Stáhnout prezentaci
1
Prosím, vypněte mobilní telefony

2
Ing. Zuzana Balounová, PhD.
Fyziologie rostlin Základní kurs 5 Fotosyntéza Ing. Zuzana Balounová, PhD.

3
Sekundární procesy asimilace CO2 (temnostní, syntetická fáze)
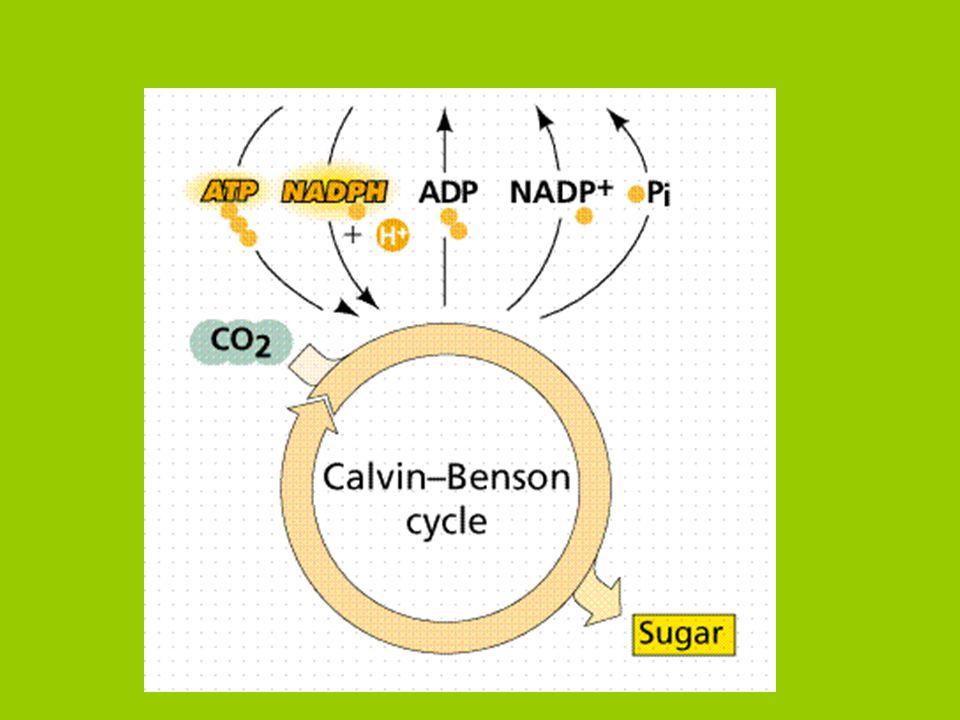
ATP a NADPH vzniklé v primárních pochodech mohou být použity k vázání a redukci CO2 v tzv. Calvinově cyklu (C3 cyklus). evolučně velmi starý proces společný všem fotosyntetizujícím organizmům. Žádný jiný cyklus se srovnatelnou funkcí není znám. redukce CO2 a syntéza org. sloučenin s vysokým energetickým obsahem ve stromatech chloroplastů
. evolučně velmi starý proces společný všem fotosyntetizujícím organizmům. Žádný jiný cyklus se srovnatelnou funkcí není znám. redukce CO2 a syntéza org. sloučenin s vysokým energetickým obsahem ve stromatech chloroplastů.")
4
Sekundární procesy asimilace CO2 (temnostní, syntetická fáze)
asimilace CO2 do organických sloučenin Calvinův cyklus 1) karboxylace 2) redukce 3) regenerace
 karboxylace. 2) redukce. 3) regenerace.")
5
1) karboxylace CO2 vázán na akceptor (ribulóza 1,5 biP)
bez dodání energie pomocí RUBISCA 2 molekuly 3-PGA (kyseliny 3-fosfoglycerové)
")
6
2) redukce Fosforylace (P z ATP) redukce (pomocí NADPH) na 3-PGAd
(3fosfoglyceraldehyd) Celkový náklad: 2 x ATP + 2 x NADPH
 Celkový náklad: 2 x ATP + 2 x NADPH.")
7
3) regenerace 3-PGA: Řetěz reakcí syntéza ribulózo-1,5-biP (RuBP)
(=akceptor pro další molekulu CO2 syntéza glukózy K čistému zisku 1 molekuly s 3C je třeba fixovat 3 x CO2 = cyklus proběhne 3x Zpracování 3C molekuly: - v chloroplastu (syntéza škrobu) - v cytosolu (jiné syntézy)
 - v cytosolu (jiné syntézy)")
8
Ze šesti molekul 3-fosfoglyceraldehydu je jen jedna použita k syntéze glukózy jako čistý výtěžek fixace tří molekul CO2 ostatních pět molekul se účastní regenerace pentózy RuBP.

9
Calvinův cyklus evolučně starý, jediný způsob asimilace C

11
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
12
„osud“ atomu uhlíku při asimilaci CO2
Převzato z: ( )
")
13
Calvinuv cyklus, C3 pentózofosfátová cesta fixace CO2
fixace CO2 přes ribulózo-1,5-bisfosfát (RuBP) (5C) který se ihned štěpí na 2 molekuly kys. 3-fosfoglycerové (PGA) (2x3C) fixace je katalyzována RuBP-karboxylázou
 (5C) který se ihned štěpí na 2 molekuly kys. 3-fosfoglycerové. (PGA) (2x3C) fixace je katalyzována RuBP-karboxylázou.")
14
ribulózo-1,5bisfosfát-karboxyláza/oxygenáza
Rubisco ribulózo-1,5bisfosfát-karboxyláza/oxygenáza Nejdůležitější enzym Calvinova cyklu (umožňující navázání C z CO2)
")
15
Rubisco –může fungovat nejen jako karboxyláza, ale i jako oxygenáza - na stejný substrát může vnášet i O2. Afinita Rubisco k CO2 je podstatně větší než k O2, ale v chloroplastech bývá v důsledku fotolýzy vody koncentrace O2 daleko vyšší, než CO2 oxygenázová aktivita Rubisco se prosadí a spustí se fotorespirace.

16
Fotorespirace Proces opačný k fotosyntéze, ztráta CO2 do okolí
probíhá jen na světle (oproti mitochondriální respiraci!) jev dán karboxylačně-oxidační aktivitou Rubisca (za běžných podmínek asi 4:1) vyšší parciální koncentrace CO2 vyšší fotosyntéza vyšší radiace a teplota vyšší fotorespirace běžně rostliny ztrácí okolo 20% získaného CO2
 jev dán karboxylačně-oxidační aktivitou Rubisca (za běžných podmínek asi 4:1) vyšší parciální koncentrace CO2 vyšší fotosyntéza. vyšší radiace a teplota. vyšší fotorespirace. běžně rostliny ztrácí okolo 20% získaného CO2.")
17
fotorespirace

18
Z P- glykolátu se odštěpí P
glykolát je transportován do peroxizomů: syntéza glycinu V mitochondriích další syntézy-uvolňuje se CO2 a NH3 Ztráta C 10% + energetické ztráty (ATP, NADPH) Snížení účinnosti fixace CO2 Enzym Rubisko se zachoval z doby, kdy měla atmosféra více CO2 a méně O2 – tehdy jen fce karboxylázy (?)
 Snížení účinnosti fixace CO2. Enzym Rubisko se zachoval z doby, kdy měla atmosféra více CO2 a méně O2 – tehdy jen fce karboxylázy ( )")
19
Fotorespirace probíhá za součinnosti různých organel: chloroplastů, peroxizómů a mitochondrií
Čistý výtěžek fotosyntézy (např. výnosy plodin) při intenzivní fotorespiraci u C3 rostlin – silný pokles
 při intenzivní fotorespiraci u C3 rostlin – silný pokles.")
20
fotorespirace

21
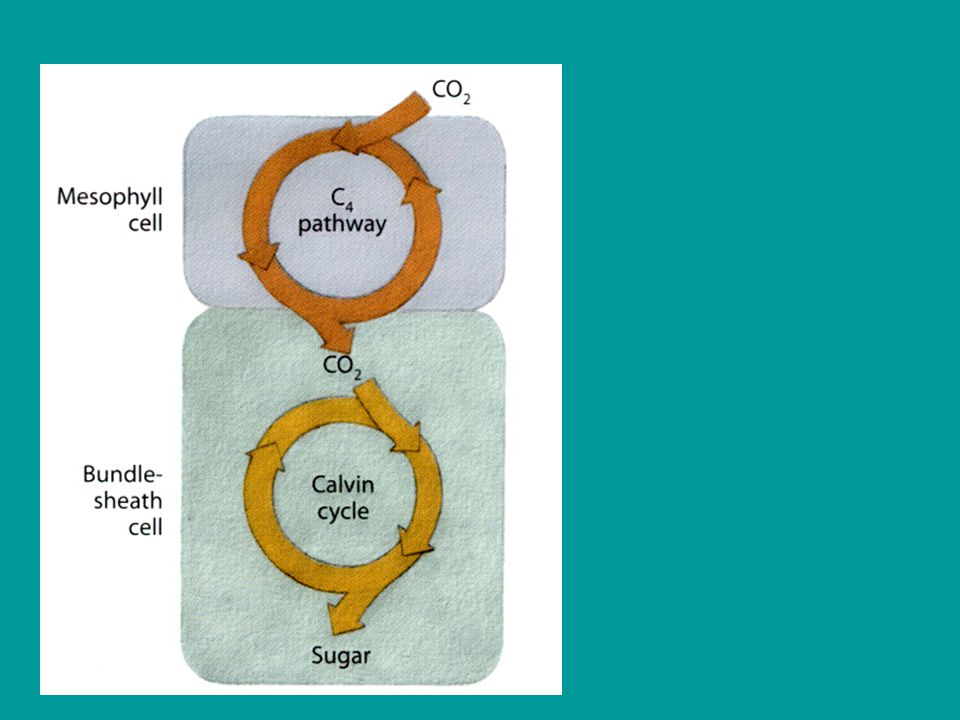
C4 - cyklus (Hatch-Slackova cesta)
V průběhu evoluce rostlin se vyvinuly některé pozoruhodné metaboIické adaptace, které umožňují zvýšit parciální tlak CO2 v místech jeho fixace Calvinovým cyklem a tím také potlačit oxygenázovou aktivitu Rubisco a tedy fotorespiraci Společným principem těchto adaptací je opakovaná karboxylace

23
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
24
není primárním akceptorem CO2 RuBP ale
U C4 rostlin není primárním akceptorem CO2 RuBP ale kyselina fosfoenolpyrohroznová (PEP) prvním stabilním produktem je C4 kyselina oxaloctová dále vznikají kyselina asparagová (asparát) kyselina jablečná (malát) ( C4 kyseliny s minimálně dvěma karboxyly)
 prvním stabilním produktem je. C4 kyselina oxaloctová. dále vznikají kyselina asparagová (asparát) kyselina jablečná (malát) ( C4 kyseliny s minimálně dvěma karboxyly)")
27
C4 - cesta dikarboxylových kyselin Hatch - Slackova
v buňkách pochev CS z OAA uvolňován CO2 následně vázán na RuBP (jako u C3-cesty) pyruvát se vrací do mezofylu
 pyruvát se vrací do mezofylu.")
28
Fixační cesta CAM (Crassulacean Acid Metabolism).
Sukulentní rostliny obsahují buňky s velkými vakuolami, které mají značnou skladovací kapacitu. Fixace CO2 u sukulentů vykazuje shodu s C4-cestou. Oba karboxylační procesy probíhají ale v téže buňce. Nejsou odděleny prostorově (jak je tomu u C4) ale časově (neprobíhají současně)
 ale časově (neprobíhají současně)")
29
Fotosyntéza – CAM cyklus

30
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
31
CAM - Crassulacean Acid Metabolism
Časové oddělení fixace CO2 a syntézy uhlíkatých sloučenin v noci fixace CO2 přes PEP, hromadění ve formě malátu ve vakuole ve dne malát převáděn zpět, dekarboxylován a v chloroplastech zpracován zpětně uvolněný CO2 Kelvinovým cyklem

32
Metabolické typy rostlin podle fixace uhlíku
C3-rostliny - převážná většina rostlin, primární způsob fixace C, asi 20% ztrát fotorespirací (díky oxidační aktivitě RuBP-karboxylázy) C4-rostliny - asi 2000 druhů rostlin (přibližně 20 čeledí), tropické trávy (Zea mays, Saccharum officinarum, čeledi Portulacaceae, Euphorbiaceae, Chenopodiaceae, také některé sinice (Anacystis nidulans), minimální ztráty fotorespirací, vyšší afinita PEP-karboxylázy vůči CO2 větší efektivita oproti C3 rostlinám CAM-rostliny - především sukulentní rostliny nejvíce z čeledí Agavaceae, Bromeliaceae, Cactaceae, Crassulaceae, Orchidaceae, ale také některé epifytické kapradiny a submerzní rostliny (Isoëtes, Crassula aquatica), smyslem CAM je minimalizovat ztráty vody
 C4-rostliny - asi 2000 druhů rostlin (přibližně 20 čeledí), tropické trávy (Zea mays, Saccharum officinarum, čeledi Portulacaceae, Euphorbiaceae, Chenopodiaceae, také některé sinice (Anacystis nidulans), minimální ztráty fotorespirací, vyšší afinita PEP-karboxylázy vůči CO2 větší efektivita oproti C3 rostlinám. CAM-rostliny - především sukulentní rostliny nejvíce z čeledí Agavaceae, Bromeliaceae, Cactaceae, Crassulaceae, Orchidaceae, ale také některé epifytické kapradiny a submerzní rostliny (Isoëtes, Crassula aquatica), smyslem CAM je minimalizovat ztráty vody.")
33
C3- vs. C4-rostliny Anatomická stavba listu - Krantz-anatomie u C4 (Krantz = věnec), tlustostěnné buňky obsahující chloroplasty tvořící věnčitý obal cévních svazků, není rozlišen palisádový a houbovitý mezofyl rozdílná afinita Rubisco a PEP-karboxylázy vůči CO2 minimální ztráty fotorespirací u C4 kompenzační bod a bod nasycení fotosyntézy vyšší u C4 geografické rozšíření - C4 schází v chladných klimatických oblastech
, tlustostěnné buňky obsahující chloroplasty tvořící věnčitý obal cévních svazků, není rozlišen palisádový a houbovitý mezofyl. rozdílná afinita Rubisco a PEP-karboxylázy vůči CO2. minimální ztráty fotorespirací u C4. kompenzační bod a bod nasycení fotosyntézy vyšší u C4. geografické rozšíření - C4 schází v chladných klimatických oblastech.")
34
C3 (tříuhlíkatý produkt fosfoglycerát):
u nás 99,7 % rostlin, součástí fotosyntézy je fotorespirace -rostliny ztrácejí % CO2, největší ztráty hmoty nastávají za horkých suchých dní, kdy je omezen výpar vody a v důsledku uzavření průduchů klesá koncentrace CO2 a podíl fotorespirace vzhledem k fotosyntéze vzrůstá C4 (čtřuhlíkatý produkt (oxalacetát): tropické, u nás 0,3 % rostlin (kukuřice, bambus, proso, cukrová třtina), rychle rostoucí, velice účinně vážou CO2, fotorespirace je potlačena, za horkých dnů produkují 2-3krát více glukózy než C3 rostliny za mírných dní jsou však C3 rostliny výkonnější, protože na fixaci CO2 spotřebují méně energie.
: tropické, u nás 0,3 % rostlin (kukuřice, bambus, proso, cukrová třtina), rychle rostoucí, velice účinně vážou CO2, fotorespirace je potlačena, za horkých dnů produkují. 2-3krát více glukózy než C3 rostliny. za mírných dní jsou však C3 rostliny výkonnější, protože na fixaci CO2 spotřebují méně energie.")
35
S Saccharum officinarum

36
Euphorbia sp.

37
Chenopodium sp.

38
Mesembryanthemaceae

40
Cactaceae

42
Anacystis nidulans (sinice)
")
43
Co se děje dál s asimiláty?
v chloroplastu - během fotosyntézy vznik sacharidů, bílkovin, lipidů, škrobu (tranzitorní š.) v cytoplazmě – škrob se rozloží na sacharózu, přenáší se cévními svazky a ukládá do zásoby (cukrová třtina, řepa) nebo vzniká zásobní škrob a ukládá se v zásobních orgánech (hlízy, plody, kořeny ...)
 v cytoplazmě – škrob se rozloží na sacharózu, přenáší se cévními svazky a ukládá do zásoby (cukrová třtina, řepa) nebo vzniká zásobní škrob a ukládá se v zásobních orgánech (hlízy, plody, kořeny ...)")
44
Fotosyntéza vodních rostlin
Vlastnosti vodního prostředí: Nízký příkon radiace (eufotická zóna) koncentrace plynů roste s tlakem nad hladinou a klesající teplotou, naopak klesá s obsahem rozpuštěných solí CO2 ve vodě snadno rozpustný, ale difúze asi 10,000x pomalejší oproti vzduchu nedostatečné zásobování CO2 k listům s rostoucím pH vody se posouvá rovnováha (H2CO3) H2O + CO2 H+ + HCO3- 2H+ + CO32- doprava, při pH > 9 schází volný CO2
 koncentrace plynů roste s tlakem nad hladinou a klesající teplotou, naopak klesá s obsahem rozpuštěných solí. CO2 ve vodě snadno rozpustný, ale difúze asi 10,000x pomalejší oproti vzduchu nedostatečné zásobování CO2 k listům. s rostoucím pH vody se posouvá rovnováha (H2CO3) H2O + CO2 H+ + HCO3- 2H+ + CO32- doprava, při pH > 9 schází volný CO2.")
45
Adaptace vodních rostlin na příjem uhlíku
Příjem celým povrchem těla (nejsou přítomny průduchy) - maximálně dělené listy (Myriophyllum, Ceratophyllum, Batrachium) redukce hraniční vrstvy okolo listů - snižuje ji proudění vody, zmenšení plochy listů (hraniční vrstva roste s kvadrátem listové plochy) příjem HCO3- : 1. aktivní transport (sinice, řada zelených řas), 2. převod na CO2 v buněčné stěně pomocí enzymu karbonát-anhydrázy (Elodea canadensis), 3. protonový polární transport - vylučování H+ lokální zvýšení acidity posun rovnováhy směrem k volnému CO2 (Characeae, makrofyta) využití CO2 z organického sedimentu - příjem kořeny a rozvod skrze kanálky k listům (Isoëtes, Litorella uniflora, Lobelia dortmana)
 - maximálně dělené listy (Myriophyllum, Ceratophyllum, Batrachium) redukce hraniční vrstvy okolo listů - snižuje ji proudění vody, zmenšení plochy listů (hraniční vrstva roste s kvadrátem listové plochy) příjem HCO3- : 1. aktivní transport (sinice, řada zelených řas), 2. převod na CO2 v buněčné stěně pomocí enzymu karbonát-anhydrázy (Elodea canadensis), 3. protonový polární transport - vylučování H+ lokální zvýšení acidity posun rovnováhy směrem k volnému CO2 (Characeae, makrofyta) využití CO2 z organického sedimentu - příjem kořeny a rozvod skrze kanálky k listům (Isoëtes, Litorella uniflora, Lobelia dortmana)")
46
CAM vodních makrofyt CAM submerzních makrofyt - cirkadiánní změny koncentrace malátu a pH v buňkách, popsáno u Isoëtes howellii, mají C3, CAM slouží jako dodatečný zdroj

47
Isoëtes andicola Terestrická rostlina s listy bez průduchů, velmi silná kutikula - nulová výměna plynů s atmosférou příjem CO2 kořeny z organického sedimentu CAM - probíhá v noci, malý příjem CO2 ze sedimentu a recyklace respiratorního CO2, dodatečný metabolismus k dennímu C3








>")
















