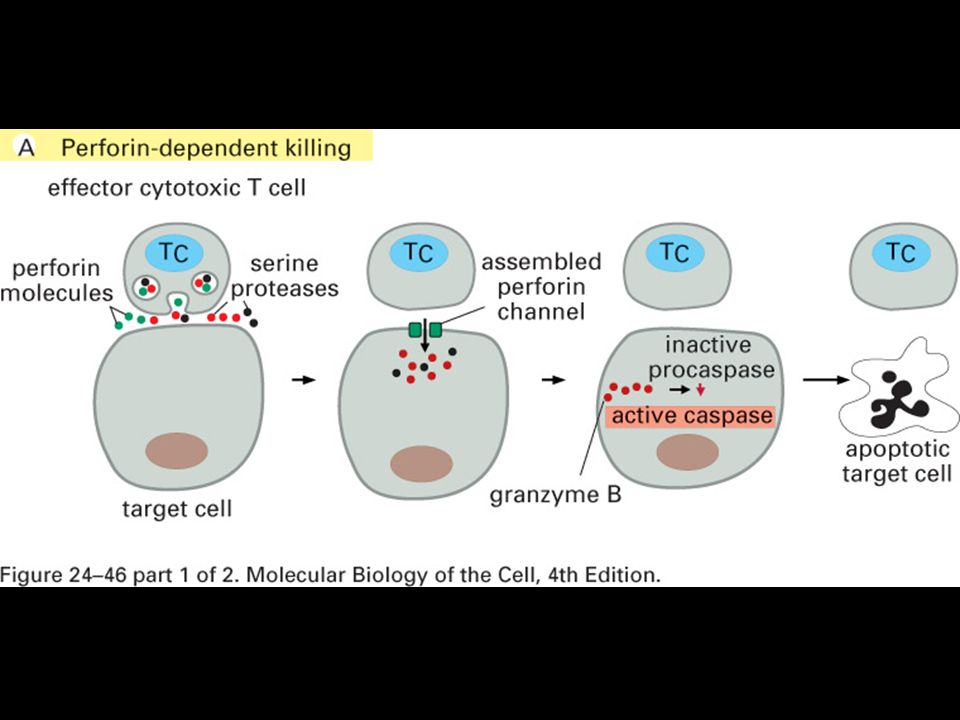
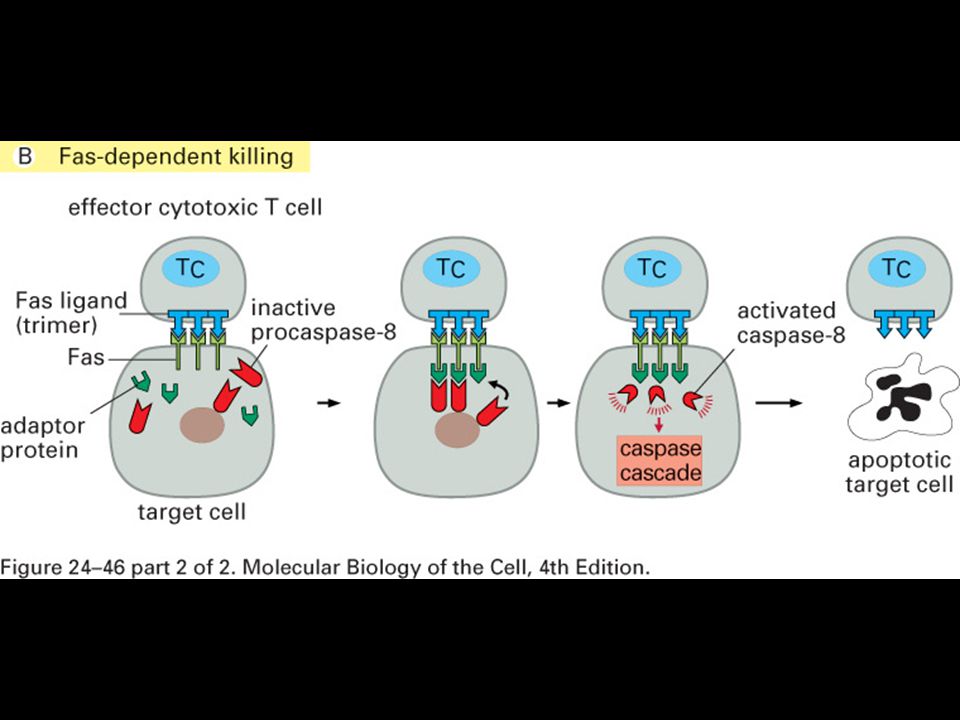
Makrofág a apoptotická buňka
Imunologie Makrofág a apoptotická buňka

2
Život je neustálý boj s parazity
Život je neustálý boj s parazity. Tento věčný boj může za mnohé i za vnímání krásna a různé podivnosti s ním spojené, m.j spojené s pohlavním rozmnožováním

3
Humorální a buněčná imunita

5
Makrofág a baktérie

6
A macrophage extends a semiliquid projection (a so-called pseudopodium) towards some bacteria in the foreground.
 towards some bacteria in the foreground.")
7
A macrophage extends a long, thin projection towards a foreign protein body.

8
Macrophages move more slowly than granulocytes but, on the other hand, are tougher and can handle larger prey. They are the armoured tanks of the immune system. Here they are in the process of enclosing microscopic glass fibres, which are also indigestible and can only be captured and transported away.

9
The Wars Within. Washington D. C. : National Geographic Society
The Wars Within. Washington D.C.: National Geographic Society. bijschrift: One of mankind's greatest inorganic threats, asbestos fibers (left) are engulfed by a macrophage, which will probably die from its indigestible meal.
 are engulfed by a macrophage, which will probably die from its indigestible meal.")
10
Infikovaný pomocný T-lymfocyt virem HIV

11
Adaptivní (specifická imunitní odpověď)
")
12
Vrozená (nespecifická) imunitní odpověď
 imunitní odpověď")
14
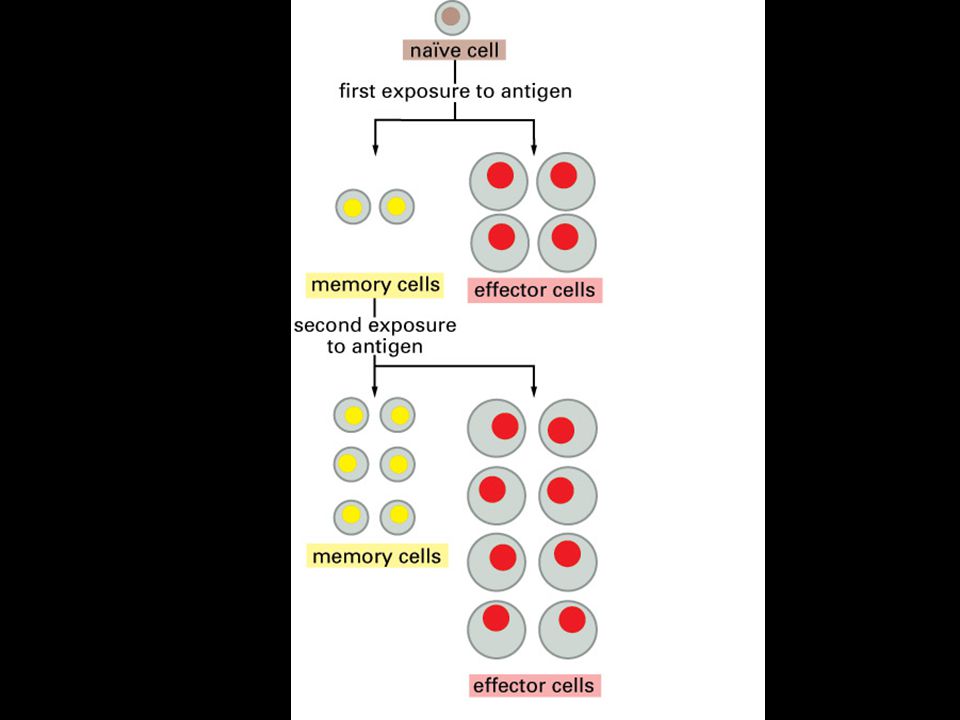
Imunologická paměť

16
Vrozená a adaptivní odpovědi fungují paralelně a kooperují

18
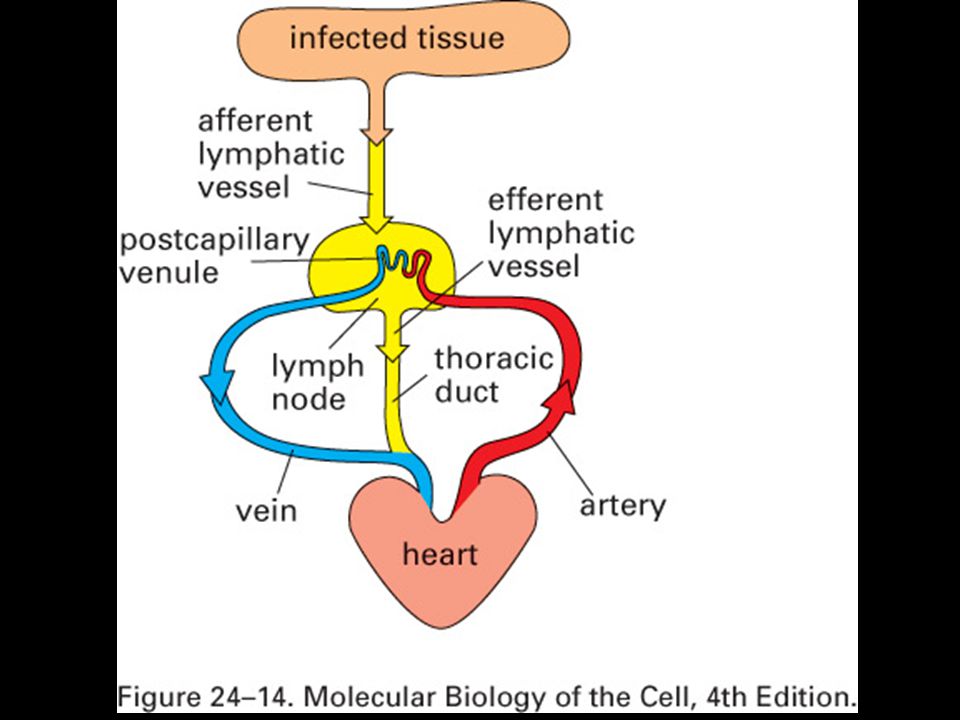
Primární (žluté) a sekundární (modré) imunologické orgány
 a sekundární (modré) imunologické orgány")
21
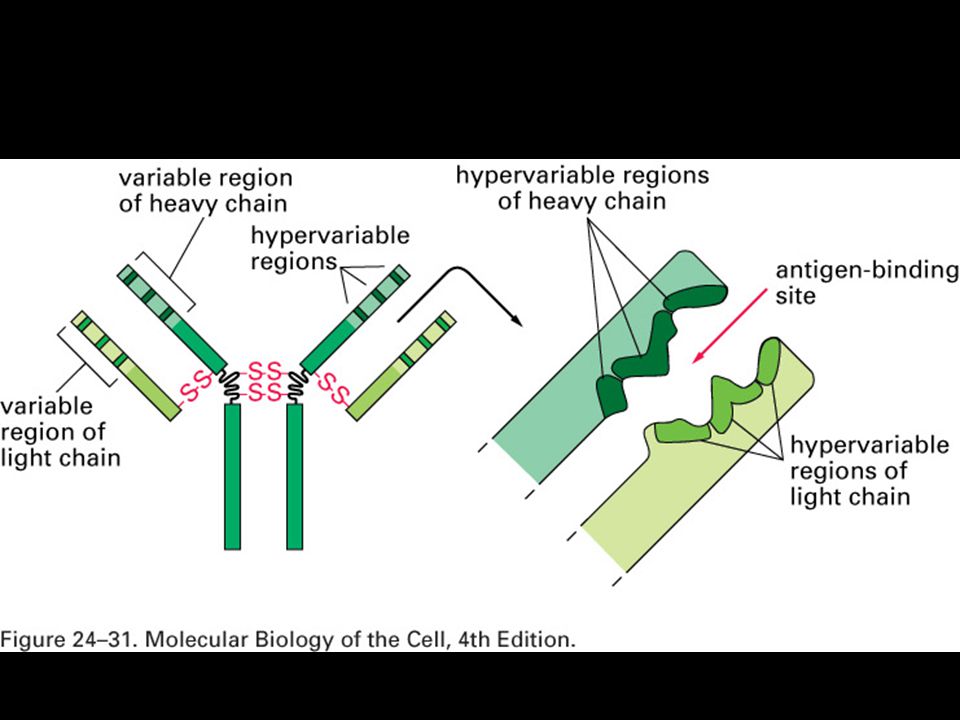
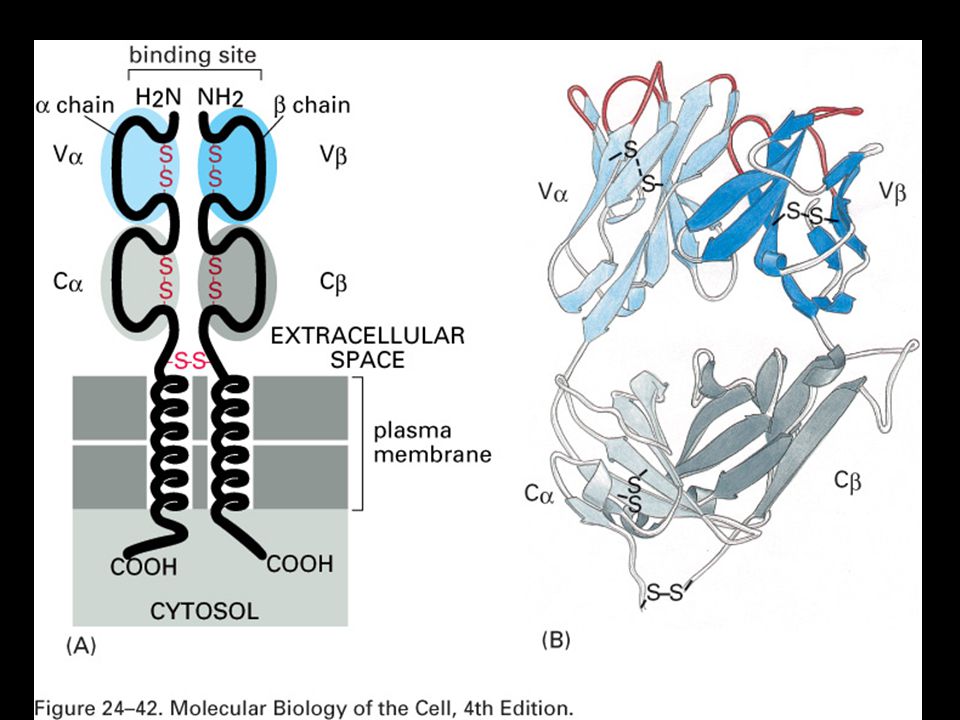
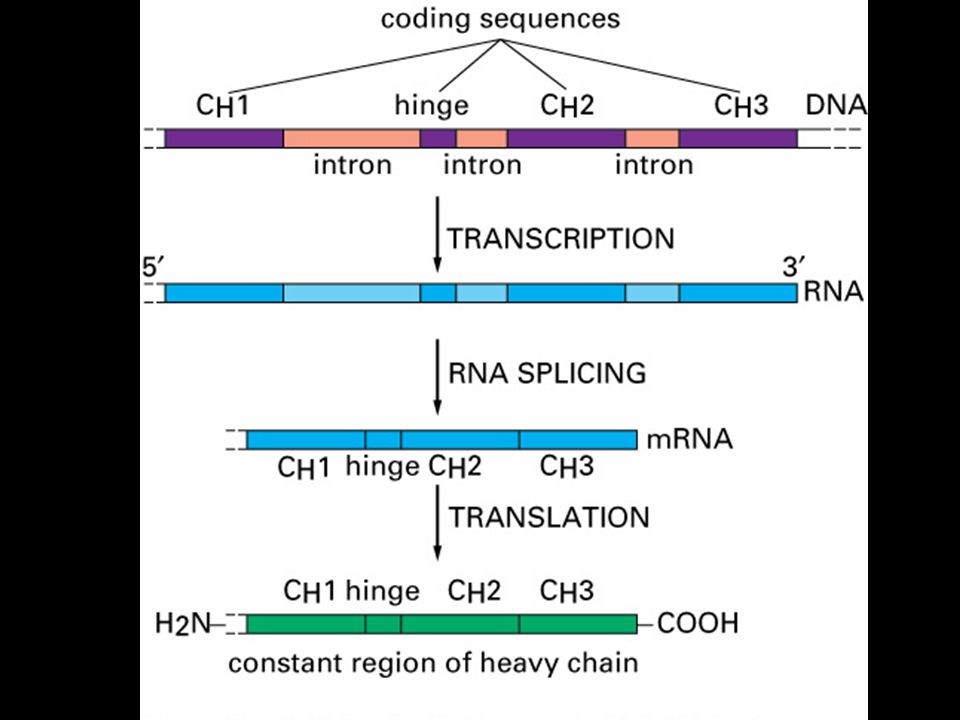
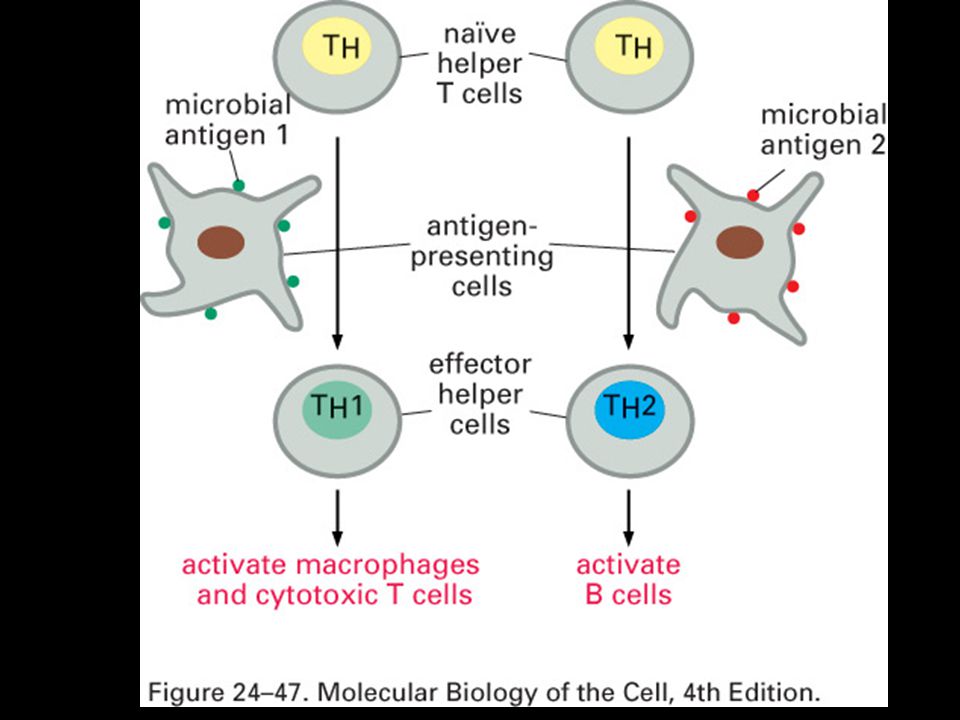
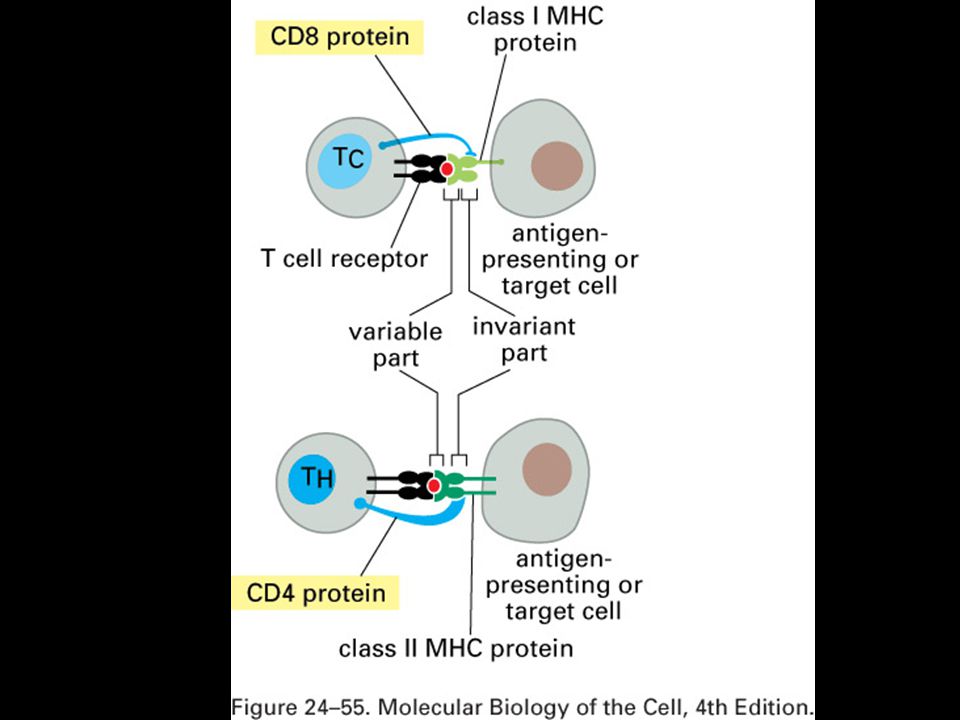
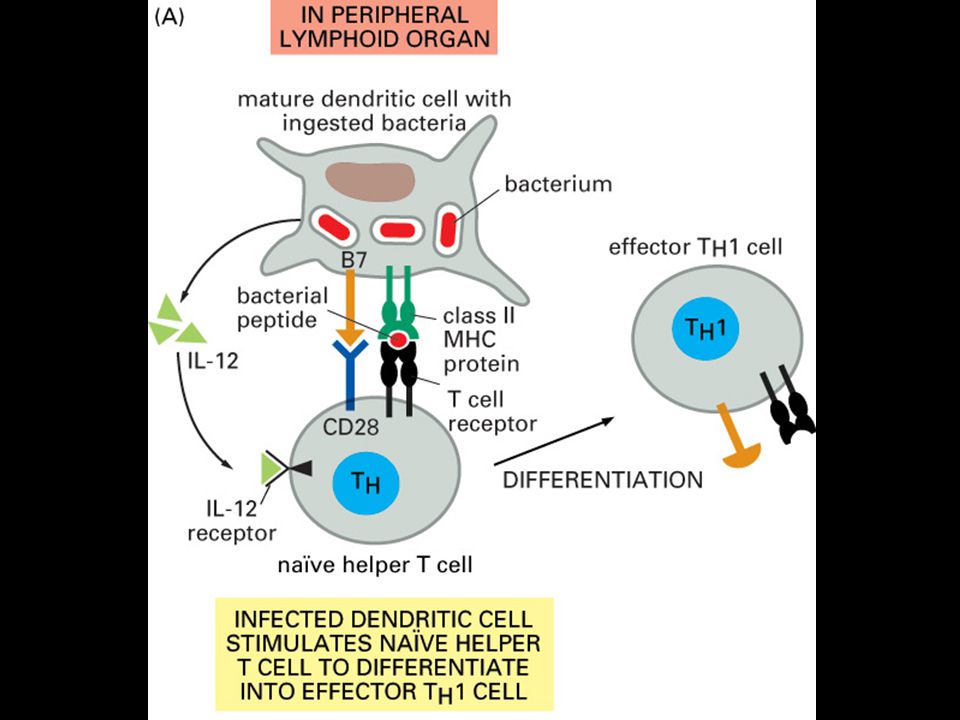
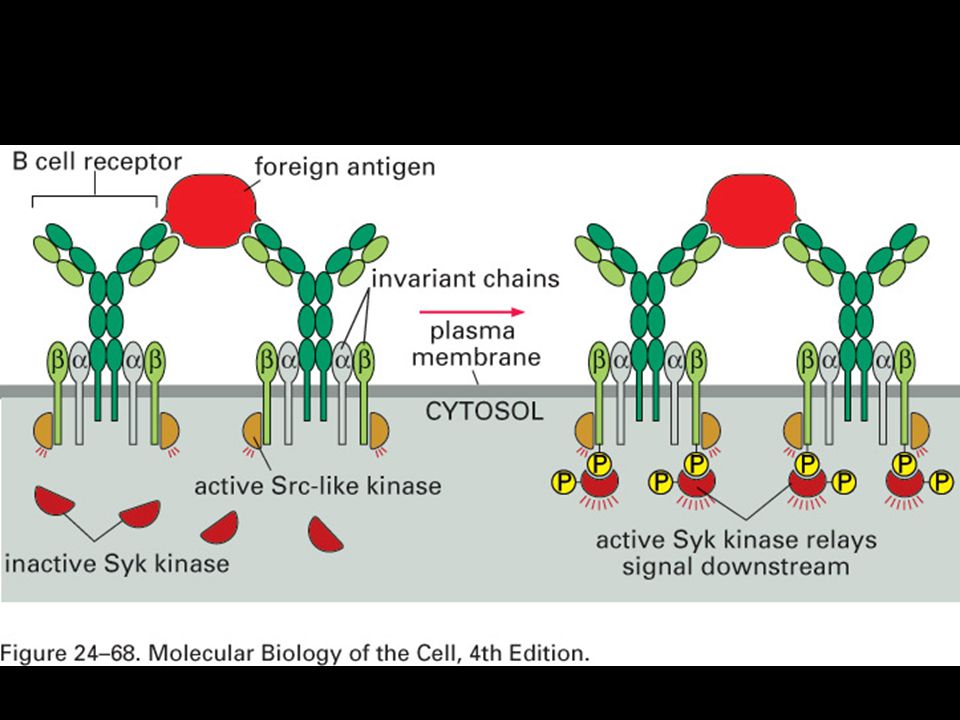
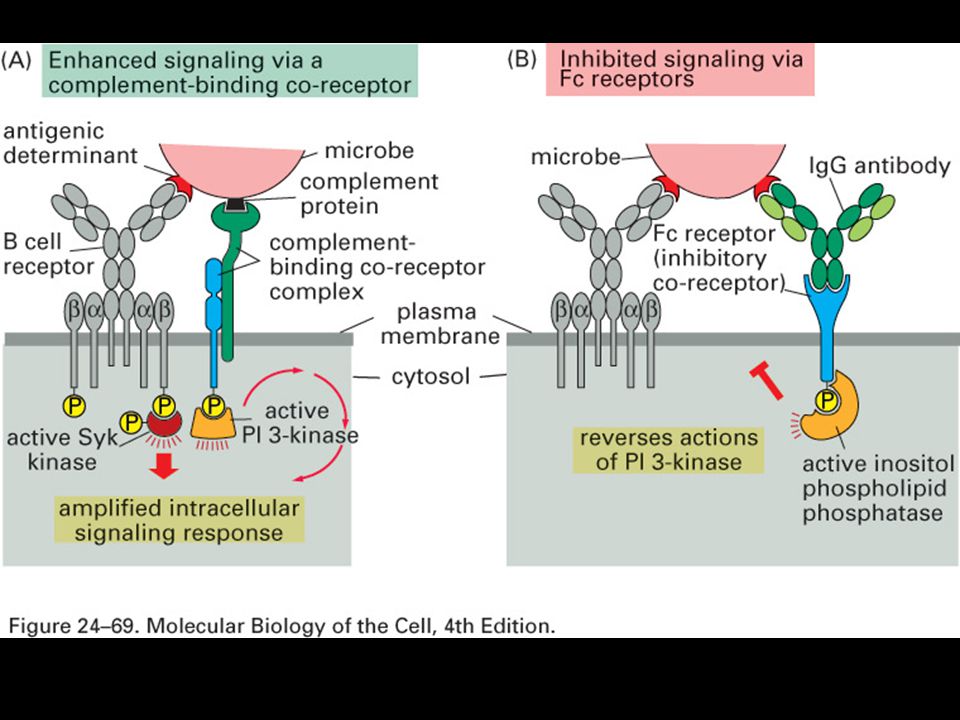
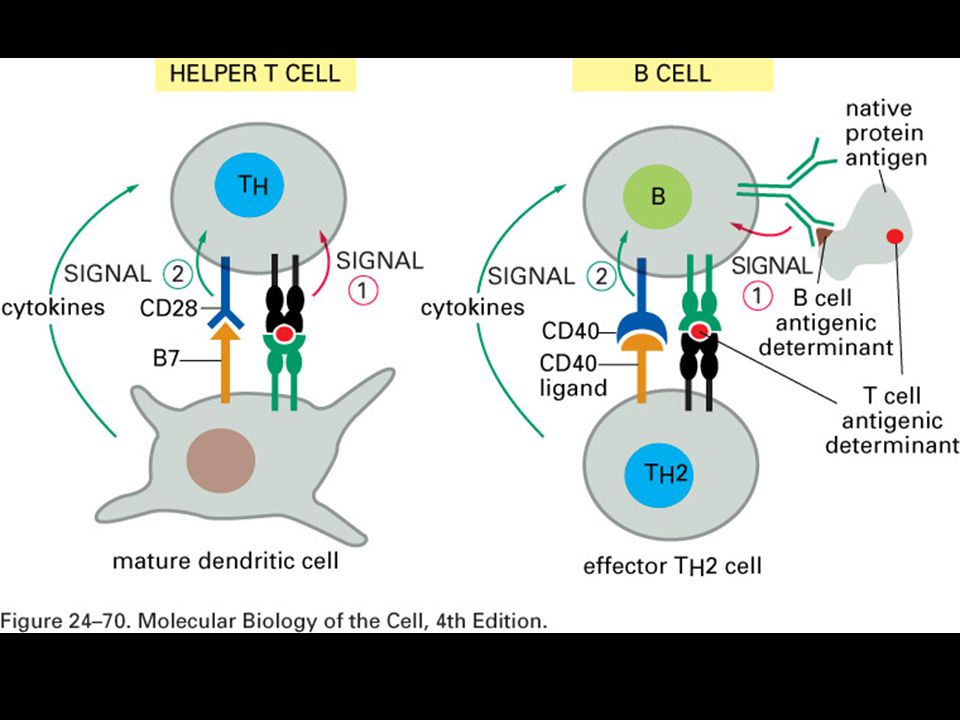
Specifická imunitní odpověď je buněčného původu

22
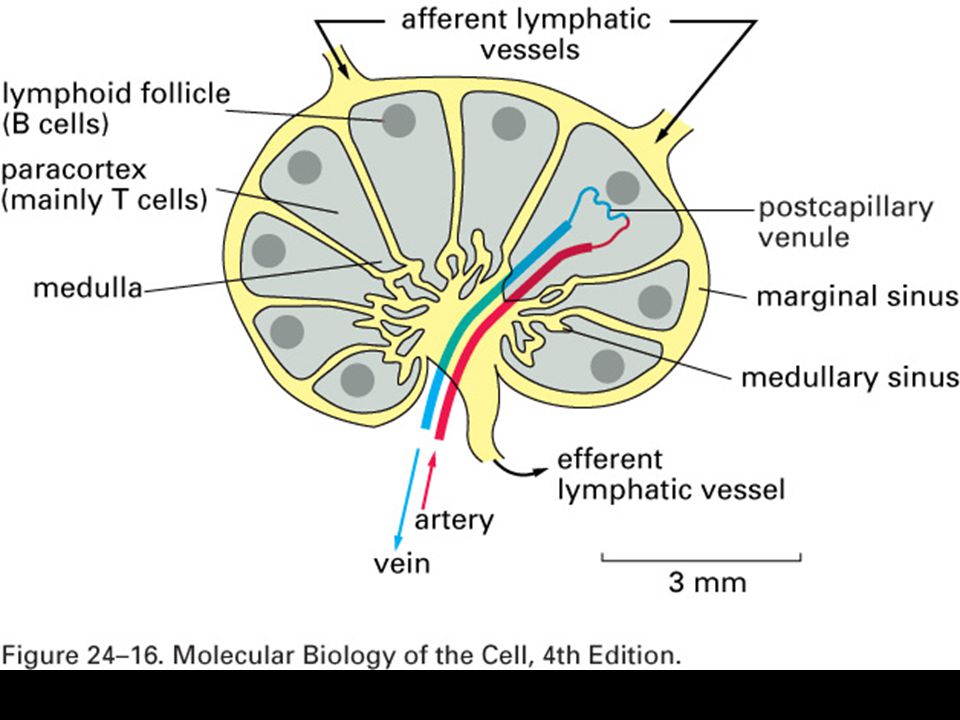
Role primárních a sekundárních imunologických orgánů

23
Buňky imunitního systému jsou schopné morgologické a funkční diferenciace

27
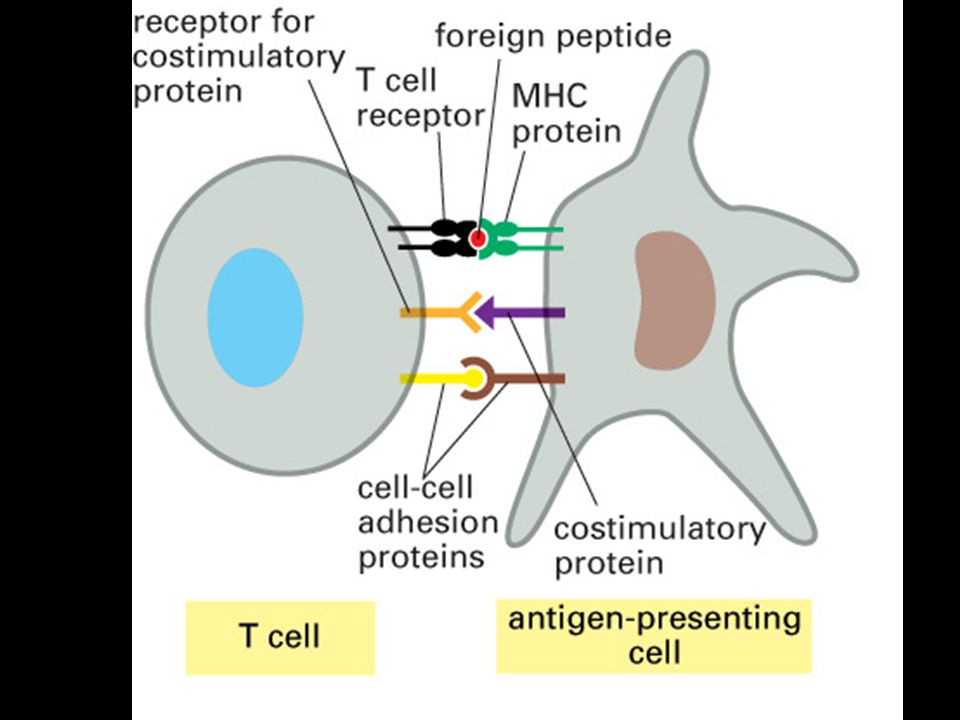
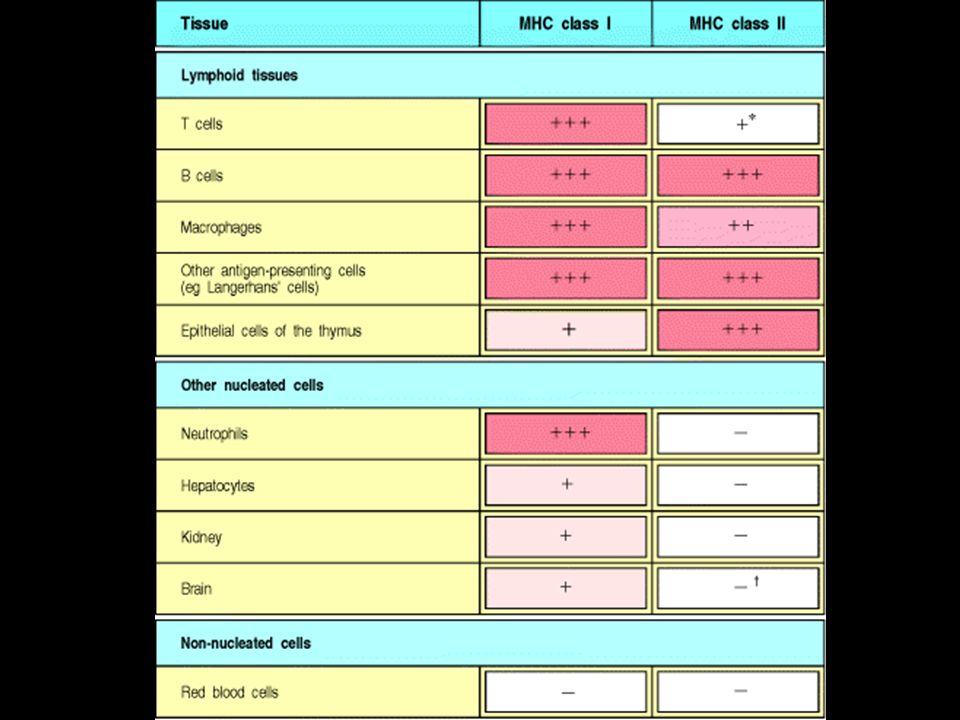
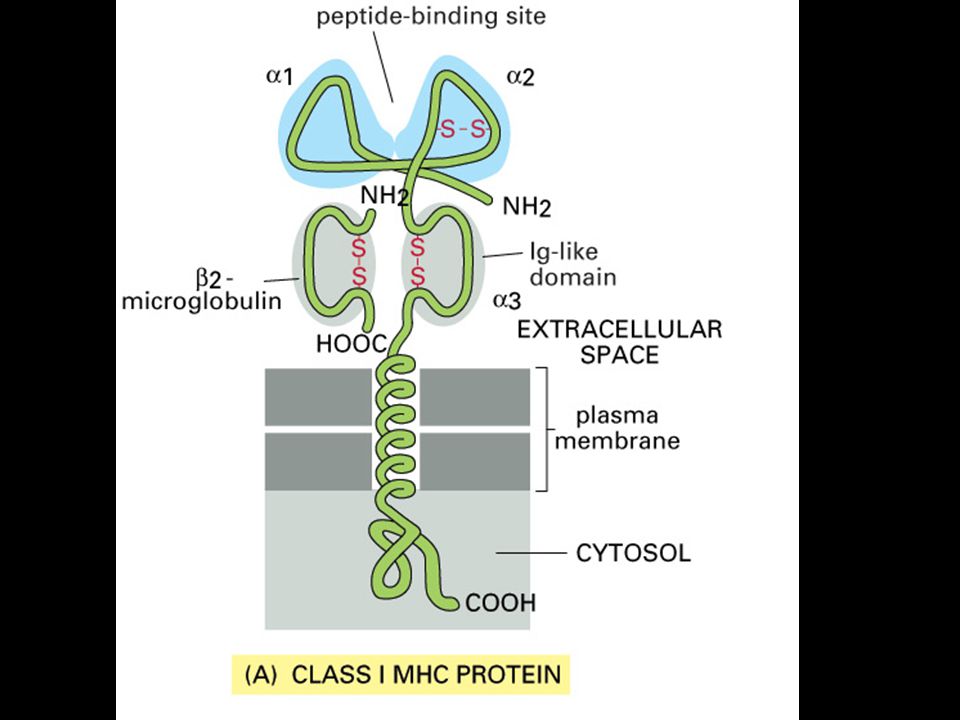
Antigenní prezentace a MHC glykoproteiny

28
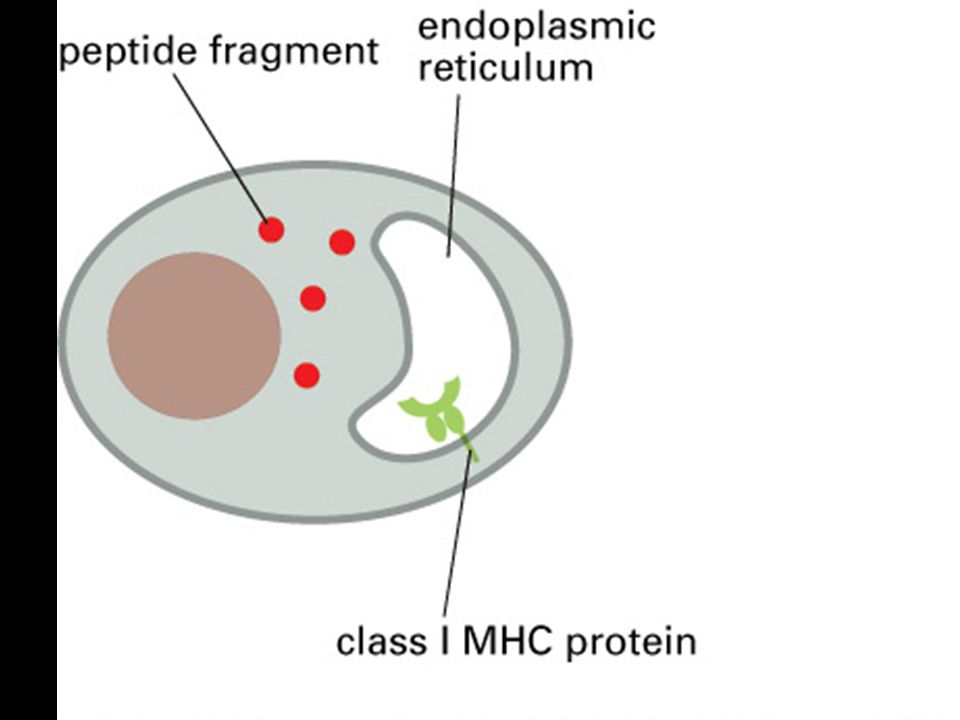
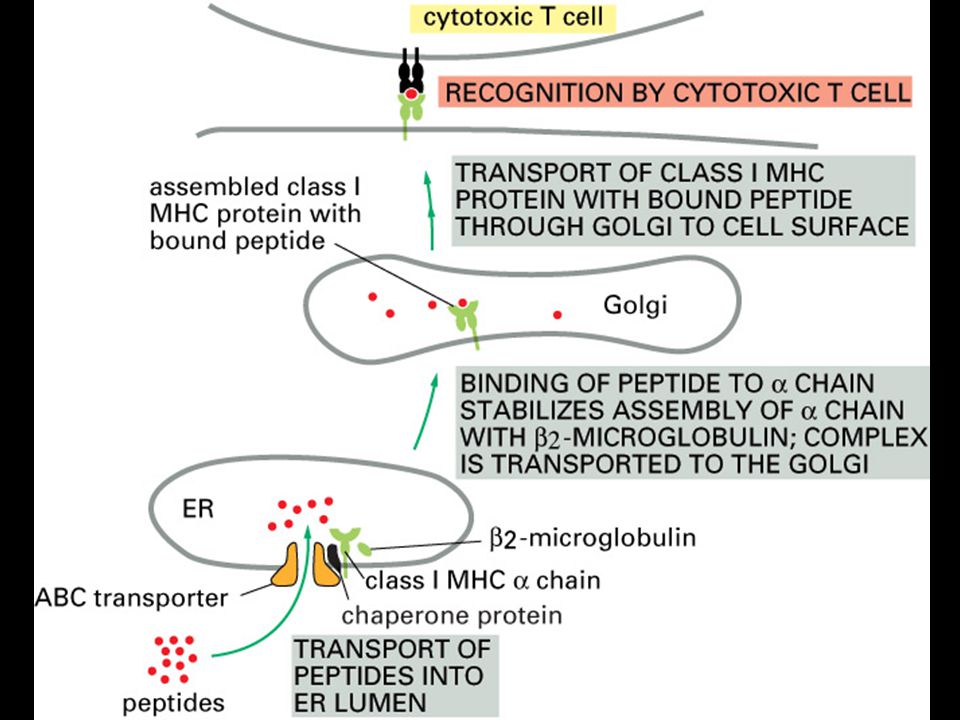
MHC class I molecules do not leave the endoplasmic reticulum unless they bind peptides.
Peptides generated by degradation of proteins in the cytoplasm are transported into the lumen of the ER. MHC class I a chains assemble in the ER with a membrane-bound protein, calnexin (Cx). When this complex binds b2-microglobulin (b2m) it is released from calnexin and the partially folded MHC class I molecule then binds to the TAP-1 subunit of the TAP transporter by interacting with one molecule of the TAP-associated protein tapasin, and a chaperonin, calreticulin. It is retained in the ER until released by binding of a peptide, completing the folding of the MHC class I molecule. The peptide:MHC complex is then transported through the Golgi complex to the cell surface. The source of peptides for MHC class I molecules is the degradation of proteins in the cytosol by a large protease complex, the proteasome. The proteasome contains 28 subunits arranged to form a cylindrical structure composed of four rings, each of seven subunits. It is not known exactly how the mammalian proteasome degrades cytosolic proteins. It has six active proteolytic sites, which correspond to the N-terminal threonine residues of three of the seven b subunits, and are found in the center of the cylinder. Thus, it is likely that proteins have to unfold and pass through the center of the cylindrical structure as shown here for degradation to occur. Peptide fragments generated by the proteasome are transported into the lumen of the endoplasmic reticulum (ER) by the TAP transporter. There, the peptides bind to the complex of the partially folded MHC molecule, liberating the MHC class I molecule from the TAP/tapasin complex, thus allowing the peptide:MHC complex to be delivered to the cell surface.
. When this complex binds b2-microglobulin (b2m) it is released from calnexin and the partially folded MHC class I molecule then binds to the TAP-1 subunit of the TAP transporter by interacting with one molecule of the TAP-associated protein tapasin, and a chaperonin, calreticulin. It is retained in the ER until released by binding of a peptide, completing the folding of the MHC class I molecule. The peptide:MHC complex is then transported through the Golgi complex to the cell surface. The source of peptides for MHC class I molecules is the degradation of proteins in the cytosol by a large protease complex, the proteasome. The proteasome contains 28 subunits arranged to form a cylindrical structure composed of four rings, each of seven subunits. It is not known exactly how the mammalian proteasome degrades cytosolic proteins. It has six active proteolytic sites, which correspond to the N-terminal threonine residues of three of the seven b subunits, and are found in the center of the cylinder. Thus, it is likely that proteins have to unfold and pass through the center of the cylindrical structure as shown here for degradation to occur. Peptide fragments generated by the proteasome are transported into the lumen of the endoplasmic reticulum (ER) by the TAP transporter. There, the peptides bind to the complex of the partially folded MHC molecule, liberating the MHC class I molecule from the TAP/tapasin complex, thus allowing the peptide:MHC complex to be delivered to the cell surface.")
30
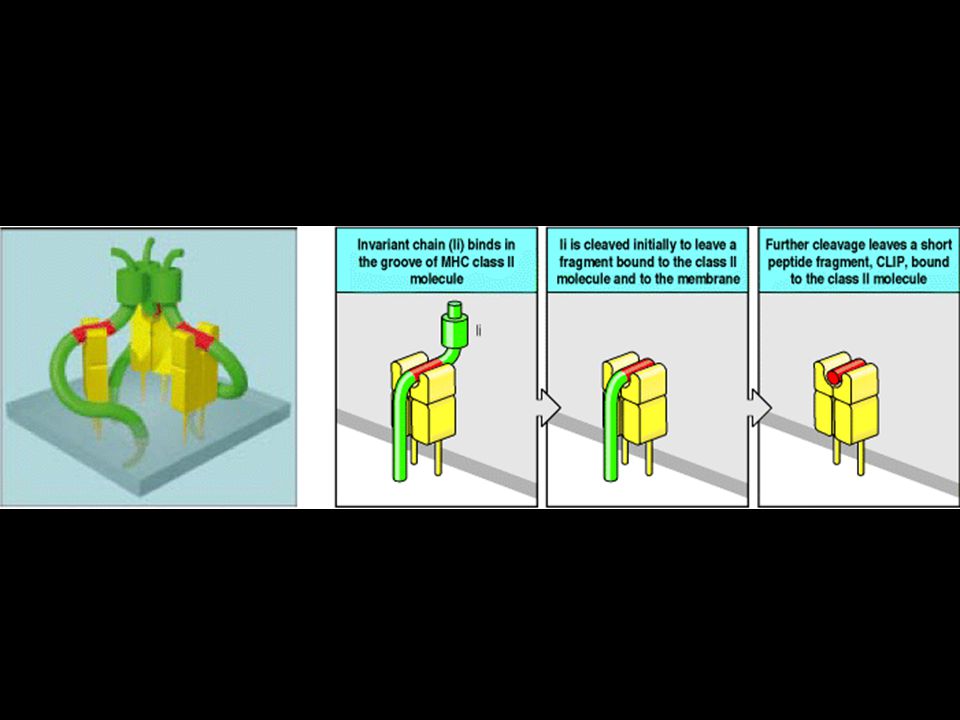
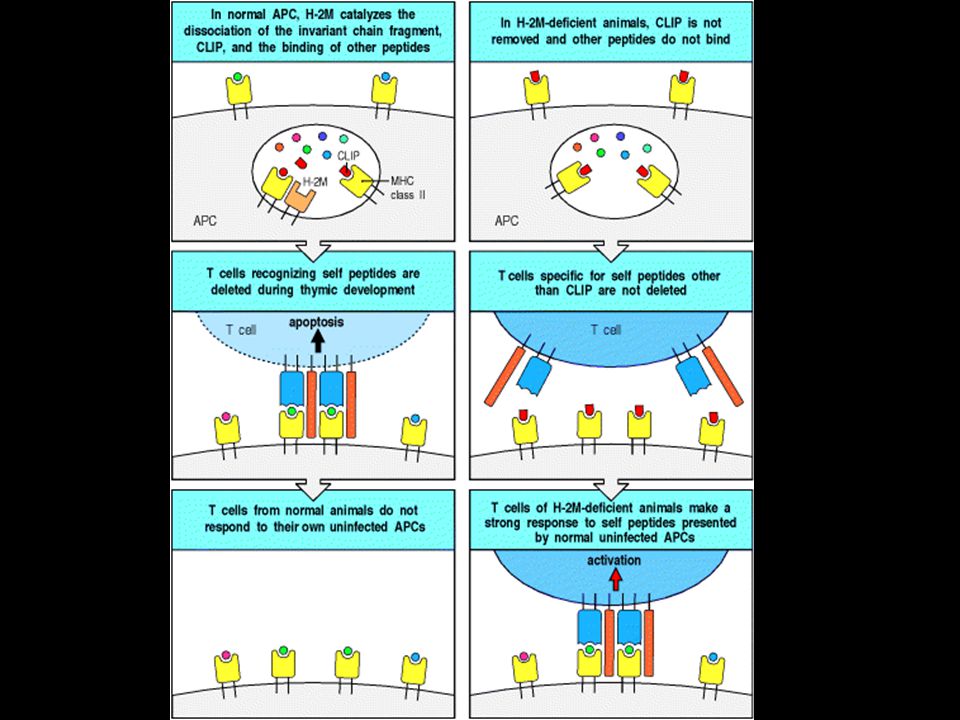
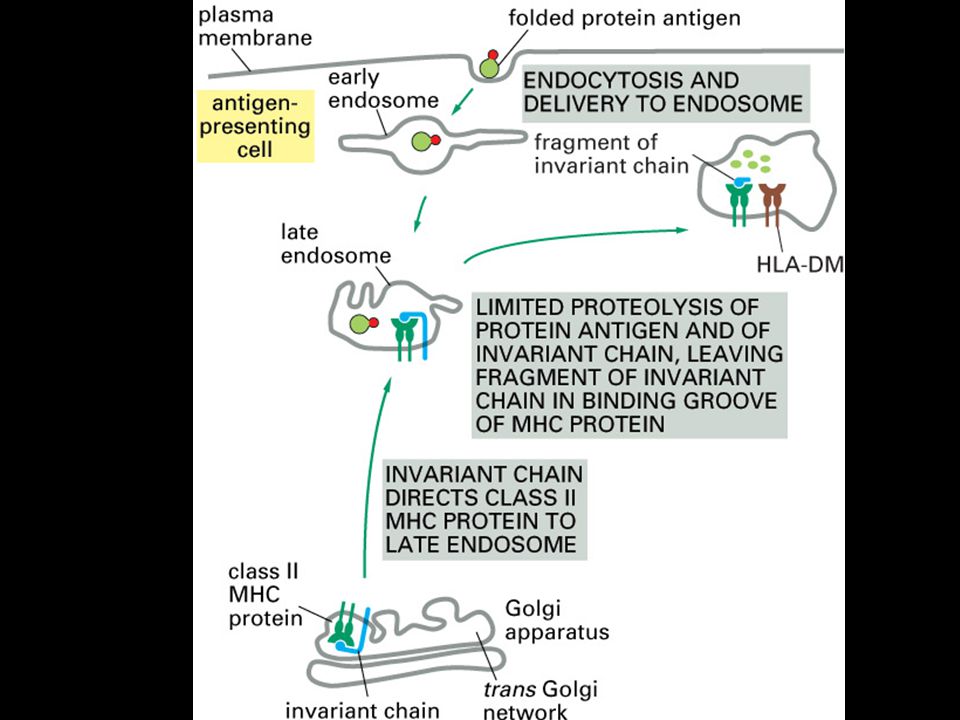
Peptides that bind to MHC class II molecules are generated in acidified endocytic vesicles. HLA-DM facilitates the loading of antigenic peptides onto class II MHC molecules. The invariant chain (Ii) assembles with newly synthesized MHC class II molecules in the ER where it prevents the MHC class II molecule from binding intracellular peptides and partially folded proteins present in the lumen, and directs its export through the Golgi apparatus to acidified endosomes containing peptides of resident bacteria or engulfed extracellular proteins. Here the invariant chain is cleaved in stages, and the MHC class II molecule binds antigenic peptide and is transported to the cell surface. A model for the trimeric invariant chain bound to ab heterodimers is also shown. The CLIP portion is shown in red, the rest of Ii is in green, and the ab MHC class II heterodimer is in yellow.
 assembles with newly synthesized MHC class II molecules in the ER where it prevents the MHC class II molecule from binding intracellular peptides and partially folded proteins present in the lumen, and directs its export through the Golgi apparatus to acidified endosomes containing peptides of resident bacteria or engulfed extracellular proteins. Here the invariant chain is cleaved in stages, and the MHC class II molecule binds antigenic peptide and is transported to the cell surface. A model for the trimeric invariant chain bound to ab heterodimers is also shown. The CLIP portion is shown in red, the rest of Ii is in green, and the ab MHC class II heterodimer is in yellow.")
69
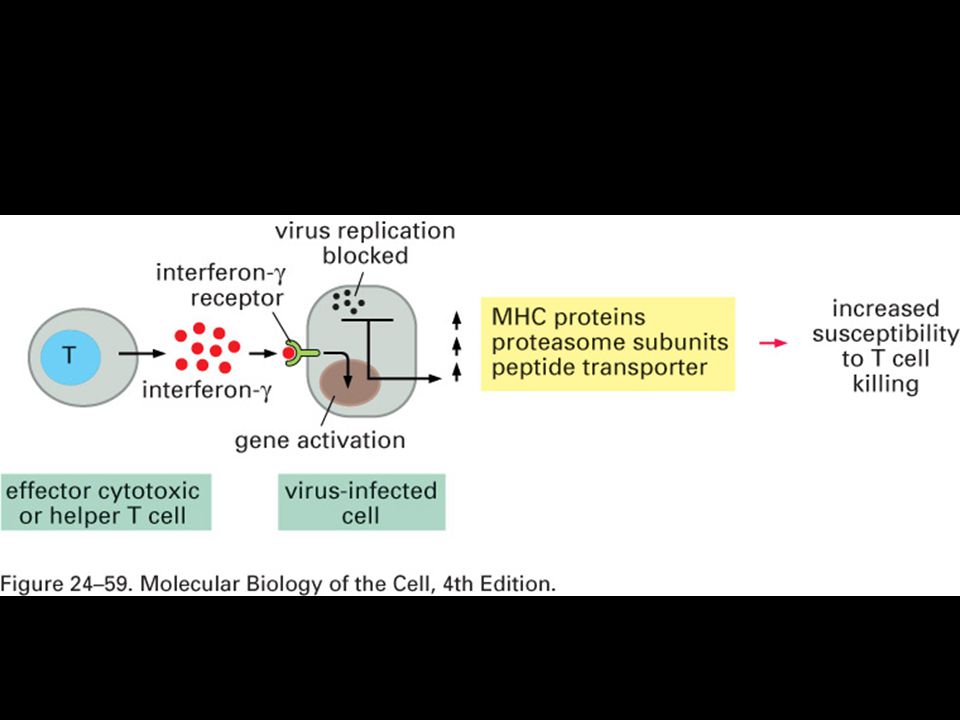
There are two major compartments within cells, separated by membranes.
Pathogens and their products can be found in either the cytoplasmic or the vesicular compartment of cells.

71
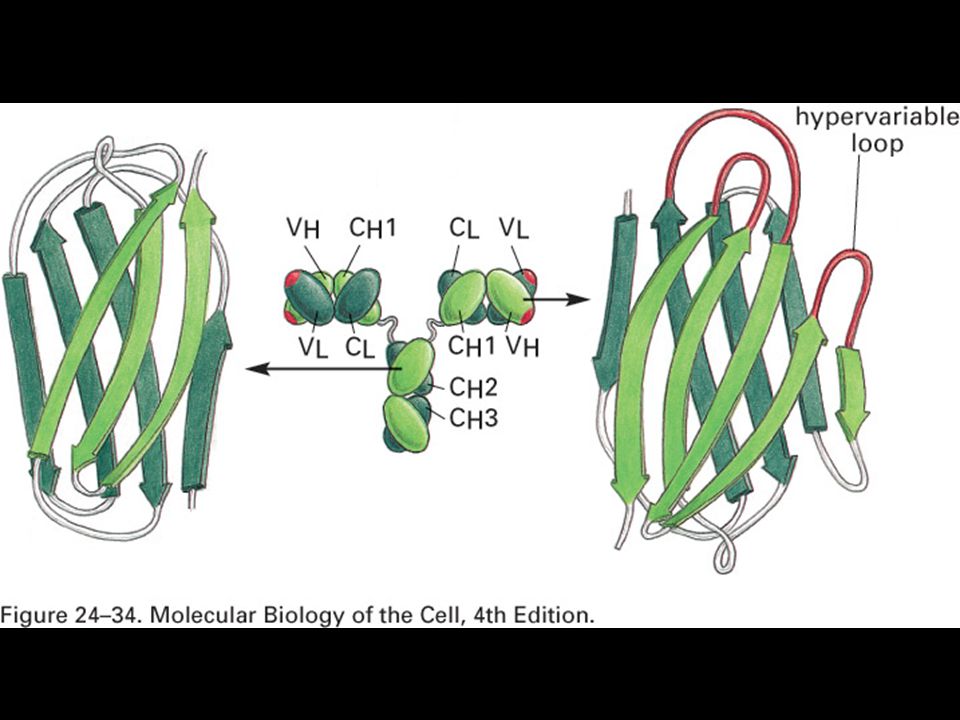
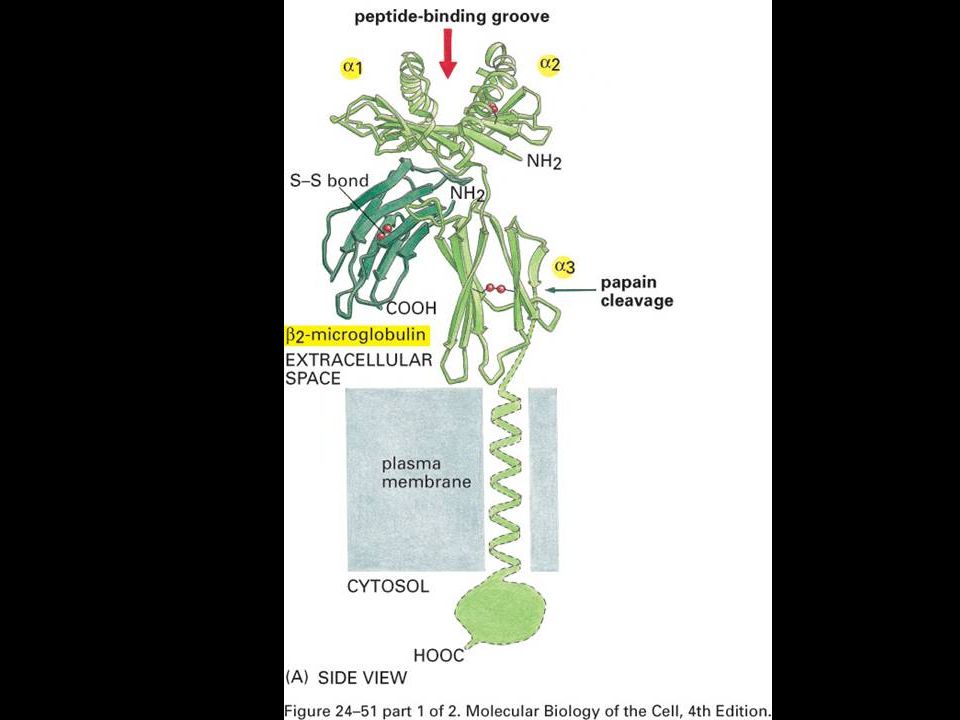
The structure of an MHC class I molecule, determined by X-ray crystallography.
Panel a shows a computer graphic representation of a human MHC class I molecule, HLA-A2, which has been cleaved from the cell surface by the enzyme papain. Panel b shows a ribbon diagram of that structure. Shown schematically in panel d, the MHC class I molecule is a heterodimer of a membrane-spanning a chain (43 kD), non-covalently associated with b2-microglobulin (b2m, 12 kD), which does not span the membrane. The a chain folds into three domains, a1, a2, and a3. The a3 domain and b2m show similarities in amino acid sequence to immunoglobulin constant domains and have a similar folded structure, while the a1 and a2 domains fold together into a single structure consisting of two segmented a helices lying on a sheet of eight antiparallel b-strands. The folding of the a1 and a2 domains creates a long cleft or groove, which is the site at which peptide antigens bind to the MHC molecules. The transmembrane region and the short stretch of peptide that connects the external domains to the cell surface are not seen in panels a and b as they have been removed by the papain digestion. As can be seen in panel c, looking down on the molecule from above, the sides of the cleft are formed from the inner faces of the two a helices, while the b-pleated sheet formed by the pairing of the a1 and a2 domains creates the floor of the cleft.
, non-covalently associated with b2-microglobulin (b2m, 12 kD), which does not span the membrane. The a chain folds into three domains, a1, a2, and a3. The a3 domain and b2m show similarities in amino acid sequence to immunoglobulin constant domains and have a similar folded structure, while the a1 and a2 domains fold together into a single structure consisting of two segmented a helices lying on a sheet of eight antiparallel b-strands. The folding of the a1 and a2 domains creates a long cleft or groove, which is the site at which peptide antigens bind to the MHC molecules. The transmembrane region and the short stretch of peptide that connects the external domains to the cell surface are not seen in panels a and b as they have been removed by the papain digestion. As can be seen in panel c, looking down on the molecule from above, the sides of the cleft are formed from the inner faces of the two a helices, while the b-pleated sheet formed by the pairing of the a1 and a2 domains creates the floor of the cleft.")
72
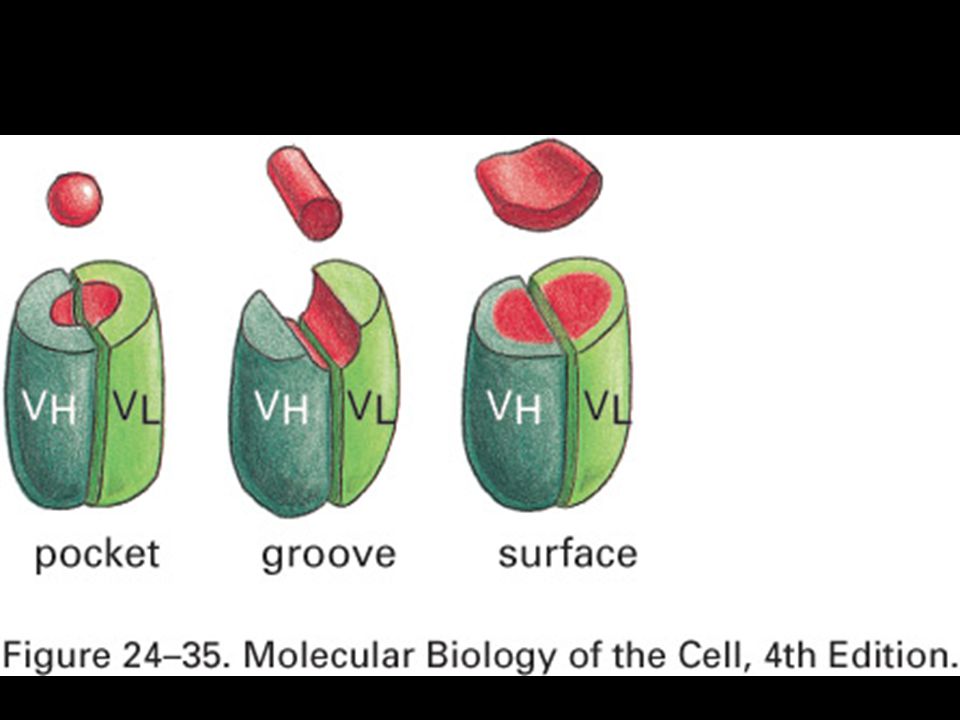
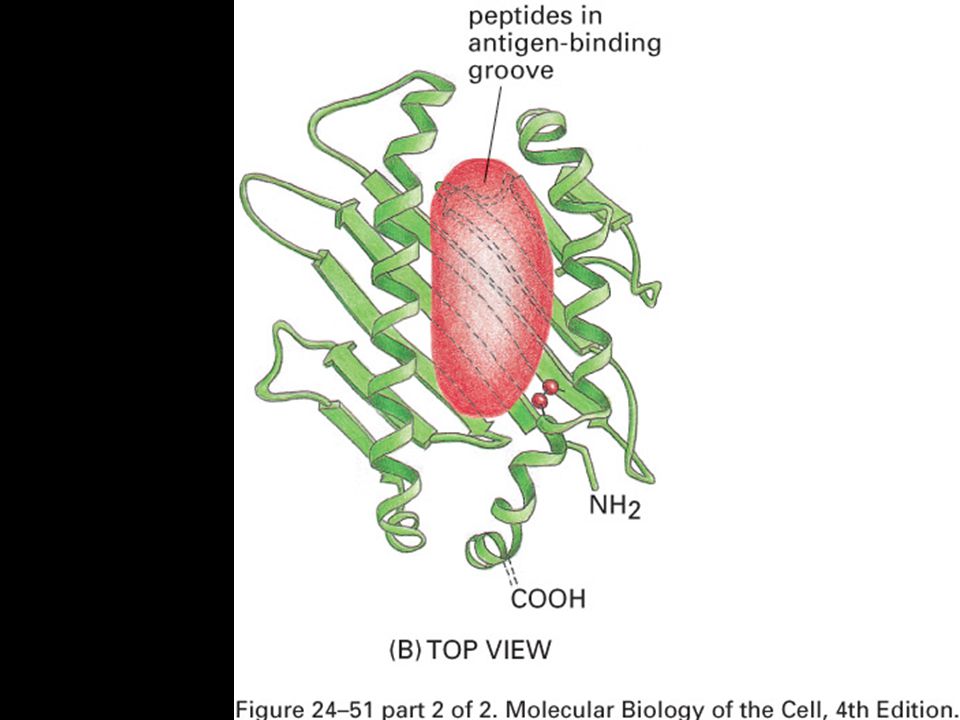
Peptides are bound to MHC class I molecules by their ends.
Peptides bind to MHC molecules through structurally related anchor residues.

73
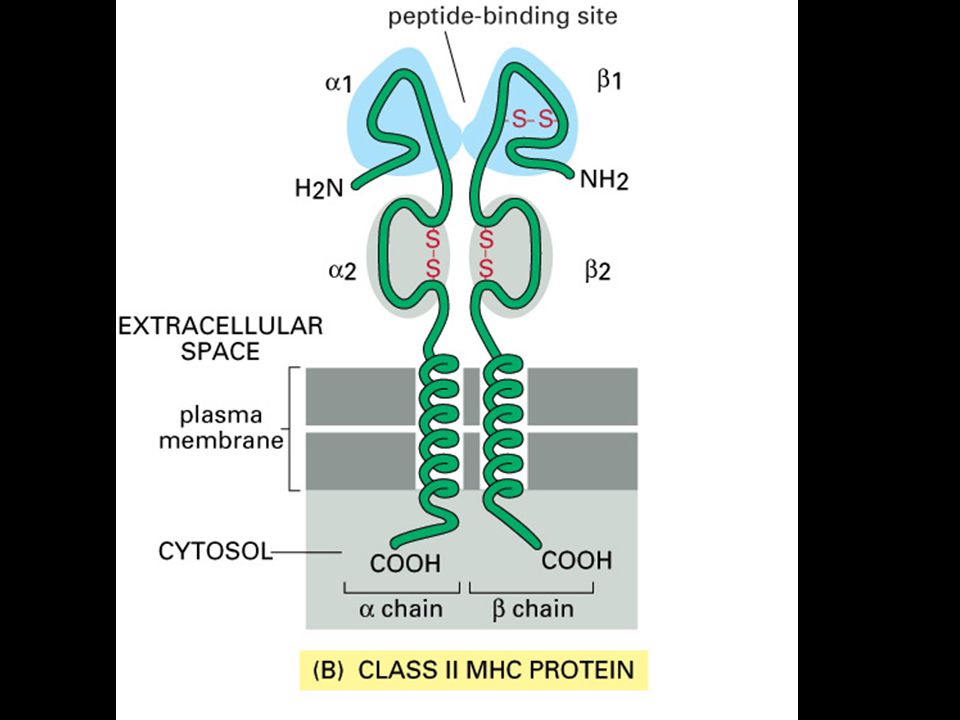
MHC class II molecules resemble MHC class I molecules in structure.
The MHC class II molecule is composed of two transmembrane glycoprotein chains, a ( Da) and b ( Da), as shown schematically in panel d. Each chain has two domains, and the two chains together form a compact four-domain structure similar to that of the class I molecule. Panel a shows a computer graphic representation of the MHC class II molecule, in this case, the human protein HLA-DR1; panel b shows the equivalent ribbon diagram. The a 2 and b2 domains, like the a 3 and b2-microglobulin domains of the MHC class I molecule, have amino acid sequence and structural similarities to immunoglobulin constant domains; in the MHC class II molecule, the two domains forming the peptide-binding cleft are contributed by different chains, and are therefore not joined by a covalent bond (see panels c and d). Another important difference, not apparent in this diagram, is that the peptide-binding groove of the MHC class II molecule is open at both ends. Photograph courtesy of C Thorpe. MHC class II molecules resemble MHC class I molecules in structure.
 and b ( Da), as shown schematically in panel d. Each chain has two domains, and the two chains together form a compact four-domain structure similar to that of the class I molecule. Panel a shows a computer graphic representation of the MHC class II molecule, in this case, the human protein HLA-DR1; panel b shows the equivalent ribbon diagram. The a 2 and b2 domains, like the a 3 and b2-microglobulin domains of the MHC class I molecule, have amino acid sequence and structural similarities to immunoglobulin constant domains; in the MHC class II molecule, the two domains forming the peptide-binding cleft are contributed by different chains, and are therefore not joined by a covalent bond (see panels c and d). Another important difference, not apparent in this diagram, is that the peptide-binding groove of the MHC class II molecule is open at both ends. Photograph courtesy of C Thorpe. MHC class II molecules resemble MHC class I molecules in structure.")
74
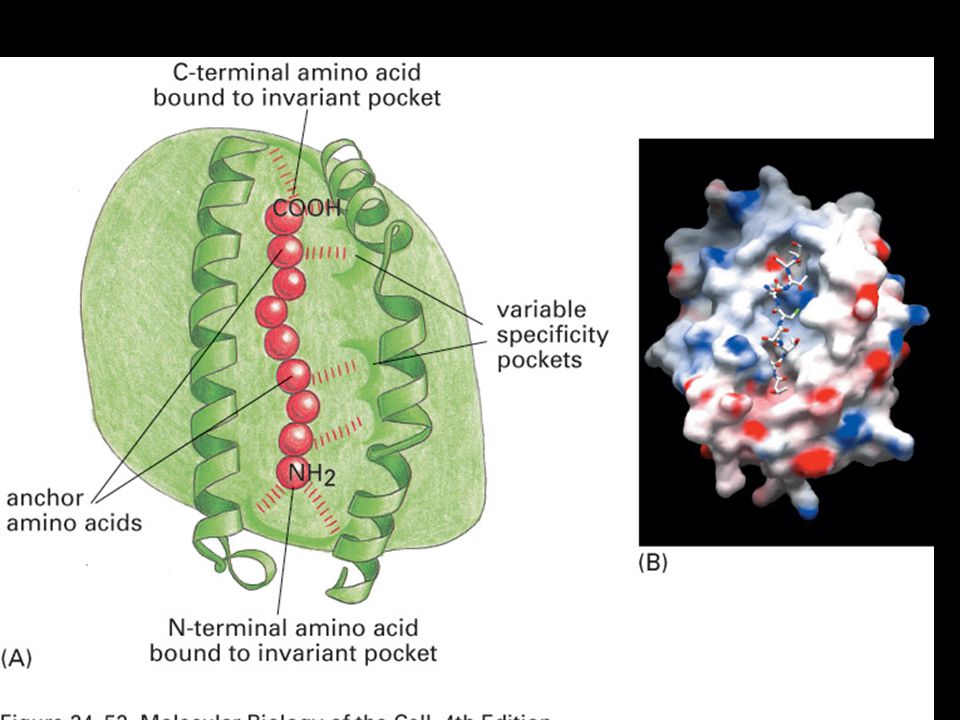
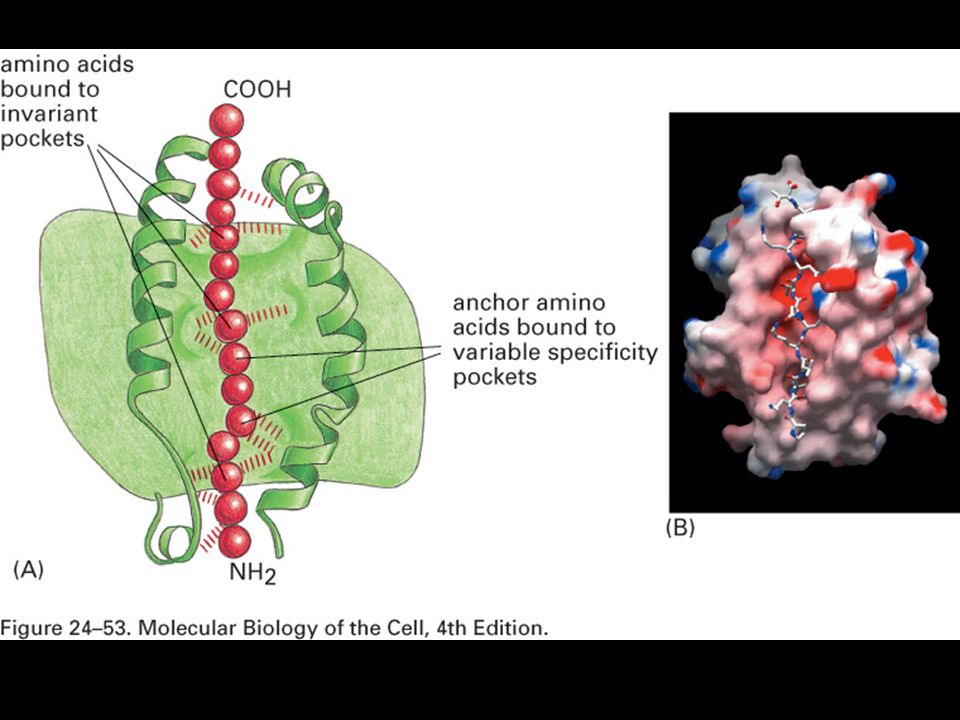
Peptides are bound to MHC class II molecules by interactions along the length of the binding groove.
Peptides that bind MHC class II molecules are variable in length and their anchor residues lie at various distances from the ends of the peptides.

76
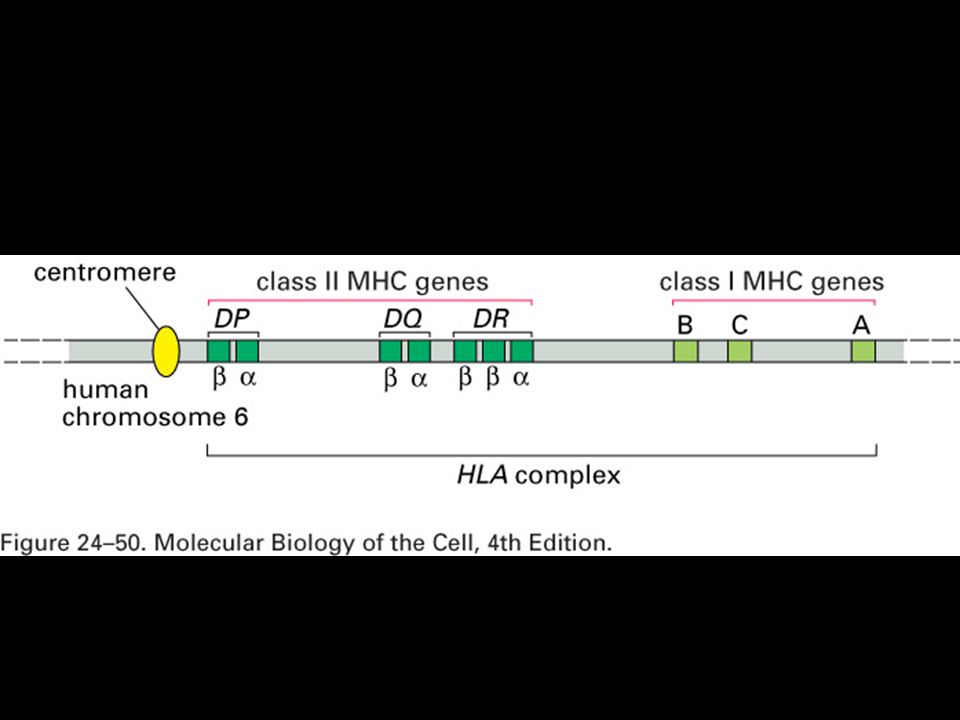
Human MHC genes are highly polymorphic
Human MHC genes are highly polymorphic. In humans MHC genes are referred to as HLA (human leukocyte antigens). Expression of MHC genes is codominant. Polymorphism and polygeny both contribute to the diversity of MHC molecule expression by an individual.
. Expression of MHC genes is codominant. Polymorphism and polygeny both contribute to the diversity of MHC molecule expression by an individual.")
77
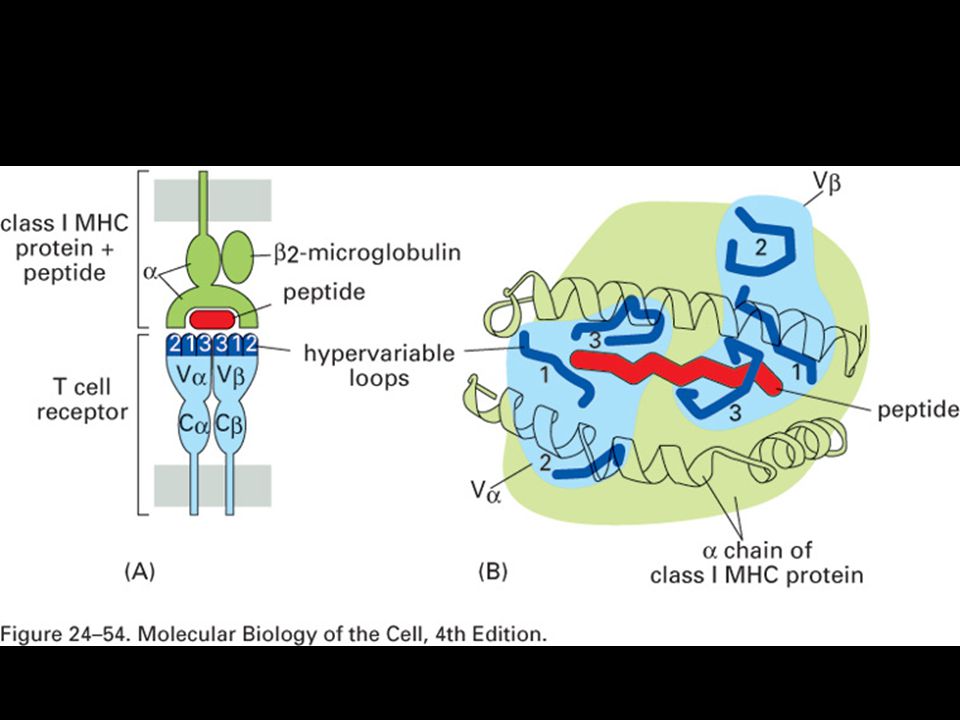
Allelic variation occurs at specific sites within the MHC molecule.
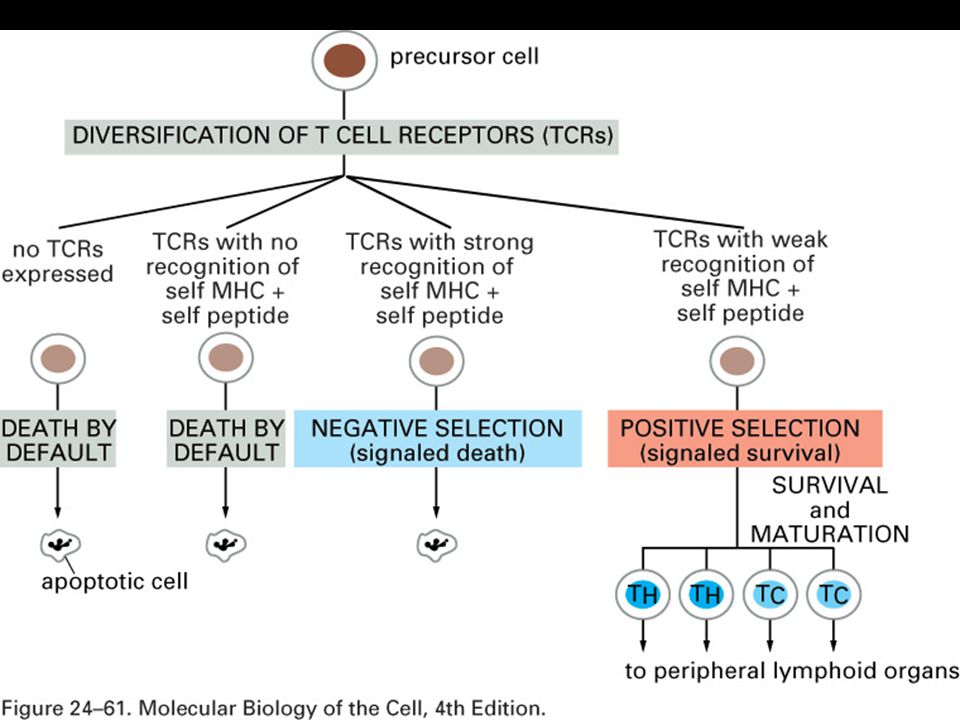
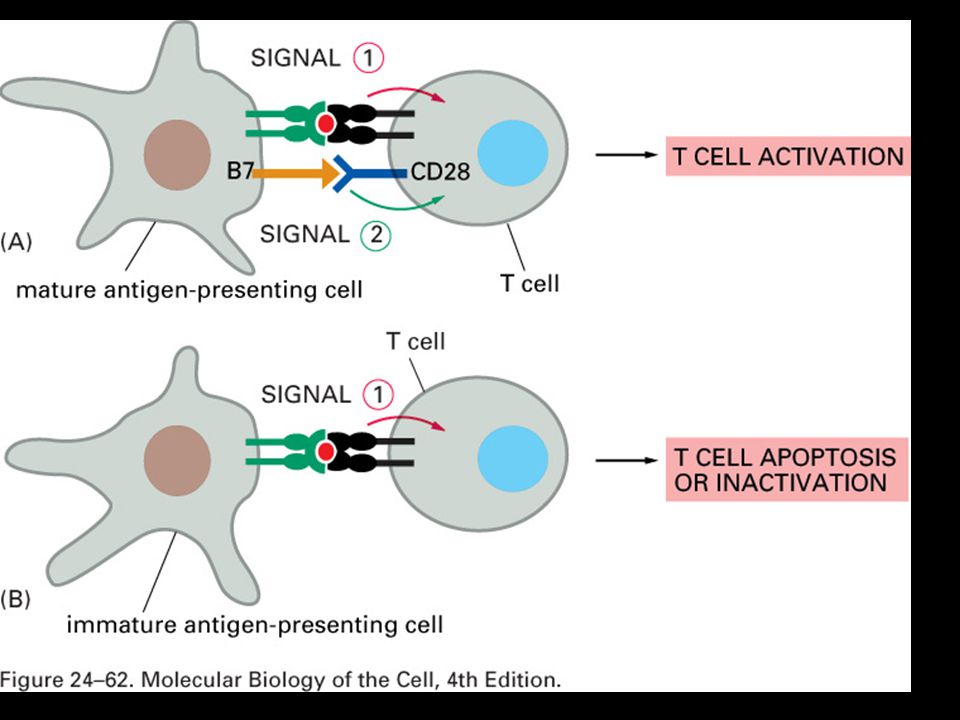
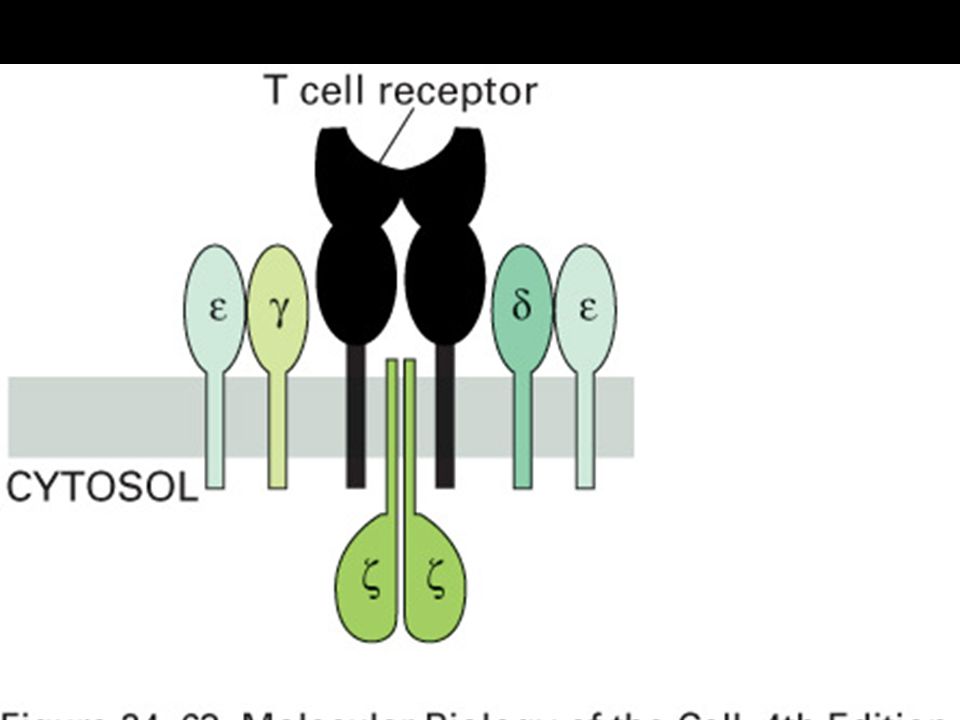
T-cell recognition of antigen is MHC-restricted. MHC polymorphisms reside in the domains that form the peptide-binding cleft. The TCR recognizes a complex of antigenic peptide and MHC. This co-recognition restricts the ability of the T cell to recognize antigen. MHC restriction may result from either direct MHC:TCR contact or an indirect effect on the peptides that bind or their bound conformation.

Podobné prezentace





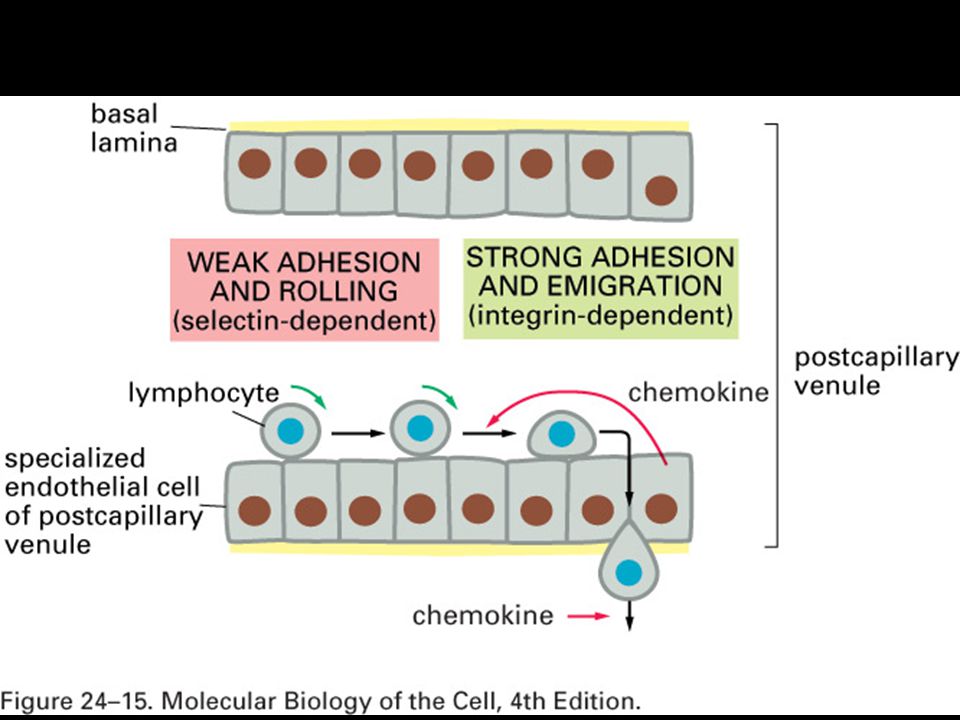
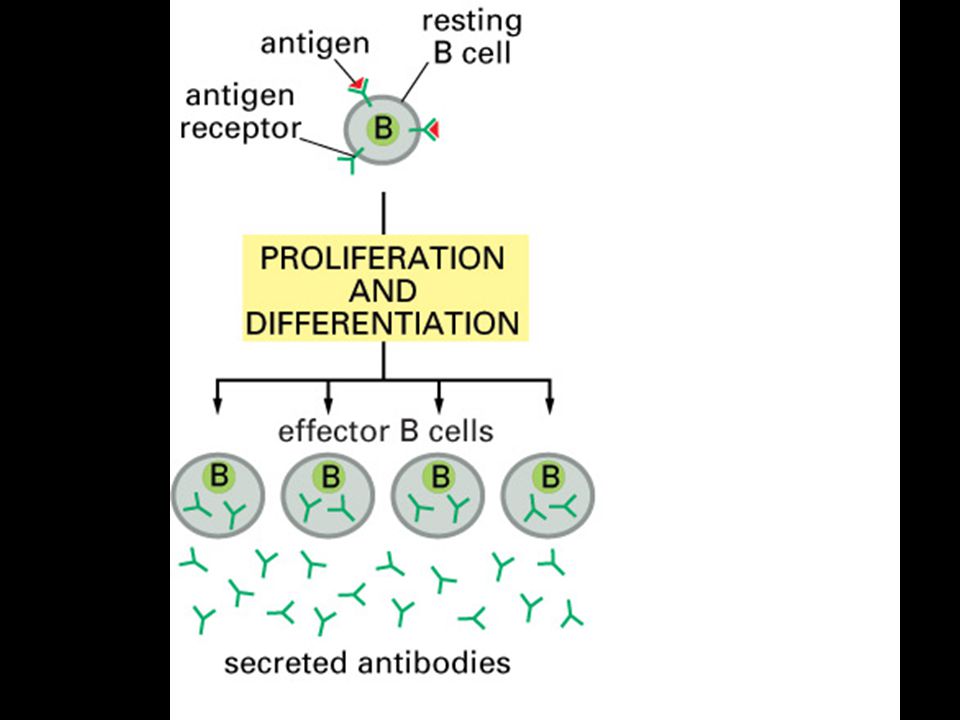
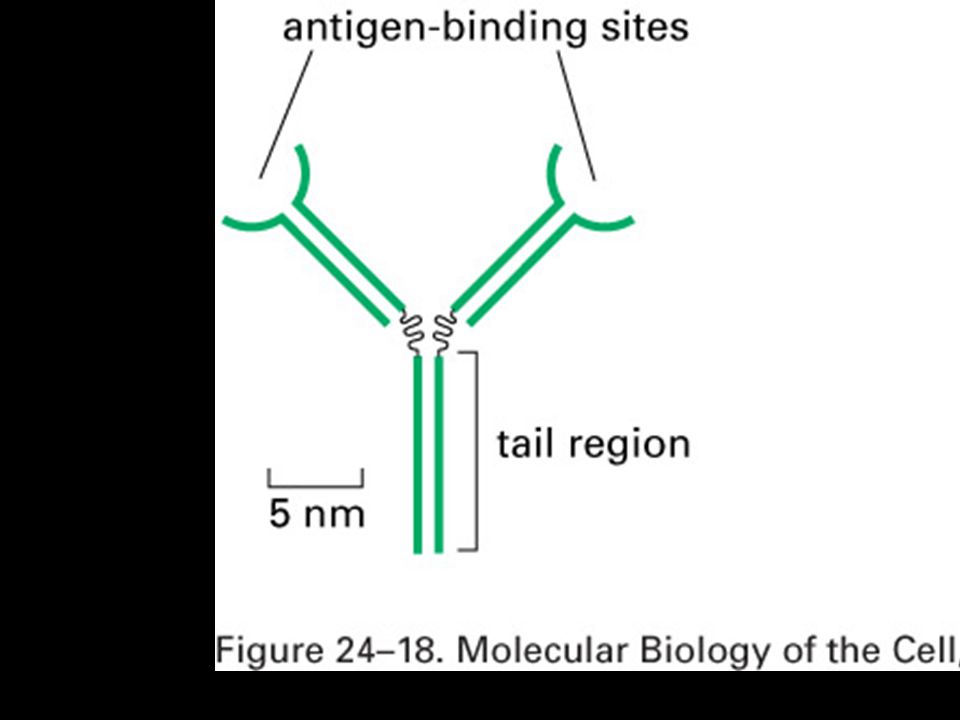
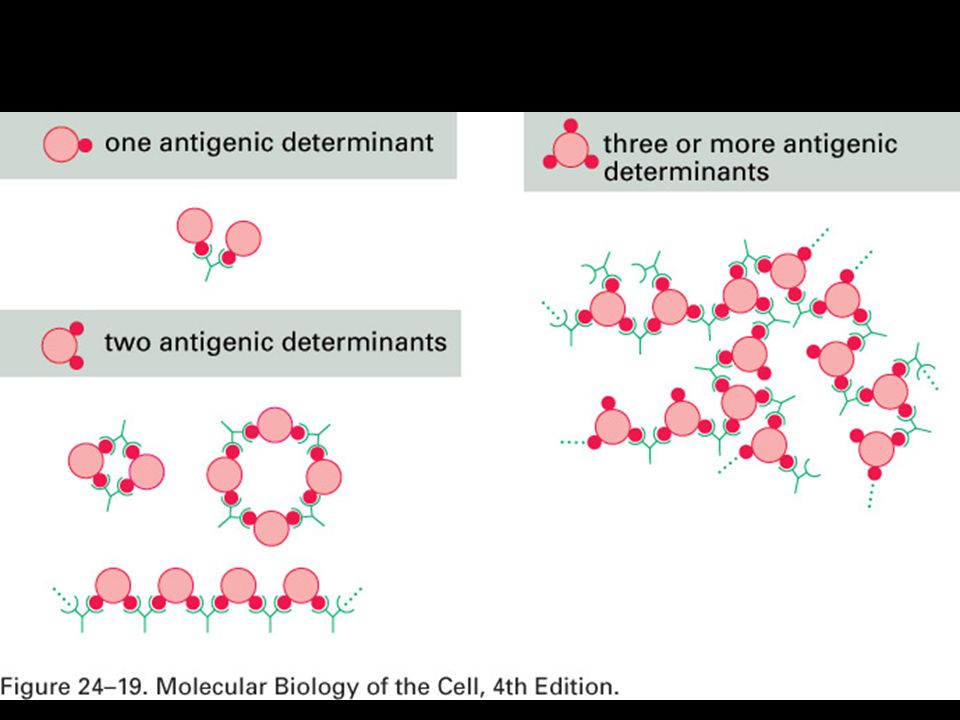
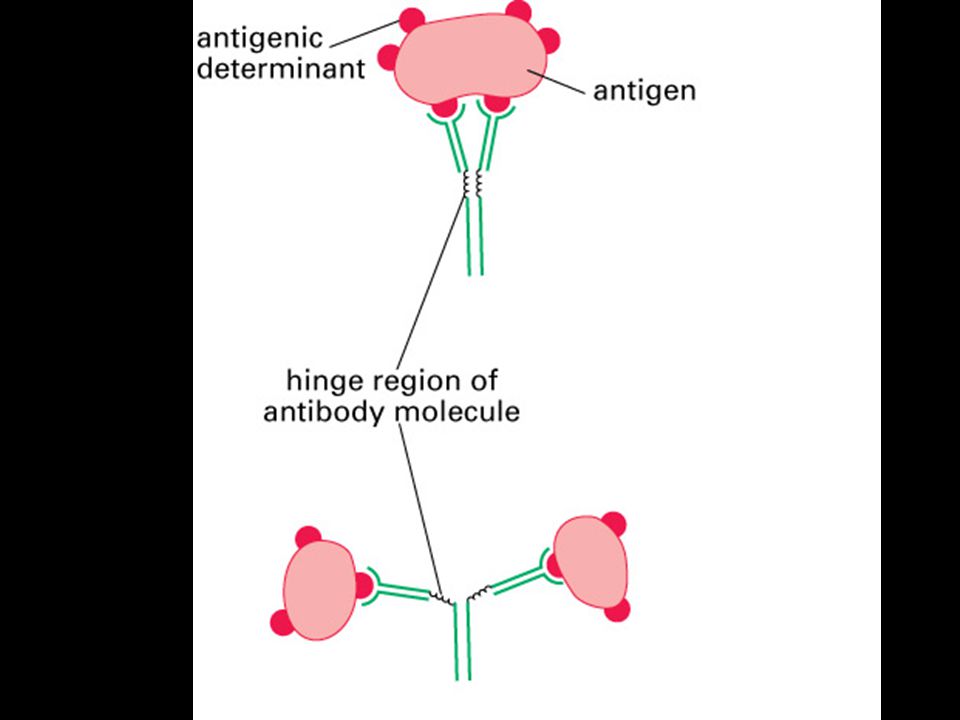
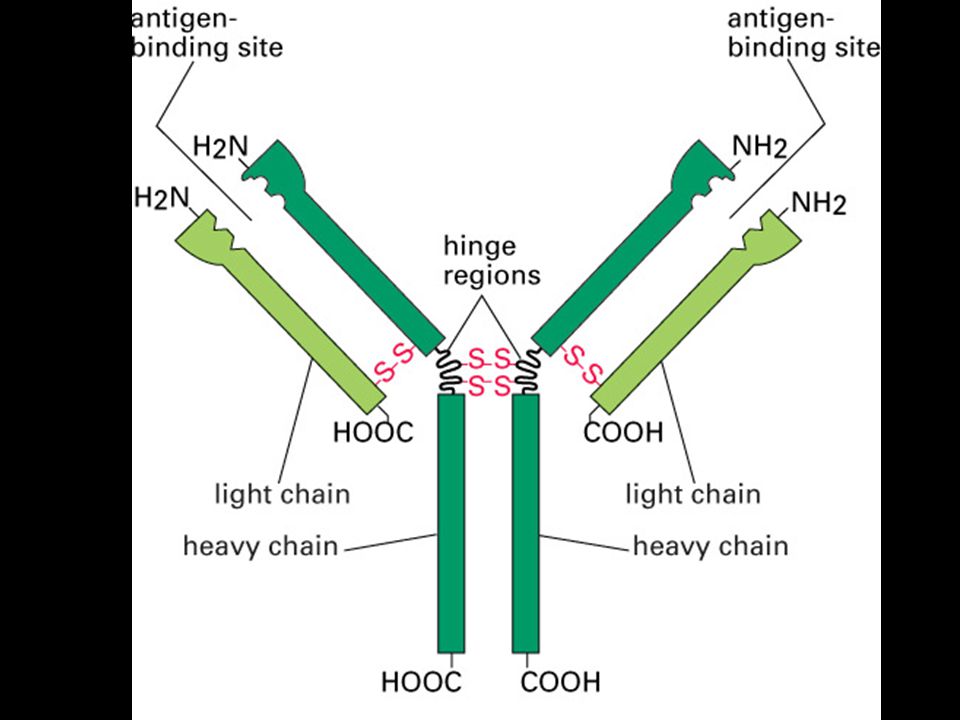
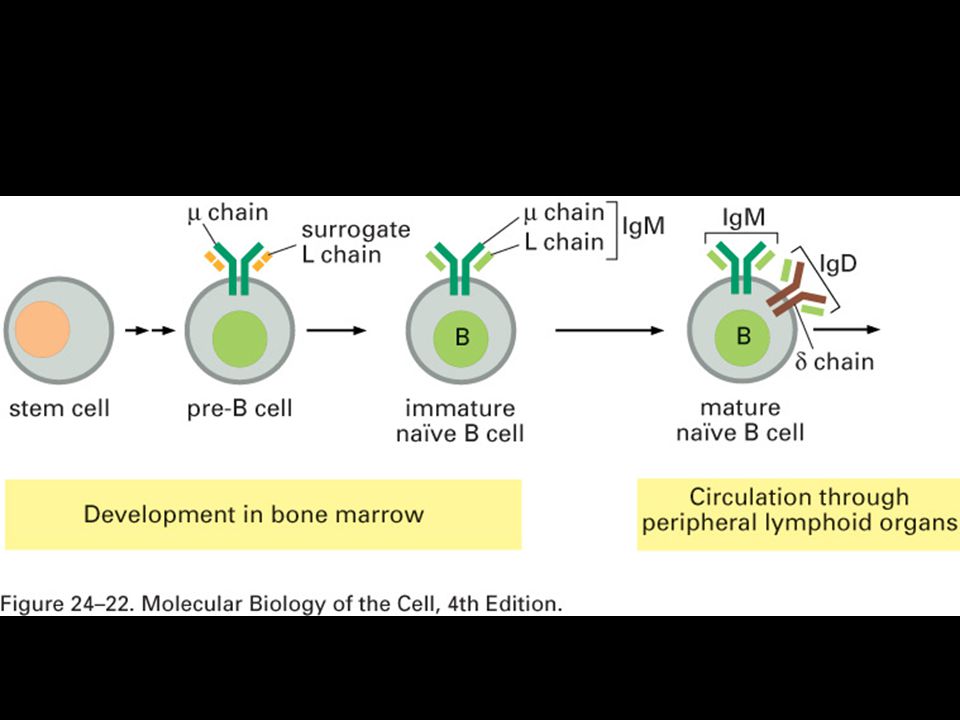
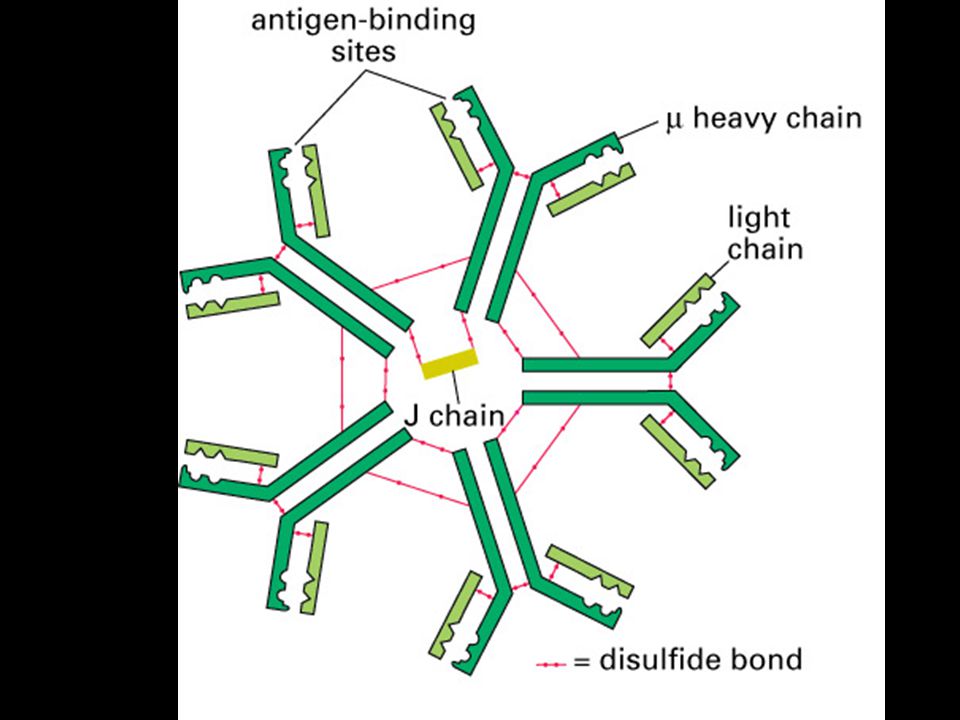













 Mgr. Martin Šmíd.>")
 Nespecifická : Fagocytóza granulocytů,monocytů.>")
















