Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Fotoinhibice, fotopoškození a fotoprotekční mechanismy

2
Měření množství dopadající energie světla
Ozářenost – W.m-2 (osvětlenost – ln .m-2 = lux) Fotonová (kvantová) ozářenost – mol.s-1.m-2 Vzájemné převody závisí na vlnových délkách fotonů (tedy přesné jsou jen pro monochromatické světlo)!
 Fotonová (kvantová) ozářenost – mol.s-1.m-2. Vzájemné převody závisí na vlnových délkách fotonů. (tedy přesné jsou jen pro monochromatické světlo)!")
3
Závislost rychlosti čisté fotosyntézy (PN) na ozářenosti (PAR)
- s rostoucí ozářeností klesá účinnost využití záření e - kvantový výtěžek e =mol CO2/mol kvant (kvantum = foton) - max. teoretická hodnota = 0,125 mol CO2 / mol kvant (min. kvantový požadavek Q (e = 1/Q) je: Q = 8 mol kvant / mol CO2) 2 H2O 4 e- 4 fotony v PSII 4 fotony PSI 2 NADPH Na redukci každé ze dvou fosfoglycerových kys. vzniklých po navázání CO2 na ribulóza-1,5- bisfosfát je potřeba 1 NADPH (které se oxiduje)
 - max. teoretická hodnota = 0,125 mol CO2 / mol kvant (min. kvantový požadavek Q (e = 1/Q) je: Q = 8 mol kvant / mol CO2) 2 H2O 4 e- 4 fotony v PSII 4 fotony PSI 2 NADPH. Na redukci každé ze dvou fosfoglycerových kys. vzniklých po navázání CO2 na ribulóza-1,5- bisfosfát je potřeba 1 NADPH (které se oxiduje)")
4
Fotoinhibice Fotopoškození (snížení počtu fčních fotosystémů)
soubor procesů projevujících se snížením rychlosti fotosyntézy při zvyšující se ozářenosti Fotopoškození (snížení počtu fčních fotosystémů) - fotosyntetických struktur (proteinů) - UV světlo, oxidativní poškození (ROS, P680+, …) - nastává i za nízké ozářenosti, ale rychlá oprava – nenastává fotoinhibice Fotoprotekce (snížení E přenesené do RC … k fixaci CO2) - předcházení fotopoškození (rychle vratné změny) ROS: 1O2
 - fotosyntetických struktur (proteinů) - UV světlo, oxidativní poškození (ROS, P680+, …) - nastává i za nízké ozářenosti, ale rychlá oprava – nenastává fotoinhibice. Fotoprotekce (snížení E přenesené do RC … k fixaci CO2) - předcházení fotopoškození (rychle vratné změny) ROS: 1O2.")
5
Fotopoškození – mechanismy a cíle
PSII blok transportu elektronů poškození D1 (D2, proteiny cyt b559) Vysoká ozářenost: akceptorová strana PSII – málo PQ – 3P680* – 1O2 donorová strana 1 - OEC poškození (UV), odpoutání vnějších proteinů 2 – P oxidativní poškození okolí Nízká ozářenost: QB- - rekombinace náboje zpět na P680 – 3P680* – 1O2
 Vysoká ozářenost: akceptorová strana PSII – málo PQ – 3P680* – 1O2. donorová strana 1 - OEC poškození (UV), odpoutání vnějších proteinů. 2 – P oxidativní poškození okolí. Nízká ozářenost: QB- - rekombinace náboje zpět na P680 – 3P680* – 1O2.")
6
Fotopoškození – mechanismy a cíle
PSI - jen při nízké teplotě (zřejmě inhibicí aktivity SOD Cu/Zn) - kumulativní poškození celého PSI 1. O2- poškození FeS klastrů (FA, FB) – blok transportu e- Pomalá oprava – mnoho dní po přenesení do tepla
 - kumulativní poškození celého PSI. 1. O2- poškození FeS klastrů (FA, FB) – blok transportu e- Pomalá oprava – mnoho dní po přenesení do tepla.")
7
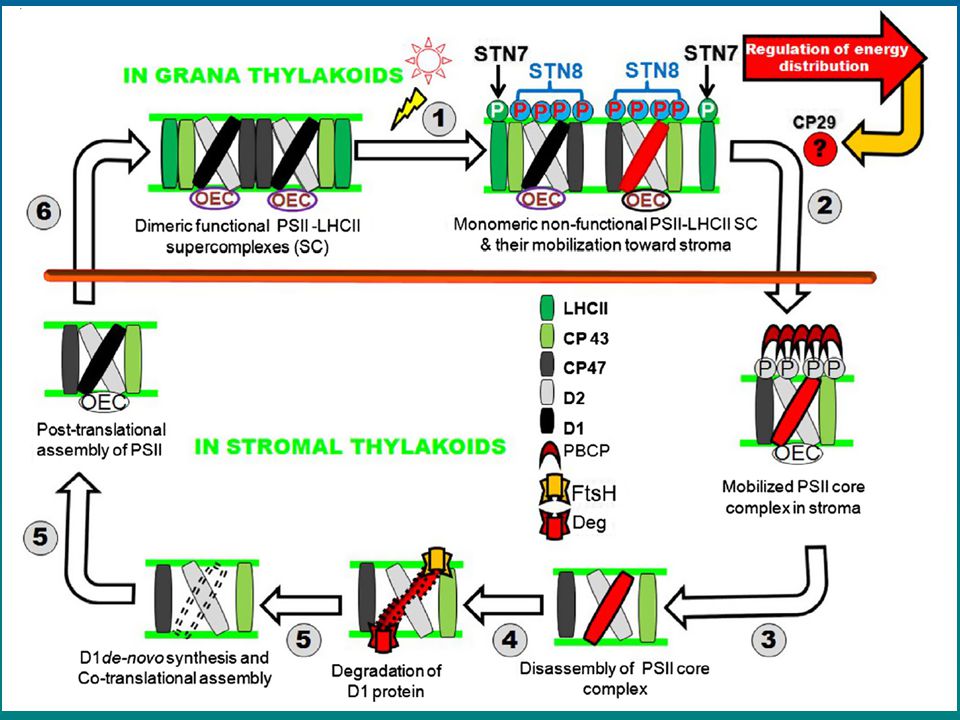
Oprava PSII – výměna poškozeného D1
Odbourání fosforylace proteinů PSII (STN8, příp. STN7) odpoutání LHCII, monomerizace transport do strom. membrán, odpoutání OEC proteinů defosforylace PBCP (TLP40) degradace D1 (FtsH, degH) Vložení nového kotranslačně (interakce s D2 a cyt b559) odštěpení C-konce v luminu interakce s anténami CP43, CP47 připojení dalších podjednotek (OEC) monomer do gran, dimerizace, LHCII Oprava PSII – výměna poškozeného D1 (PSII repair cycle)
 odpoutání LHCII, monomerizace. transport do strom. membrán, odpoutání OEC proteinů. defosforylace PBCP (TLP40) degradace D1 (FtsH, degH) Vložení nového. kotranslačně (interakce s D2 a cyt b559) odštěpení C-konce v luminu. interakce s anténami CP43, CP47. připojení dalších podjednotek (OEC) monomer do gran, dimerizace, LHCII. Oprava PSII – výměna poškozeného D1. (PSII repair cycle)")
9
Fotoinhibice je způsobena příliš silnou ozářeností
ALE (při bloku repair – chloramfenicol, lincomycin) - rychlost poškození je zcela úměrná ozářenosti! (tedy nastává i při nízké!) - účinné vlnové délky se liší od absorpce Chl a karotenoidů - nemění se inhibicí DCMU (blok QA → QB) ani glykol (glycer)aldehydem (blok fosforibulokinase), ani ROS! Původní představa: - D1 je poškozován přebytkem záření zachyceného fotosyntetickými pigmenty
 - rychlost poškození. je zcela úměrná ozářenosti! (tedy nastává i při nízké!) - účinné vlnové délky se liší. od absorpce Chl a karotenoidů. - nemění se inhibicí DCMU. (blok QA → QB) ani glykol. (glycer)aldehydem (blok. fosforibulokinase), ani ROS! Původní představa: - D1 je poškozován přebytkem záření zachyceného. fotosyntetickými pigmenty.")
10
Revize: ozářenost vede k fotopoškození D1,
příliš silná ozářenost vede k inhibici reparace D1 Primární poškození – ne fotos. absorpce, ne ROS Cíl - Mn klastr - přímá absorpce záření (UV, žluté) - uvolnění Mn – neredukuje se primární donor (alt. donor ascorbate) - P680+ poškození D1 Výměna poškozeného D1 (PSII repair cycle) - silně inhibována ROS (~ ozářenosti!) - inhibice translace D1 (EF-G)
 - uvolnění Mn – neredukuje se primární donor. (alt. donor ascorbate) - P680+ poškození D1. Výměna poškozeného D1 (PSII repair cycle) - silně inhibována ROS (~ ozářenosti!) - inhibice translace D1 (EF-G)")
11
Fotoinhibice (fotopoškození)
– důsledek nerovnováhy mezi poškozením a opravnými pochody

12
Fotoprotekce - ochrana před nadbytečným
tokem energie Strukturní úroveň listu - paraheliotropismus (x dia-) - tenčí listy, ochranné pigmenty (x UV, VIS) (fenolické l., antokyany, flavonoidy) úroveň buňky: dlouhodobé: méně chloroplastů, krátkodobé: pohyb chloroplastů - PHOT1, 2, CHUP1, aktin, … - méně poškození, méně ROS (repair) pevné nastavení – není tropismus!
 - tenčí listy, ochranné pigmenty (x UV, VIS) (fenolické l., antokyany, flavonoidy) úroveň buňky: dlouhodobé: méně chloroplastů, krátkodobé: pohyb chloroplastů. - PHOT1, 2, CHUP1, aktin, … - méně poškození, méně ROS (repair) pevné nastavení. – není tropismus!")
13
Fotoprotekce Zhášení ROS: - tvorba na PSII i PSI
- blok chloroplastové translace

14
Fotoprotekce Změny na úrovni chloroplastu
pomalejší: méně LHCII a LHCI ku PSII a PSI (rel. rychlá degradace LHC 1,2,3 a 6) méně PSI a PSII ku Rubisco krátkodobé: zvýšení disipace - Xantofylový cyklus (+ protonace PsbS) PQH/PQ → STN7 → fosforylace LHCII → posílení cyklického transportu elektronů → vyšší gradient H+ → aktivace VDE → vyšší disipace → méně ROS → rychlejší reparace D1 fosforylace LHCII (STN7 kinase – Lhcb 1,2,4) – cyklický transport PSI
 méně PSI a PSII ku Rubisco. krátkodobé: zvýšení disipace - Xantofylový cyklus (+ protonace PsbS) PQH/PQ → STN7 → fosforylace LHCII → posílení. cyklického transportu elektronů → vyšší gradient H+ → aktivace VDE → vyšší disipace → méně ROS → rychlejší. reparace D1. fosforylace LHCII (STN7 kinase – Lhcb 1,2,4) – cyklický transport PSI.")
15
Xanthofyly anteraxanthin a zeaxanthin
- zvýšená disipace světelné energie (teplo): chlexcit + karotenoid => karotenoidexcit + chl karotenoid + teplo (kinetická E) RC přenos energie je doprovázen přenosem elektronu na chl a
: chlexcit + karotenoid => karotenoidexcit + chl. karotenoid + teplo (kinetická E) RC. přenos energie je doprovázen. přenosem elektronu na chl a.")
16
Xantofylový „cyklus“ - modifikace karotenoidů
- (de)epoxidace při poklesu pH - zvýšená schopnost odvádění nadbytečné energie z chlorofylu - méně energie do fotochemie Diurnální změny - violaxanthin a zeaxanthin anteraxanthin – poměrně stabilní Místo disipace - PsbS a LHCII (?)
epoxidace při poklesu pH. - zvýšená schopnost odvádění nadbytečné energie z chlorofylu. - méně energie do fotochemie. Diurnální změny. - violaxanthin a zeaxanthin. anteraxanthin – poměrně stabilní. Místo disipace - PsbS a LHCII ( )")
17
Disipace světelné energie xantofyly
Regulace prostřednictvím pH v lumen tylakoidu - prostřednictvím violaxanthin deepoxidase (VDE) - PsbS protein (protonizace – vazba zeaxanthinu) - až 80 % exc. singletových chlorofylů - u zastíněných zůstává deepoxid. stav déle - u jehličnanů v zimě permanentně deepoxidováno Disipace světelné energie = nefotochemické zhášení fluorescence chlorofylu
 - PsbS protein (protonizace – vazba zeaxanthinu) - až 80 % exc. singletových chlorofylů. - u zastíněných zůstává deepoxid. stav déle. - u jehličnanů v zimě permanentně deepoxidováno. Disipace světelné energie. = nefotochemické zhášení fluorescence chlorofylu.")
18
Osudy energie fotonu po jeho absorpci molekulou chlorofylu
Uvolnění elektronu v RC - fotochemie Přenos energie na další molekulu pigmentu Reakce s O2 Disipace (teplo)
")
19
Fluorescence chlorofylu:
680 nm LHCII 685 nm CP43 695 nm CP47 720 nm PSI core 740 nm LHCI

20
Fluorescence chlorofylu:
za optimálních podmínek: 0,6 – 3 % re-emitováno v podobě fluorescence (pokud je elektron delší dobu v excitovaném stavu!) (chlorofyl a v roztoku 30 %) Zhášení fluorescence - fotochemické 80 % absorbované světelné energie použito pro fotochemii (= separace elektronu – využití energie na syntézu ATP a redukci NADP+) = zhášení fluorescence - nefotochemické cca 15 % vyzářeno v podobě tepla – zahřívání listu Využívá se k měření účinnosti fotosyntézy (vyšší výtěžek fluorescence = nižší účinnost fotochemie)
 (chlorofyl a v roztoku 30 %) Zhášení fluorescence - fotochemické. 80 % absorbované světelné energie použito pro fotochemii. (= separace elektronu – využití energie na syntézu ATP a redukci NADP+) = zhášení fluorescence. - nefotochemické. cca 15 % vyzářeno v podobě tepla – zahřívání listu. Využívá se k měření účinnosti fotosyntézy. (vyšší výtěžek fluorescence = nižší účinnost fotochemie)")
21
Fluorescence chlorofylu (bez dalších
pulzů = Kautského křivka) Fotochemické zhášení qP = (Fm’ – Fs)/(Fm’ – F0’ ) kolik E jde k PSI Nefotochemické zhášení qN = (Fm – Fm’)/(Fm – F0’ ) S sat. pulzy FR light F0 – všechny PSII mohou přijímat energii (= min. fluorescence) Fm –žádný PSII nemůže přijmout energii (saturační pulz) (QA red. = max. fluorescence) Fv = Fm – F0 … variabilní fluorescence Fv / Fm … účinnost využití excitační energie otevřenými PSII (max. účinnost) (poměrně konstantní = 0,83 kvantovému požadavku 9-10 fotonů na O2 ) Fs – „steady state“ fluorescence při daném osvětlení
 Fotochemické zhášení. qP = (Fm’ – Fs)/(Fm’ – F0’ ) kolik E jde k PSI. Nefotochemické zhášení. qN = (Fm – Fm’)/(Fm – F0’ ) S. sat. pulzy FR light. F0 – všechny PSII mohou přijímat energii (= min. fluorescence) Fm –žádný PSII nemůže přijmout energii (saturační pulz) (QA red. = max. fluorescence) Fv = Fm – F0 … variabilní fluorescence. Fv / Fm … účinnost využití excitační energie otevřenými PSII (max. účinnost) (poměrně konstantní = 0,83 kvantovému požadavku 9-10 fotonů na O2 ) Fs – „steady state fluorescence při daném osvětlení.")
22
Nefotochemické zhášení fluorescence chlorofylu = fotoprotekce
qE – feed-back deexcitation (pH aktivace xantofylů disipace) qT – state transition (fosforylace LHCII) qI – fotoinhibice (poškozené fotosystémy disipují)
 qT – state transition (fosforylace LHCII) qI – fotoinhibice (poškozené fotosystémy disipují)")
23
Využití energie záření - účinnost využití difúzního záření je vyšší

24
Asimilace záření listem
Anatomické adaptace: Epidermis – (koncentrování světla) Palisádový – prostup světla Houbový – odrazy na površích (rozptyl)
 Palisádový – prostup světla. Houbový – odrazy na površích (rozptyl)")
25
Variabilita asimilačních orgánů
Vybrané fyziologické charakteristiky listů: (2548 druhů na 175 lokalitách planety) Specifická hmotnost 14 až 1500 g (DM) m-2 Kapacita fotosyntézy (PN) 5 až 660 nmol (CO2) g-1 s-1 Obsah N v sušině listů 0,2 až 6,4 % Doba života: 0,9 až 288 měsíců Velikost listové čepele - různé strategie investic do vytváření listů v závislosti na vnějších podmínkách - Wright et al. 2004, Nature
 Specifická hmotnost 14 až 1500 g (DM) m-2. Kapacita fotosyntézy (PN) 5 až 660 nmol (CO2) g-1 s-1. Obsah N v sušině listů 0,2 až 6,4 % Doba života: 0,9 až 288 měsíců. Velikost listové čepele. - různé strategie investic do vytváření listů v závislosti na vnějších podmínkách. - Wright et al. 2004, Nature.")
26
Celková účinnost využití záření
se v porostu zvyšuje 20,7x1= =20,7 17,6x2= =35,2 13x4= =52 8x8= =64

27
Využití záření – struktura porostu
Absorpce záření porostem velmi záleží na struktuře porostu a LAI – leaf area index – kumulativní index pokryvnosti listoví (plocha listů / plocha půdy) LAI – optimálně hodnoty 4 – 5 Adaptace k asimilaci v porostu: (konkurovat ostatním x nekonkurovat sobě) anatomie listu biochemie listu (rubisco, chlorofyl, xathofyly) stomy, liány, epifyty, růžice fylotaxe natáčení listů (úhel)
 LAI – optimálně hodnoty 4 – 5. Adaptace k asimilaci v porostu: (konkurovat ostatním x nekonkurovat sobě) anatomie listu. biochemie listu (rubisco, chlorofyl, xathofyly) stomy, liány, epifyty, růžice. fylotaxe. natáčení listů (úhel)")
28
Produkce biomasy rostlin (hospodářského výnosu plodin)
Hospodářský výnos (g m-2 rok-1) YP = PN . HI PN = S . εi . εc / k kde YP: hospodářský výnos (g m-2 rok-1) HI: sklizňový index (podíl hospodářsky hlavního produktu a hmotnosti sušiny celé rostliny, porostu) PN: čistá fotosyntetická produkce S: roční suma slunečního záření (MJ m-2 rok-1) εi: účinnost pohlcení záření dopadajícího na porost rostlinami εc: účinnost přeměny pohlceného záření ve fotosyntéze k: obsah energie v biomase (MJ g-1)
 YP = PN . HI PN = S . εi . εc / k. kde. YP: hospodářský výnos (g m-2 rok-1) HI: sklizňový index (podíl hospodářsky hlavního produktu a hmotnosti sušiny celé rostliny, porostu) PN: čistá fotosyntetická produkce. S: roční suma slunečního záření (MJ m-2 rok-1) εi: účinnost pohlcení záření dopadajícího na porost rostlinami εc: účinnost přeměny pohlceného záření ve fotosyntéze k: obsah energie v biomase (MJ g-1)")
29
Fotosyntetická produktivita ekosystémů
na Zemi Celosvětová produkce: 105 Pg (C) rok-1 tedy 200 g (C) rok-1 m-2 (P – peta – 1015) 1 g C > 3,7 g CO2 a 2,5 g DM (dry matter) 1 g CO2 > 0,27 g C a 0,675 g DM 1 g DM > 1,47 g CO2 a 0,378 g C
 rok-1 tedy 200 g (C) rok-1 m-2. (P – peta – 1015) 1 g C > 3,7 g CO2 a 2,5 g DM (dry matter) 1 g CO2 > 0,27 g C a 0,675 g DM. 1 g DM > 1,47 g CO2 a 0,378 g C.")
30
Ilustrace pro podmínky ČR
Charakteristické hodnoty: S = J m-2 rok-1 εi = 0,8 εc = 0,01 k = J εi: účinnost pohlcení záření εc: účinnost přeměny záření k: obsah energie v biomase (MJ g-1) Produkce biomasy PN = ,8 . 0,01 / = = g m-2 rok-1 = = 2,285 kg m-2 rok-1 Hospodářský výnos YP = PN . HI = 22,85 . 0,5 = 11,43 t ha-1 rok-1
 Produkce biomasy. PN = ,8 . 0,01 / = = 2285 g m-2 rok-1 = = 2,285 kg m-2 rok-1. Hospodářský výnos YP = PN . HI = 22,85 . 0,5 = 11,43 t ha-1 rok-1.")
31
Možnosti zvyšování produkce biomasy a výnosů polních plodin
S: - je určeno geografickou polohou εi: - je dáno fyzikálními vlastnostmi porostu lze ovlivnit strukturou porostu dynamikou rozvoje listů během vegetace εc: - lze teor. zvýšit optimalizací ozářenosti jednotlivých listů - vertikální a prostorová orientace anatomická stavba listů (?) fyziologických funkcí (obsah N, snížení RL,…) zvýšení aktivity rubisco HI: - maximum je patrně 0,6 (dosaženo u obilnin)
 - fyziologických funkcí (obsah N, snížení RL,…) - zvýšení aktivity rubisco. HI: - maximum je patrně 0,6 (dosaženo u obilnin)")
32
Využití slunečního záření ve fotosyntéze
Příčiny ztráty Ztráta (%) Využitelný zůstatek Ne FAR , ,0 Odraz a propustnost ,0 (10) ,0 Absorpce nefotos. částmi ,8 (4) ,2 Fotochem. neúčinnost (teplo) 8,4 (20) ,8 Typ fotosyntézy C C C C4 Metabolismus ,8 (65) , , ,0 (syntéza sacharidů) Fotorespirace ,5 (10) , Temnotní dýchání ,4 (10) , Výsledná účinnost využití ve fotosyntéze ,1 % ,0 % v období vegetace (z absorbovaného PAR (43 %) = cca %)
 Využitelný zůstatek Ne FAR 50,0 50,0. Odraz a propustnost 5,0 (10) 45,0. Absorpce nefotos. částmi 1,8 (4) 43,2. Fotochem. neúčinnost (teplo) 8,4 (20) 34,8. Typ fotosyntézy C3 C4 C3 C4. Metabolismus 22,8 (65) 24,8 12,0 10,0 (syntéza sacharidů) Fotorespirace 3,5 (10) 0 8,3 10 Temnotní dýchání 3,4 (10) 4 5,1 6. Výsledná účinnost využití ve fotosyntéze 5,1 % 6,0 % v období vegetace (z absorbovaného PAR (43 %) = cca %)")
Podobné prezentace