Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
NK buňky Interferony

2
NK buňky Součást antigenně nespecifických mechanismů
Nemají antigenně specifické receptory Rozeznávají bb., které mají abnormálně málo MHCgpI (některé nádorové a virem infikované bb.) Jsou schopny zabíjet rychle – bez předchozí stimulace, proliferace a diferenciace Aktivátory NK bb. – IFNa, IFNb
 Jsou schopny zabíjet rychle – bez předchozí stimulace, proliferace a diferenciace. Aktivátory NK bb. – IFNa, IFNb.")
3
Stimulační receptory NK bb
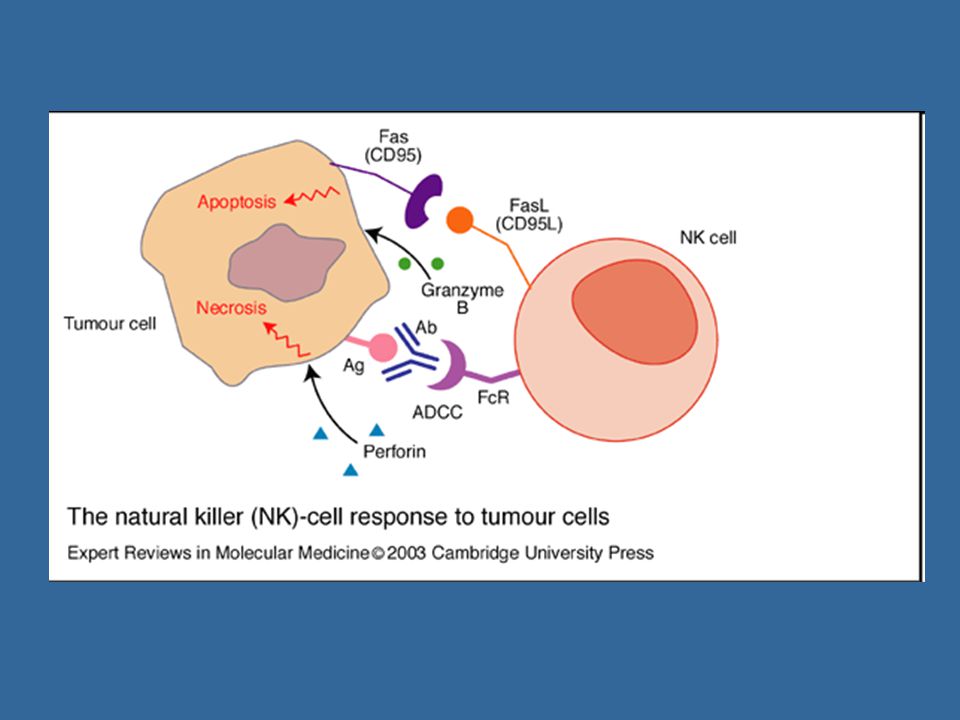
Stimulační receptory NK bb. - některé povrchové lektiny, Fc receptor CD16 ADCC (antibody-dependent cellular cytotoxicity) cytotoxická reakce závislá na protilátkách; NK b. prostřednictvím Fc receptorů CD16 rozpozná buňku opsonizovanou protilátkami třídy IgG, to vede k aktivaci cytotoxických mechanismů (degranulaci NK bb.) Inhibiční receptory NK bb. – signály poskytnuté prostřednictvím těchto receptorů inhibují cytotoxické mechanismy Imunoglobulinová skupina – tzv. KIR (killer inhibitor receptors) C-lektinová skupina – např. CD94/NKG2
 cytotoxická reakce závislá na protilátkách; NK b. prostřednictvím Fc receptorů CD16 rozpozná buňku opsonizovanou protilátkami třídy IgG, to vede k aktivaci cytotoxických mechanismů (degranulaci NK bb.) Inhibiční receptory NK bb. – signály poskytnuté prostřednictvím těchto receptorů inhibují cytotoxické mechanismy. Imunoglobulinová skupina – tzv. KIR (killer inhibitor receptors) C-lektinová skupina – např. CD94/NKG2.")
4
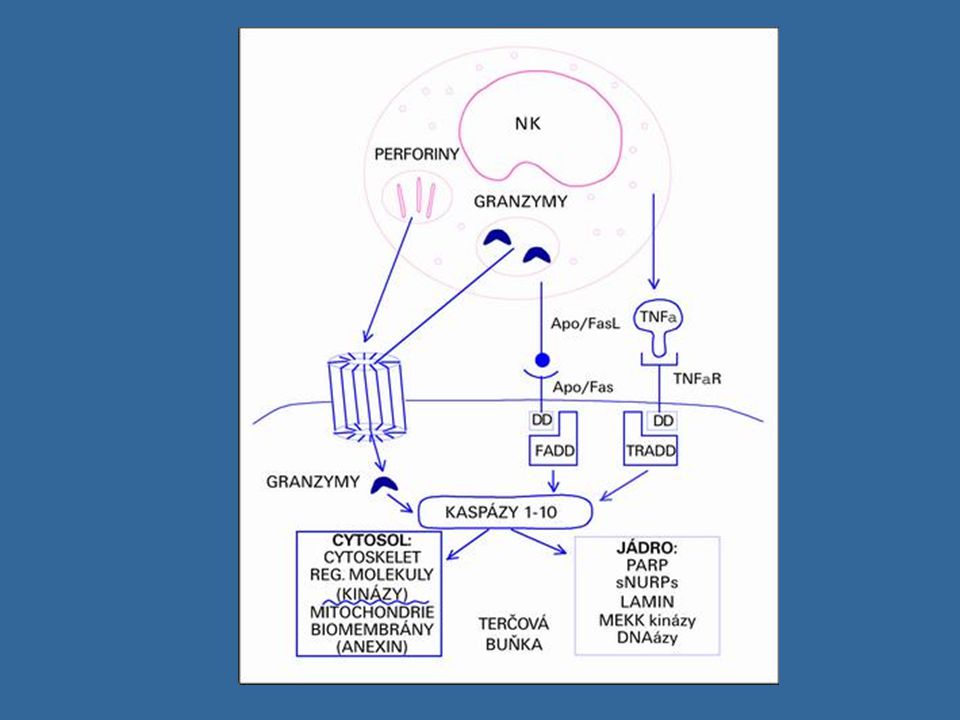
Cytotoxické mechanismy NK buněk
Výsledná reakce NK b. po setkání s jinou buňkou závisí na tom, zda převáží stimulační nebo inhibiční signály Cytotoxická granula obsahující perforin a granzymy (perforin vytváří póry v cytoplazmatické membráně napadené buňky,v některých případech může dojít k osmotické lýze napadené buňky, vytvořenými póry se do buňky dostávají granzymy, které aktivují caspázy, což vede k apoptóze napadené buňky Fas-ligand (FasL) – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. TNFa
 – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. TNFa.")
7
Interferony Patří k humorální složce nespecifických mechanismů
IFNa - produkován virem napadenými lymfocyty, monocyty a makrofágy IFNb - produkován virem infikovanými fibroblasty a epiteliemi IFNa a IFNb – váží se na receptory na povrchu infikovaných a zdravých bb. a navozují v nich antivirový stav (syntéza enzymů, které blokují replikaci viru v buňce) IFNg – produkován TH1 buňkami, má regulační funkci, aktivuje makrofágy a stimuluje expresi MHCgp
 IFNg – produkován TH1 buňkami, má regulační funkci, aktivuje makrofágy a stimuluje expresi MHCgp.")
8
Bazofily a mastocyty a jejich význam v imunitních reakcích

9
Mastocyty ( žírné buňky)
Slizniční mastocyty – ve sliznicích dýchacího a gasrtointestinálního traktu, produkují histamin, serotonin, heparin, tryptázu,leukotrien C4…, účastní se při parazitózách a při alergiích Pojivové mastocyty – v pojivové tkáni, produkují tryptázu, chymázu, PGD2…, jsou zmnoženy při fibróze, při parazitózách a alergiích se neúčastní

10
Funkce mastocytů obrana proti parazitárním infekcím
za patologických okolností jsou zodpovědné za časný typ přecitlivělosti (imunopatologická reakce typu I) regulace imunitní odpovědi uplatňují se při zánětu, při angiogenezi, při remodelaci tkání podílejí se na udržování fyziologických funkcí sliznic přispívají k normálnímu metabolismu pojivových tkání komunikace mezi imunitním a nervovým systémem
 regulace imunitní odpovědi. uplatňují se při zánětu, při angiogenezi, při remodelaci tkání. podílejí se na udržování fyziologických funkcí sliznic. přispívají k normálnímu metabolismu pojivových tkání. komunikace mezi imunitním a nervovým systémem.")
11
Schema aktivace mastocytu
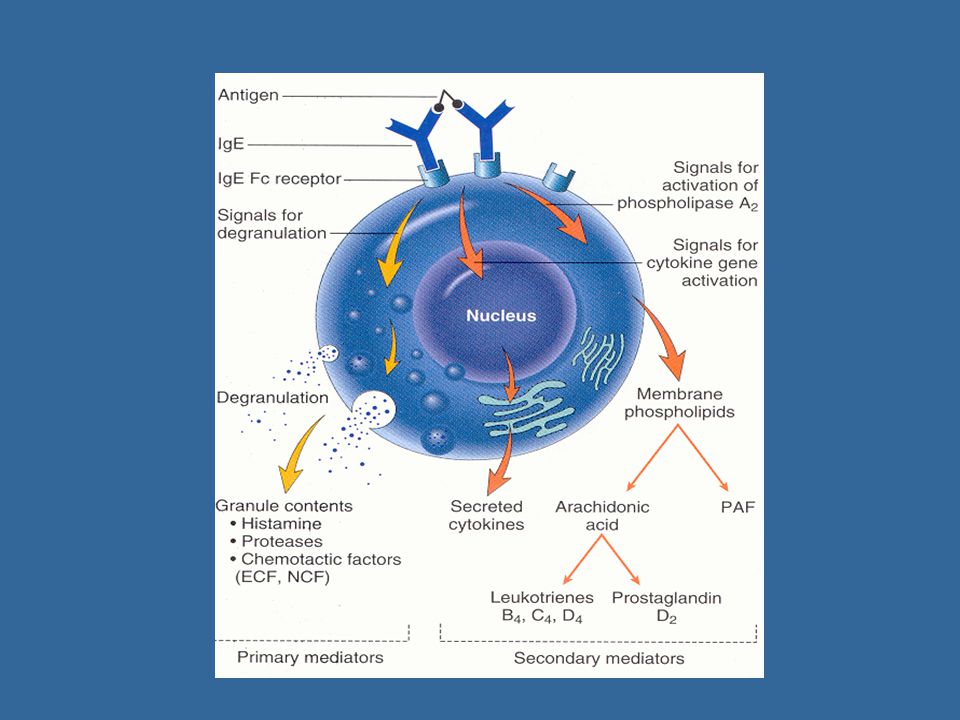
Žírné buňky mohou být stimulovány k degranulaci prostřednictvím přímého poškození (opiáty, alkohol, a některá antibiotika), propojením Fc receptorů pro IgE, nebo anafylatoxiny (C3a, C5a) Po navázání multivalentního antigenu ( mnohobuněčného parazita) pomocí IgE na vysokoafinní Fc receptor pro IgE (FcRI) dojde k agregaci několika molekul FcRI Iniciace degranulace mastocytu ( fúze cytoplazmatických granulí s povrchovou membránou a uvolnění jejich obsahu) Aktivace metabolismu kyseliny arachidonové (leukotrien C4, prostaglandin PGD2) Zahájení produkce cytokinů (TNF, TGF, IL-4,5,6…) Receptor pro komplementový fragment C5a (aktivace degranulace nezávislé na IgE)
, propojením Fc receptorů pro IgE, nebo anafylatoxiny (C3a, C5a) Po navázání multivalentního antigenu ( mnohobuněčného parazita) pomocí IgE na vysokoafinní Fc receptor pro IgE (FcRI) dojde k agregaci několika molekul FcRI. Iniciace degranulace mastocytu ( fúze cytoplazmatických granulí s povrchovou membránou a uvolnění jejich obsahu) Aktivace metabolismu kyseliny arachidonové (leukotrien C4, prostaglandin PGD2) Zahájení produkce cytokinů (TNF, TGF, IL-4,5,6…) Receptor pro komplementový fragment C5a (aktivace degranulace nezávislé na IgE)")
13
Sekreční produkty mastocytů
cytoplazmatická granula: hydrolytické enzymy, proteoglykany (heparin, chondroitinsulfát), biogenní aminy (histamin,serotonin) Histamin způsobuje vasodilataci, zvýšení vaskulární permeability, erytém, edém, svědění, kontrakci hladké svaloviny bronchů, zvýšení peristaltiky střev, zvýšení sekrece hlenu slizničními žlázkami v respiračním traktu a GITu (napomáhá eliminaci parazita) Metabolity kys. arachidonové (leukotrien C4, prostaglandin PGD2) Cytokiny (TNF, TGF, IL-4,5,6…)
, biogenní aminy (histamin,serotonin) Histamin způsobuje vasodilataci, zvýšení vaskulární permeability, erytém, edém, svědění, kontrakci hladké svaloviny bronchů, zvýšení peristaltiky střev, zvýšení sekrece hlenu slizničními žlázkami v respiračním traktu a GITu (napomáhá eliminaci parazita) Metabolity kys. arachidonové (leukotrien C4, prostaglandin PGD2) Cytokiny (TNF, TGF, IL-4,5,6…)")
14
Úloha mastocytů při rozvoji alergické reakce

15
Bazofily diferencují se z myeloidního prekurzoru
bývají považovány za cirkulující formu mastocytů receptorovou výbavou, obsahem granul, mechanismy stimulace a funkcemi jsou velmi podobné mastocytům jsou zodpovědné za vznik anafylaktického šoku

16
HLA systém (MHC glykoproteiny)
")
17
MHC glykoproteiny I. třídy (major histocompatibility complex)
Funkcí MHC gpI je prezentace peptidových fragmentů, které jsou produkovány buňkou (včetně virových, pokud jsou přítomny), na buněčném povrchu tak, aby byly rozpoznávány T lymfocyty (cytotoxickými CD8) Přítomny na všech jaderných buňkách organismu 3 izotypy klasických lidských MHC gp. ( HLA - A, -B, -C ) 3 izotypy neklasických MHC gp. ( HLA – E, -F, -G; molekuly CD1)
, na buněčném povrchu tak, aby byly rozpoznávány T lymfocyty (cytotoxickými CD8) Přítomny na všech jaderných buňkách organismu. 3 izotypy klasických lidských MHC gp. ( HLA - A, -B, -C ) 3 izotypy neklasických MHC gp. ( HLA – E, -F, -G; molekuly CD1)")
18
Struktura MHC gp I MHC gp. I. třídy se skládají z transmembránového řetězce a a nekovalentně asociovaného b2mikrotubulinu Řetězec a má 3 domény, 2 N-terminální (a1, a2 – vazebné místo pro peptidy) a 1 C-terminální doménu (a3 – zakotvena v cytoplazmatické membráně, struktura podobná imunoglobulinové doméně) Vazebné místo pro peptid je strukturně význačná rýha, jejíž dno je tvořeno b strukturou a boky jsou ohraničeny 2 a šroubovicemi Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu
 a 1 C-terminální doménu (a3 – zakotvena v cytoplazmatické membráně, struktura podobná imunoglobulinové doméně) Vazebné místo pro peptid je strukturně význačná rýha, jejíž dno je tvořeno b strukturou a boky jsou ohraničeny 2 a šroubovicemi. Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu.")
19
Vazba peptidů na MHC gp I
MHC gp I váží peptidy o délce 8 až 10 AK Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv (rozhodující jsou AK poblíž konců peptidu) K vazbě endogenních peptidů dochází v endoplazmatickém retikulu během biosyntézy MHC gp. Po vytvoření řetězce a a b2mikrotubulinu dochází v ER k poskládání do správné konformace a k vzájemné asociaci a k asociaci vhodného peptidu, tento komplex je dále zpracován v Golgiho aparátu a pak je prezentován na buněčném povrchu Navázané peptidy pocházejí z proteinů degradovaných proteazómem, který štěpí cytoplasmatické proteiny určené k likvidaci (označené ubiquitinem), peptidové fragmenty jsou transportovány do ER pomocí specifických membránových pump
 K vazbě endogenních peptidů dochází v endoplazmatickém retikulu během biosyntézy MHC gp. Po vytvoření řetězce a a b2mikrotubulinu dochází v ER k poskládání do správné konformace a k vzájemné asociaci a k asociaci vhodného peptidu, tento komplex je dále zpracován v Golgiho aparátu a pak je prezentován na buněčném povrchu. Navázané peptidy pocházejí z proteinů degradovaných proteazómem, který štěpí cytoplasmatické proteiny určené k likvidaci (označené ubiquitinem), peptidové fragmenty jsou transportovány do ER pomocí specifických membránových pump.")
20
Neklasické MHC gp. I HLA – E, -F, -G; molekuly CD1 Strukturně podobné klasickým MHC gp Jsou méně polymorfní Vyskytují se jen na některých buňkách Specializují se na vazbu zvláštních ligandů

21
HLA-E a HLA-G - vyskytují se na buňkách trofoblastu
Komplexy HLA-E a HLA-G s peptidy jsou rozpoznávány inhibičními receptory NK buněk a přispívají k toleranci plodu v děloze Molekuly CD1 - váží glykolipidy či jiné silně hydrofobní látky, tyto komplexy jsou rozpoznávány specializovanými ab T lymfocyty (NK-T lymfocyty)
")
22
MHC glykoproteiny II. třídy
Funkcí MHC gp II je prezentace peptidových fragmentů z proteinů pohlcených buňkou tak, aby byly rozpoznatelné T lymfocyty (pomocnými CD4) Vyskytují se na APC ( dendritické buňky, monocyty, makrofágy, B lymfocyty) 3 izotypy MHC gp II ( DR, DQ, DP )
 Vyskytují se na APC ( dendritické buňky, monocyty, makrofágy, B lymfocyty) 3 izotypy MHC gp II ( DR, DQ, DP )")
23
Struktura MHC gp II MHC gp. II se skládají ze 2 nekovalentně asociovaných transmembránových podjednotek a a b Vazebné místo pro peptid je tvořeno N-terminálními doménami a1 a b1 Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu

24
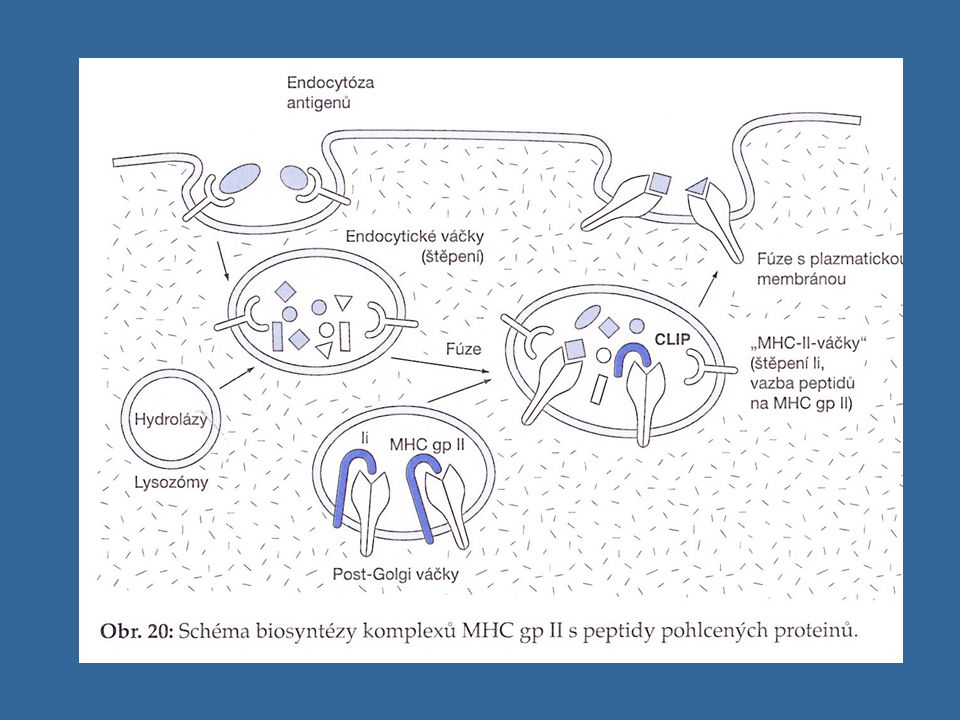
Vazba peptidů na MHC gp II
MHC gp II váží peptidy o délce 15 až 35 AK (ale i delší - vazebné místo pro peptid je na obou koncích otevřené) Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv Po vytvoření řetězce a a b v ER dochází k poskládání do správné konformace a k vzájemné asociaci a k připojení dalšího transmembránového řetězce, tzv. invariantního řetězce, který blokuje vazebné místo pro peptid, tento komplex je dále zpracován v Golgiho aparátu; sekreční váčky oddělené od GA fúzují s endozómy, poté se rozštěpí invariantní řetězce a do vazebného místa MHC gp se naváží peptidové fragmenty endocytovaných proteinů a poté je komplex prezentován na buněčném povrchu
 Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv. Po vytvoření řetězce a a b v ER dochází k poskládání do správné konformace a k vzájemné asociaci a k připojení dalšího transmembránového řetězce, tzv. invariantního řetězce, který blokuje vazebné místo pro peptid, tento komplex je dále zpracován v Golgiho aparátu; sekreční váčky oddělené od GA fúzují s endozómy, poté se rozštěpí invariantní řetězce a do vazebného místa MHC gp se naváží peptidové fragmenty endocytovaných proteinů a poté je komplex prezentován na buněčném povrchu.")
26
Polymorfismus MHC glykoproteinů
HLA komplex se nachází na chromozómu 6 U MHC gp je vysoký polymorfismus, tzn. existují až stovky různých alelických forem jednotlivých izotypů (kromě neklasických MHC gp. I a kromě DR řetězce a) Kodominantní dědičnost alelických forem (jedinec má na povrchu buněk 3 izotypy HLA molekul (HLA-A, -B, -C) většinou ve 2 různých alelických formách) Polymorfismus má ochranný význam na úrovni jedince i na úrovni populace Polymorfismus MHC gp způsobuje komplikace při transplantacích
 Kodominantní dědičnost alelických forem (jedinec má na povrchu buněk 3 izotypy HLA molekul (HLA-A, -B, -C) většinou ve 2 různých alelických formách) Polymorfismus má ochranný význam na úrovni jedince i na úrovni populace. Polymorfismus MHC gp způsobuje komplikace při transplantacích.")
27
HLA typizace = určení HLA antigenů na povrchu lymfocytů Provádí se při předtransplantačním vyšetření a při určení paternity 1) Sérologická typizace mikrolymfocytotoxický test allospecifická séra ( získaná od vícenásobných rodiček do 6 týdnů po porodu, získaná vakcinací dobrovolníků, nebo komerčně připravené sety typizačních sér (monoklonální protilátky)) princip - inkubace lymfocytů s typizačními séry za přítomnosti králičího komplementu, poté je přidáno vitální barvivo, které obarví mrtvé buňky buňky nesoucí určité HLA jsou usmrceny cytotoxickými Ab proti tomuto Ag, procento mrtvých buněk je mírou toxicity séra (síly a titru antileukocytárních protilátek) za pozitivní reakci se považuje více než 10% mrtvých bb. (sérologickou typizaci lze provádět i pomocí průtokové cytometrie)
 Sérologická typizace. mikrolymfocytotoxický test. allospecifická séra ( získaná od vícenásobných rodiček do 6 týdnů po porodu, získaná vakcinací dobrovolníků, nebo komerčně připravené sety typizačních sér (monoklonální protilátky)) princip - inkubace lymfocytů s typizačními séry za přítomnosti králičího komplementu, poté je přidáno vitální barvivo, které obarví mrtvé buňky - buňky nesoucí určité HLA jsou usmrceny cytotoxickými Ab proti tomuto Ag, procento mrtvých buněk je mírou toxicity séra (síly a titru antileukocytárních protilátek) za pozitivní reakci se považuje více než 10% mrtvých bb. (sérologickou typizaci lze provádět i pomocí průtokové cytometrie)")
28
2) Molekulárně genetické metody
Pro typizaci se používají hypervariabilní úseky ve II. exonu genů kódujících HLA II. třídy, pro určení HLA I. třídy se používá polymorfismus v II. a III. exonu kódujících genů 2a) PCR-SSP = polymerázová řetězová reakce se sekvenčními specifickými primery extrahovaná DNA slouží jako substrát v sadě PCR reakcí každá PCR reakce obsahuje primerový pár specifický pro určitou alelu (resp. skupinu alel) pozitivní a negativní reakce se hodnotí elektroforézou každá kombinace alel má svůj specifický elektroforetický obraz
 PCR-SSP. = polymerázová řetězová reakce se sekvenčními specifickými primery. extrahovaná DNA slouží jako substrát v sadě PCR reakcí. každá PCR reakce obsahuje primerový pár specifický pro určitou alelu (resp. skupinu alel) pozitivní a negativní reakce se hodnotí elektroforézou. každá kombinace alel má svůj specifický elektroforetický obraz.")
29
2b) PCR-SSO = PCR reakce se sekvenčně specifickými oligonukleotidy namnoží se hypervariabilní úseky genů kódujících HLA hybridizace s enzymaticky nebo radioaktivně značenými DNA sondami specifickými pro jednotlivé alely 2c) PCR- SBT = sequencing based typing; sekvenování nejpřesnější metodika HLA typizace získáme přesnou sekvenci nukleotidů, kterou porovnáme s databází známých sekvencí HLA alel
 PCR- SBT. = sequencing based typing; sekvenování. nejpřesnější metodika HLA typizace. získáme přesnou sekvenci nukleotidů, kterou porovnáme s databází známých sekvencí HLA alel.")
30
Imunoglobuliny

31
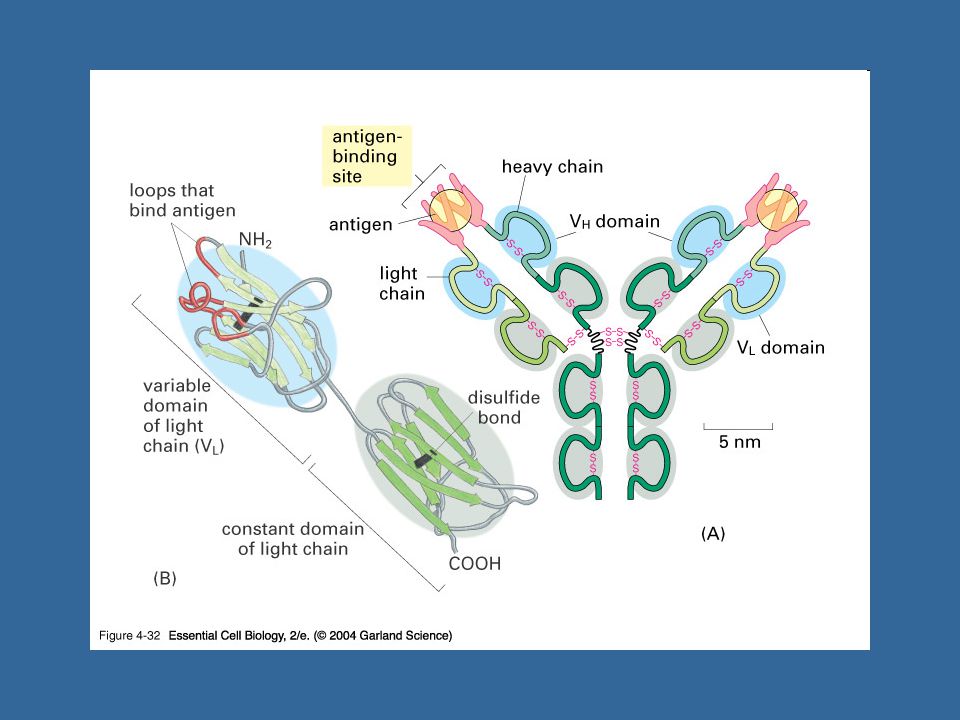
Struktura imunoglobulinů
2 těžké (H) řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l Typy H řetězců – m, d, g (g1-4), a (a1,a2), e
 řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec. H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l. Typy H řetězců – m, d, g (g1-4), a (a1,a2), e.")
32
Variabilní domény L a H řetězce tvoří vazebné místo pro Ag
Pantová oblast = místo, kde jsou těžké řetězce spojeny cystinovými můstky Fc části imunoglobulínů jsou silně glykosilovány, váží se na Fc receptory J řetězec Sekreční komponenta

34
Protilátka štěpená papainem Protilátka štěpená pepsinem

35
Funkce imunoglobulinů
Neutralizace Ag Aglutinace Ag Aktivace komplementu Opsonizace (IgM, IgG, IgE) Aktivace mastocytů prostřednictvím IgE ADCC
 Aktivace mastocytů prostřednictvím IgE. ADCC.")
36
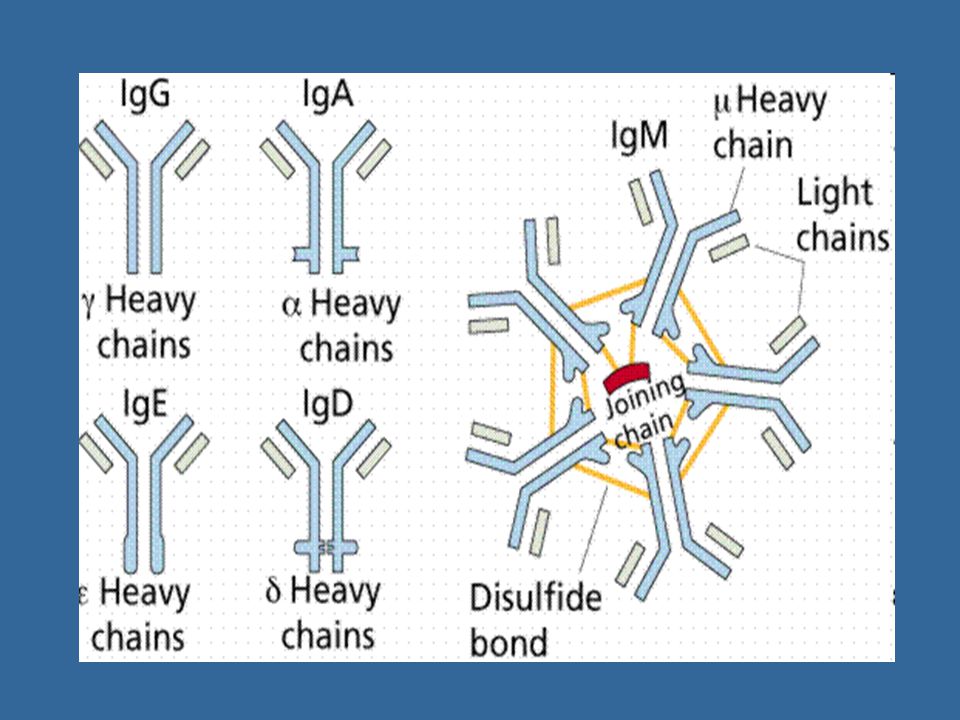
Třídy imunoglobulínů a jejich funkce
Rozlišujeme podle konstantní části H řetězce IgM, IgD, IgG ( IgG1 – IgG4), IgA (IgA1, IgA2), IgE IgM - jako monomer tvoří BCR sekretovaný ve formě pentamerů (10 vazebných míst) první izotyp, který se tvoří po setkání s Ag neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR v séru se nachází ve velmi malé koncentraci - (koncentrace 0,1 g/l; biol. poločas 3 dny)
, IgA (IgA1, IgA2), IgE. IgM - jako monomer tvoří BCR - sekretovaný ve formě pentamerů (10 vazebných míst) - první izotyp, který se tvoří po setkání s Ag - neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů - (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR - v séru se nachází ve velmi malé koncentraci. - (koncentrace 0,1 g/l; biol. poločas 3 dny)")
37
IgG - jednotlivé izotypy IgG1-IgG4 se liší schopností aktivovat komplement a vázat se na Fc receptory fagocytů (opsonizace) - funkce: neutralizace, opsonizace, aktivace komplementu - přestupuje placentou - tvořen při sekundární imunitní odpovědi (koncentrace 8 – 18 g/l; biol. poločas 21 dnů)
")
38
IgA - slizniční IgA - ochrana sliznic, neutralizace, opsonizace, neaktivuje komplement dimér se sekreční komponentou sliny, slzy, mateřské mléko sérový IgA – monomer, dimer či trimer (koncentrace 0,9 – 3,5 g/l; biol. poločas 6 dnů) IgE - uplatňuje se při obraně proti mnohobuněčným parazitům je hlavní příčinou alergických reakcí - (koncentrace 3x10-4 g/l; biol. poločas 2 dny)
")
40
Genetický základ tvorby imunoglobulinů

41
Genové segmenty pro H řetězce - chromozóm 14
V (variabilita) - několik set D (diversita) - asi 50 J (joining) – 9 C konstantní části H řetězce Genové segmenty pro L řetězce - k chromozóm 2 - l chromozóm 22 V (variabilita) J (joining) C konstantní části L řetězce Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk
 - několik set. D (diversita) - asi 50. J (joining) – 9. C konstantní části H řetězce. Genové segmenty pro L řetězce - k chromozóm 2. - l chromozóm 22. V (variabilita) J (joining) C konstantní části L řetězce. Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů. Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk.")
42
Přeskupování genů kódujících H řetězce
D-J přeskupení - vyštěpení úseku IgH mezi některým D a J segmentem (probíhá na obou chromozómech) V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.
 V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.")
43
Přeskupování genů kódujících L řetězce
Nejprve se přeskupují geny kódující L řetězec k, dochází k vyštěpování úseků mezi některým V a J segmentem (souběžně na obou chromozómech), pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.
, pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.")
44
Variabilita imunoglobulinů
Variabilita imunoglobulinů je dána: Různorodostí kombinací V(D)J segmentů Spojovací variabilitou – po vyštěpení genových úseků nejsou konce odstřiženy zcela přesně Enzym terminální transferáza – prodlužuje odstřižené konce o krátké náhodné tzv. N-sekvence Somatické mutace V segmentů přeskupených genů po kontaktu s Ag na povrchu FDC
J segmentů. Spojovací variabilitou – po vyštěpení genových úseků nejsou konce odstřiženy zcela přesně. Enzym terminální transferáza – prodlužuje odstřižené konce o krátké náhodné tzv. N-sekvence. Somatické mutace V segmentů přeskupených genů po kontaktu s Ag na povrchu FDC.")
45
Izotypový přesmyk (class switch)
Dochází k němu během terminální diferenciace B lymfocytu po aktivaci Ag na povrchu FDC (nezbytný kostimulační signál přes CD40) Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů (tato sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň) a vyštěpují úseky genů Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce
 Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů (tato sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň) a vyštěpují úseky genů. Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce.")
47
Izotypový přesmyk (class switch) - pokračování
Cytokiny regulují k jakému izotypovému přesmyku dojde: IL-4 stimuluje přesmyk na IgE a IgG1, IgG4 TGFb stimuluje přesmyk na IgG2 a IgA Regulace, zda půjde o sekretovanou či membránovou formu probíhá na úrovni mRNA (na 3´konci C segmentu jsou za sekvencemi kódujícími secernovanou formu sekvence pro membránovou formu)
")
48
Antiidiotypové protilátky
IDIOTOP = každá determinanta variabilní části protilátky, může být součástí paratopu, či může ležet mimo paratop IDIOTYP = soubor idiotopů; souhrn identických vazebných struktur pro Ag na protilátkách stejné specifity Idiotypické struktury protilátek 1. generace jsou rozpoznávány některými B lymfocyty jako antigeny a mohou se proti nim tvořit tzv. antiidiotypové protilátky (protilátky 2. generace; některá vazebná místa mohou připomínat Ag, který vyvolal tvorbu protilátek 1. generace) Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi
 Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi.")
49
B lymfocyty

50
B lymfocyty B-lymfocyty (B buňky) jsou buňky zodpovědné především za specifickou, protilátkami zprostředkovanou imunitní odpověď. Mají rovněž velký význam pro imunitní paměť (využívá se při očkování). B-lymfocyty rozpoznávají nativní antigen pomocí BCR (B cell receptor) Příslušný B-lymfocyt, na jehož receptorech došlo k vazbě antigenu, je stimulován k pomnožení a diferenciaci na efektorové neboli plazmatické bb., které produkují velké množství protilátek stejné specifity, jako je BCR (jde vlastně o tentýž protein v rozpustné formě), vážou se tedy na stejný antigen. Z části stimulovaných B-lymfocytů se diferencují paměťové buňky.
 jsou buňky zodpovědné především za specifickou, protilátkami zprostředkovanou imunitní odpověď. Mají rovněž velký význam pro imunitní paměť (využívá se při očkování). B-lymfocyty rozpoznávají nativní antigen pomocí BCR (B cell receptor) Příslušný B-lymfocyt, na jehož receptorech došlo k vazbě antigenu, je stimulován k pomnožení a diferenciaci na efektorové neboli plazmatické bb., které produkují velké množství protilátek stejné specifity, jako je BCR (jde vlastně o tentýž protein v rozpustné formě), vážou se tedy na stejný antigen. Z části stimulovaných B-lymfocytů se diferencují paměťové buňky.")
51
Povrchové znaky B lymfocytů
CD 10 - nezralý B lymfocyt CD 19 - charakteristický povrchový znak B lymfocytů CD 20 - na povrchu Ig-pozitivních B lymfocytů IgM, IgD - BCR MHC gp II.třídy - Ag prezentující molekuly CD 40 – kostimulační receptor

52
Vývoj B lymfocytů Vývoj B lymfocytů probíhá v kostní dřeni a dokončuje se posetkání s Ag v sekundárních lymfatických orgánech. Pluripotentní hematopoetická kmenová buňka Progenitor B lymfocytu → zahájení rekombinačních procesů, které vedou ke vzniku velkého množství klonů B lymfocytů s individuálně specifickými BCR Pre B lymfocyt → exprese pre-B receptoru (tvořen H(m) řetězcem a náhradním L řetězcem) Nezralý B lymfocyt → exprese povrchového IgM (BCR); v této fázi vývoje dochází k eliminaci autoreaktivních klonů Zralý B lymfocyt → exprese povrchového IgM a IgD (BCR)
 řetězcem a náhradním L řetězcem) Nezralý B lymfocyt → exprese povrchového IgM (BCR); v této fázi vývoje dochází k eliminaci autoreaktivních klonů. Zralý B lymfocyt → exprese povrchového IgM a IgD (BCR)")
53
Kritické momenty při vývoji B lymfocytů
Dokončení přeskupování genů pro H řetězce a povrchová exprese pre-BCR Úspěšné přeskupení genů pro L řetězec a povrchová exprese IgM (BCR) Testování nezralých B lymfocytů, zda nejsou autoreaktivní Dalším kritickým stádiem procházejí při zahájení somatických mutací a afinitní maturaci, kdy přežívají pouze B lymfocyty s nejvyšší afinitou k antigenu.
 Testování nezralých B lymfocytů, zda nejsou autoreaktivní. Dalším kritickým stádiem procházejí při zahájení somatických mutací a afinitní maturaci, kdy přežívají pouze B lymfocyty s nejvyšší afinitou k antigenu.")
54
BCR BCR se skládá z povrchového imunoglobulinu (IgM, IgD – H řetězce jsou transmembránové ; rozeznává Ag) a asociovaných signalizačních molekul (Iga a Igb), které jsou asociovány s cytoplazmatickými protein tyrosin-kinázami (PTK) skupiny Src Po současném navázání Ag na 2 či více BCR dojde k přiblížení PTK, vzájemné fosforylaci a fosforylaci dalších cytoplazmatických proteinů, což vede ke změnám transkripce genů, proliferaci, diferenciaci a sekreci protilátek Signály spuštěné vazbou Ag na BCR mohou být zesíleny spoluprací s CR2, který váže C3dg (opsonin)
 a asociovaných signalizačních molekul (Iga a Igb), které jsou asociovány s cytoplazmatickými protein tyrosin-kinázami (PTK) skupiny Src. Po současném navázání Ag na 2 či více BCR dojde k přiblížení PTK, vzájemné fosforylaci a fosforylaci dalších cytoplazmatických proteinů, což vede ke změnám transkripce genů, proliferaci, diferenciaci a sekreci protilátek. Signály spuštěné vazbou Ag na BCR mohou být zesíleny spoluprací s CR2, který váže C3dg (opsonin)")
55
Eliminace autoreaktivních klonů B lymfocytů
Při náhodném přeskupováním genů, spojovacích nepřesnostech, párování H-L a somatických mutací mohou vzniknout i klony B lymfocytů nesoucí autoreaktivní receptory a produkující autoreaktivní protilátky. Většina autoreaktivních B lymfocytů je eliminována na úrovni nezralých B lymfocytů (v kostní dřeni), jestliže svým BCR váží autoantigen s dostatečnou afinitou, obdrží signál vedoucí k apoptotické smrti. Pokud touto eliminací projdou některé autoreaktivní klony, jejich autoreaktivita se většinou neprojeví, protože k jejich aktivaci chybí příslušné TH lymfocyty, mnohé autoantigeny jsou kryptické, či se vyskytují v malé koncentraci a jsou imunitním systémem ignorovány.
, jestliže svým BCR váží autoantigen s dostatečnou afinitou, obdrží signál vedoucí k apoptotické smrti. Pokud touto eliminací projdou některé autoreaktivní klony, jejich autoreaktivita se většinou neprojeví, protože k jejich aktivaci chybí příslušné TH lymfocyty, mnohé autoantigeny jsou kryptické, či se vyskytují v malé koncentraci a jsou imunitním systémem ignorovány.")
56
Setkání B lymfocytu s Ag v sekundárních lymfatických orgánech

57
Ontogeneze tvorby protilátek
Syntéza specifických protilátek začíná kolem týdne gestace, celková koncentrace IgA a IgM zůstává až do porodu neměřitelná, IgG se začínají tvořit až po porodu B lymfocyty na imunizaci reagují převážně tvorbou IgM, přesmyk na jiné izotypy je pomalejší Pozvolný nárůst tvorby vlastních IgG za poklesu mateřských IgG (kolem 3.-6.měs.)
")
58
Ontogeneze tvorby protilátek
Koncentrace IgM dosahuje hodnot srovnatelných s dospělými v 1.-3.roce života, IgG+A mezi r. Protilátková reakce na polysacharidové antigeny se objevuje až kolem 2.roku života Ve stáří je slabší protilátková odpověď na nové podněty a vyšší produkce autoprotilátek

Podobné prezentace


















>")

