Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Calvinův cyklus a pentosafosfátová dráha.
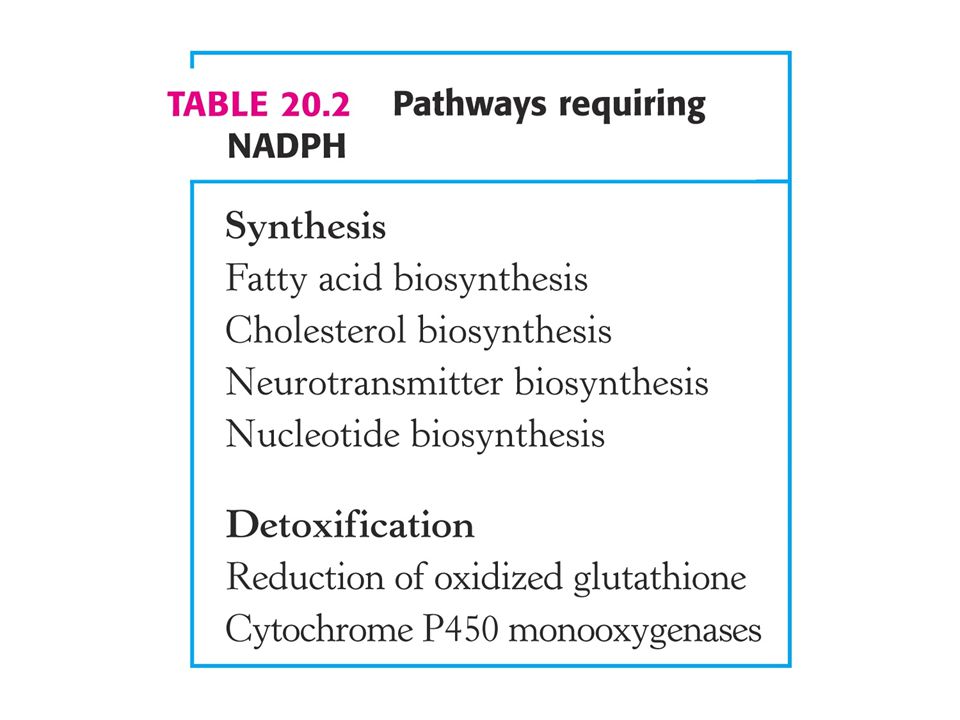
V Calvinově cyklu se syntetizují hexosy z CO2 a vody. Pentosafosfátová dráha vede ke tvorbě NADPH a syntéze pentos Meatabolismus glukosa-6-fosfátu je spojovacím článkem s glykolýzou. Glukosa-6-fosfátdehydrogenasa hraje ústřední roli při ochraně před reaktivními kyslíkatými radikály.

2
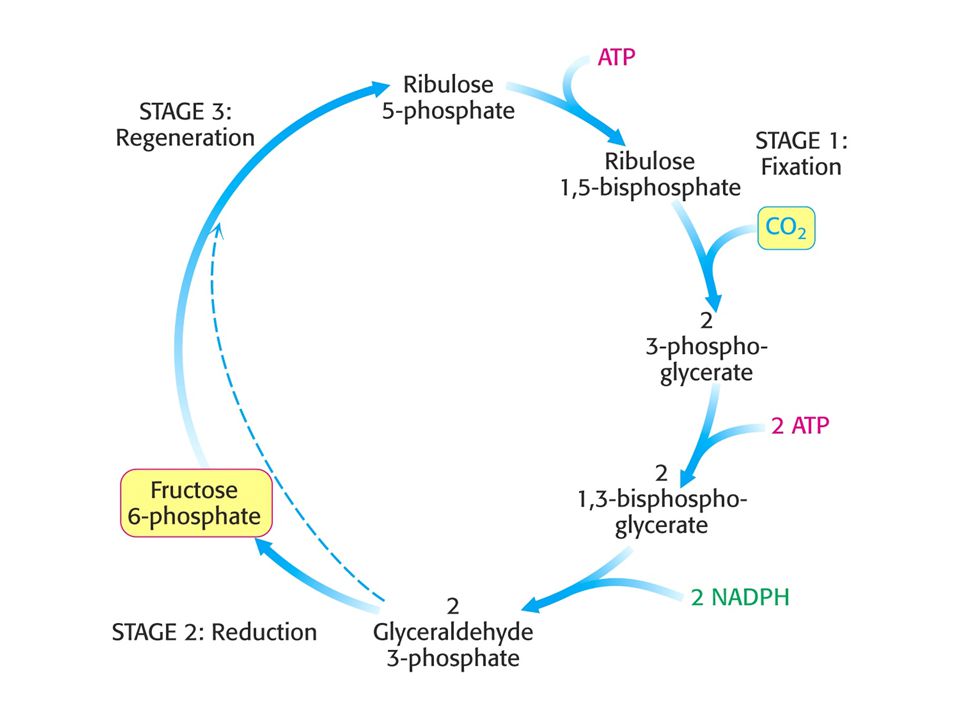
Stupně Calvinova cyklu.
Fixace CO2 ribulosa-1,5-bisfosfátem za vzniku dvou molekul 3-fosfogylcerátu. Redukce 3-fosfoglycerátu za tvorby hexos. Regenerace ribulosa-1,5-bisfosfátu, aby mohl být fixován další oxid uhličitý.

3
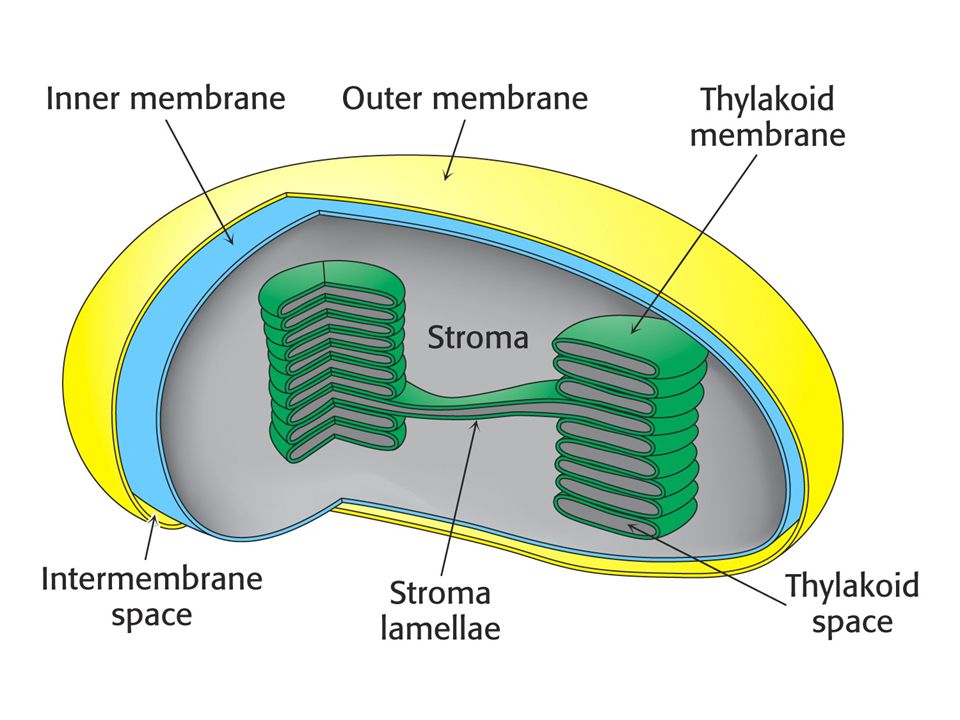
Chloroplasty – místo fotosyntézy
Chloroplasty – místo fotosyntézy. Vstup CO2 do ribulosa-1,5-bisfosfátu na na membráně thylakoidu obrácené ke stromatu a celý Calvinův cyklus ve stromatu.

7
Tvorba 3-fosfoglycerátu zaznamenaná pomocí 14CO2.

8
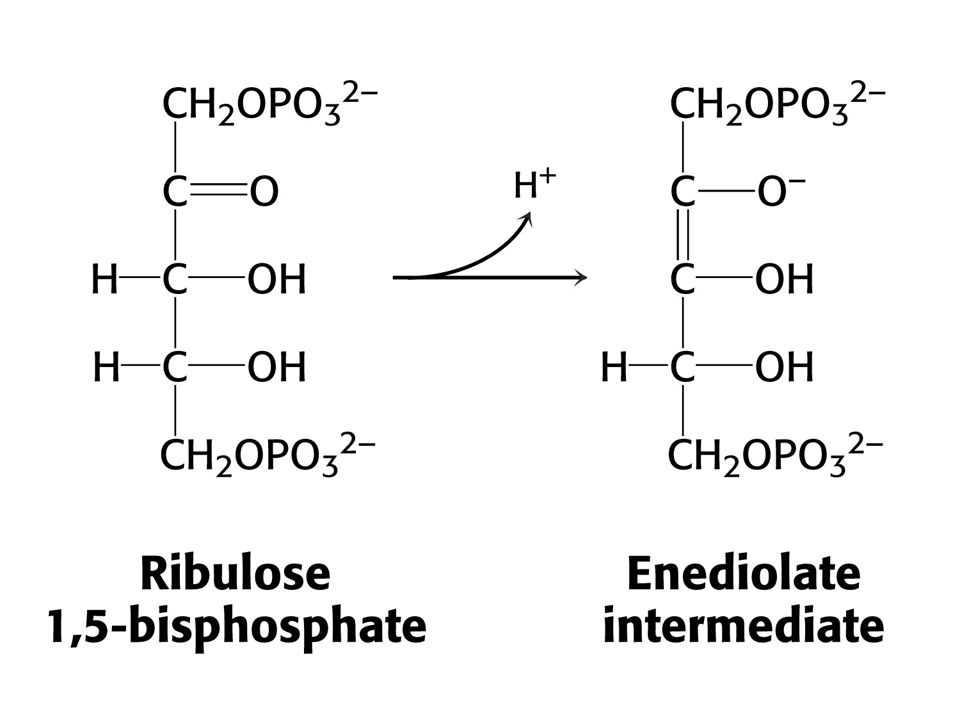
Ribulosa-1,5-bisfosfátkarboxylasa/oxygenasa (RUBISCO)
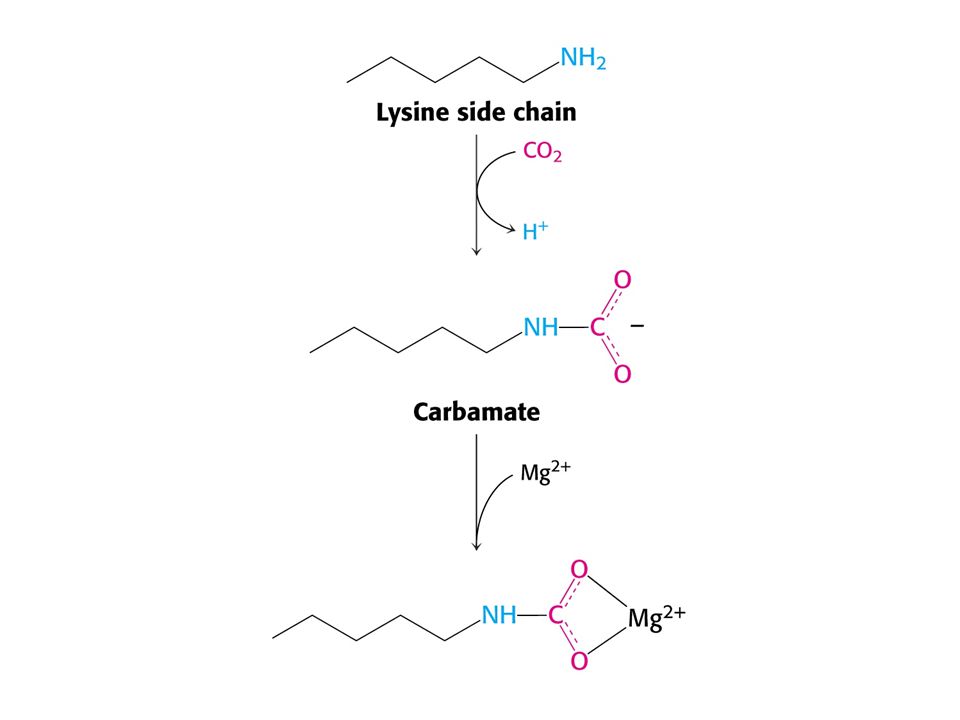
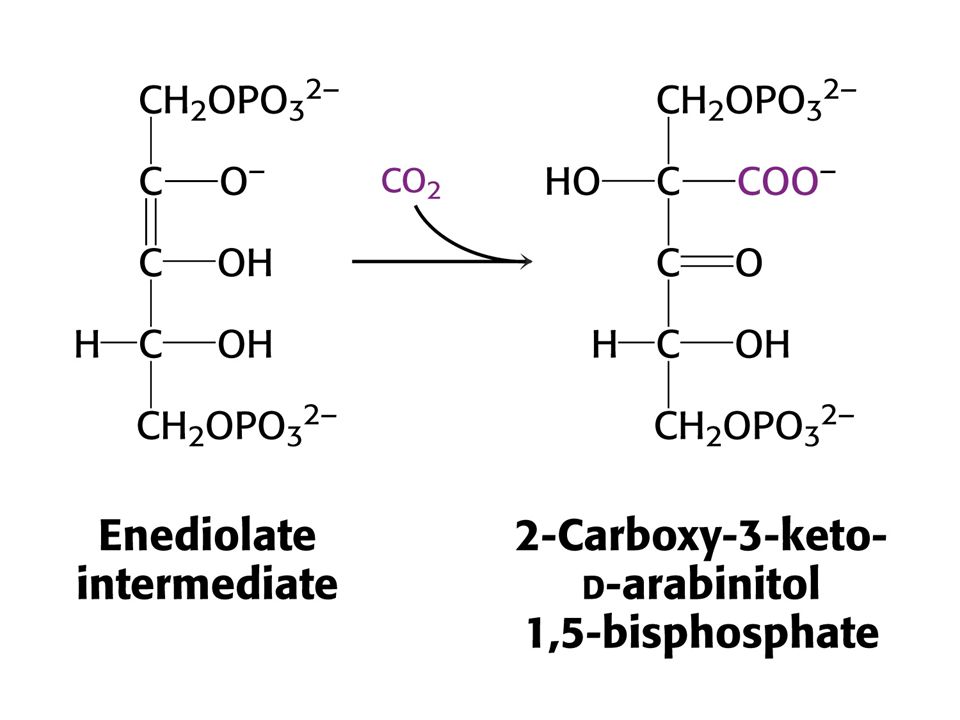
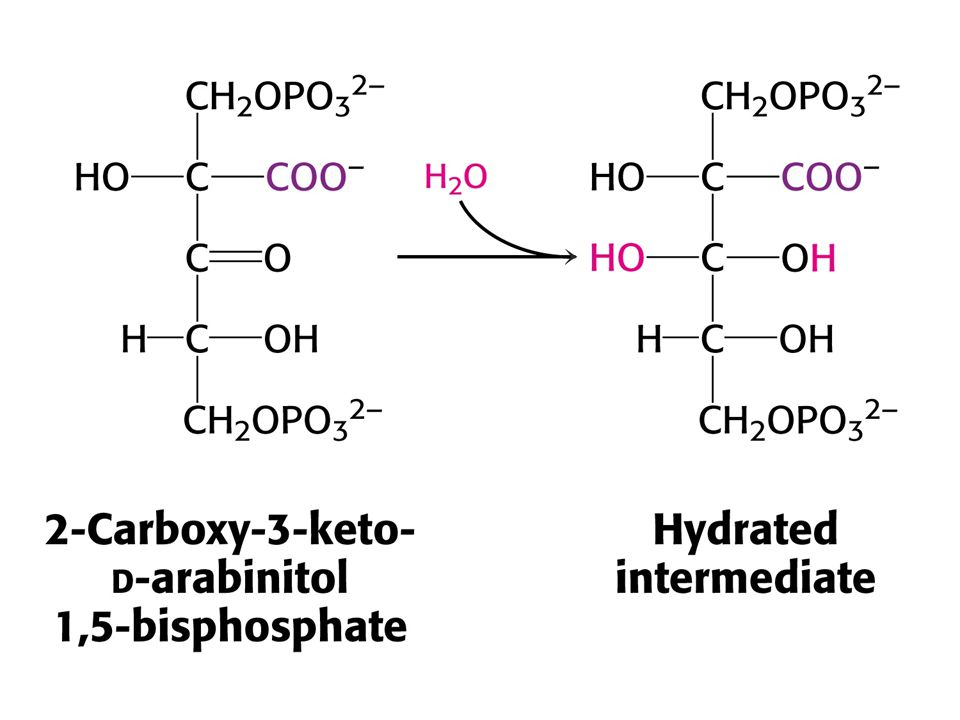
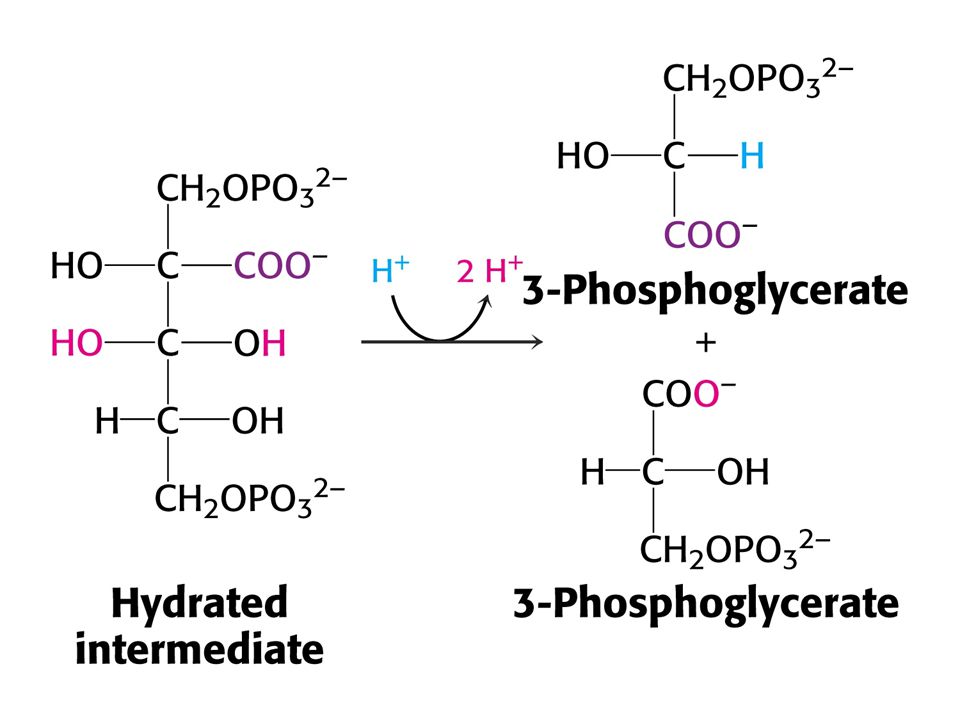
Vstup CO do ribulosa-1,5-bisfosfát je silně exergonní reakcí kJ/mol !!! (Standardní). RUBISCO v chloroplastech se skládá z 8 velkých podjednotek (L, 55-kd) a osmi malých podjednotek (S, 13 kd). Každá L podjednotka obsahuje katalytické a regulační místo. Podjednotky S zvyšují katalytickou aktivitu RUBISCa. V aktivním místě enzymu je nutný Mg2+, který stabilizuje kladným nábojem záporný náboj substrátu. Pro vazbu Mg2+ je nutný další vázaný CO2 na vedlejší řetězec Lys v poloze 201 ve formě karbamátu. Enzymu je v chloroplastech 17 % celkových proteinů. Nejvíce zastoupený enzym v biosféře. Katalytická rychlost je nízká, pouze 3 s-1
. RUBISCO v chloroplastech se skládá z 8 velkých podjednotek (L, 55-kd) a osmi malých podjednotek (S, 13 kd). Každá L podjednotka obsahuje katalytické a regulační místo. Podjednotky S zvyšují katalytickou aktivitu RUBISCa. V aktivním místě enzymu je nutný Mg2+, který stabilizuje kladným nábojem záporný náboj substrátu. Pro vazbu Mg2+ je nutný další vázaný CO2 na vedlejší řetězec Lys v poloze 201 ve formě karbamátu. Enzymu je v chloroplastech 17 % celkových proteinů. Nejvíce zastoupený enzym v biosféře. Katalytická rychlost je nízká, pouze 3 s-1.")
9
Struktura RUBISCO. Složeno z osmi podjednotek.

11
Role hořečnatých iontů v RUBISCu.

17
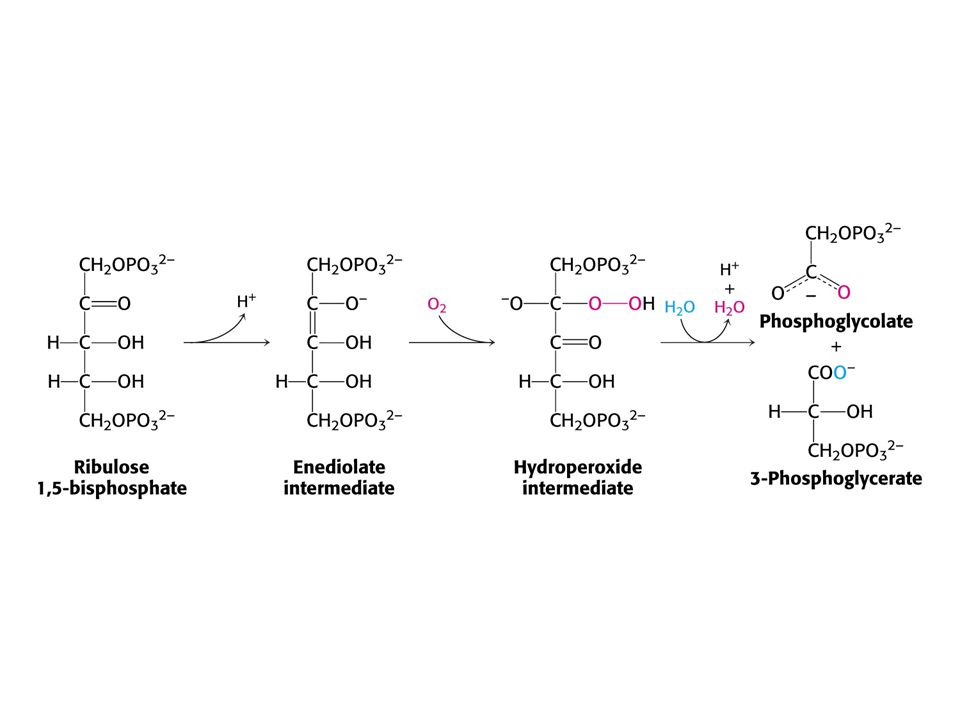
OXYGENASOVÁ REAKCE RUBISCA.

18
Rychlost karboxylasové reakce je 4 x vyšší než oxygenasové za normálních atmosférických podmínek při 25oC. Koncentrace CO2 ve stromatu je 10 mM a kyslíku 250 mM. Oxygenasová reakce, stejně jako karboxylasová, potřebuje Lys 201 v karbamátové formě. Ta však vzniká jen za přítomnosti CO2. Nepřítomnost CO2 tak chrání RUBISCO před oxygenasovou reakcí.

20
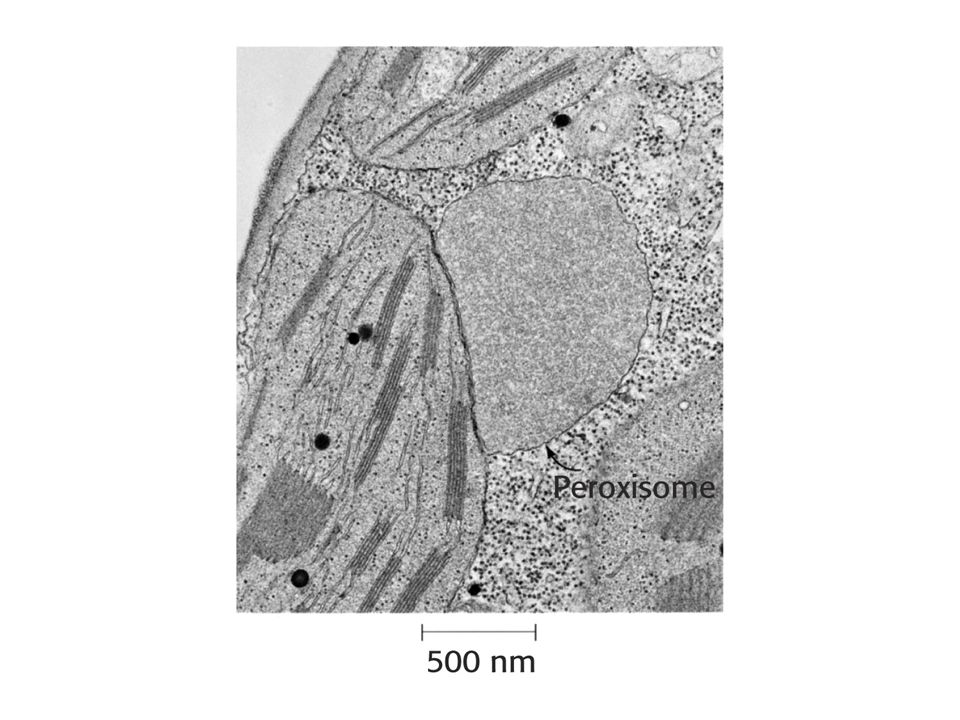
Fosfoglykolát vzniká při oxygenasové reakci v chloroplastu.
Po defosforylaci je glykolát transportován do peroxisomů, kde je převeden na glyoxylát a poté glycin. V mitochondrii jsou dva Gly převedeny na Ser. Dochází k dekarboxylaci a uvolnění amoniaku. Amoniak je využit v chloroplastu.

21
Fotorespirační reakce.

23
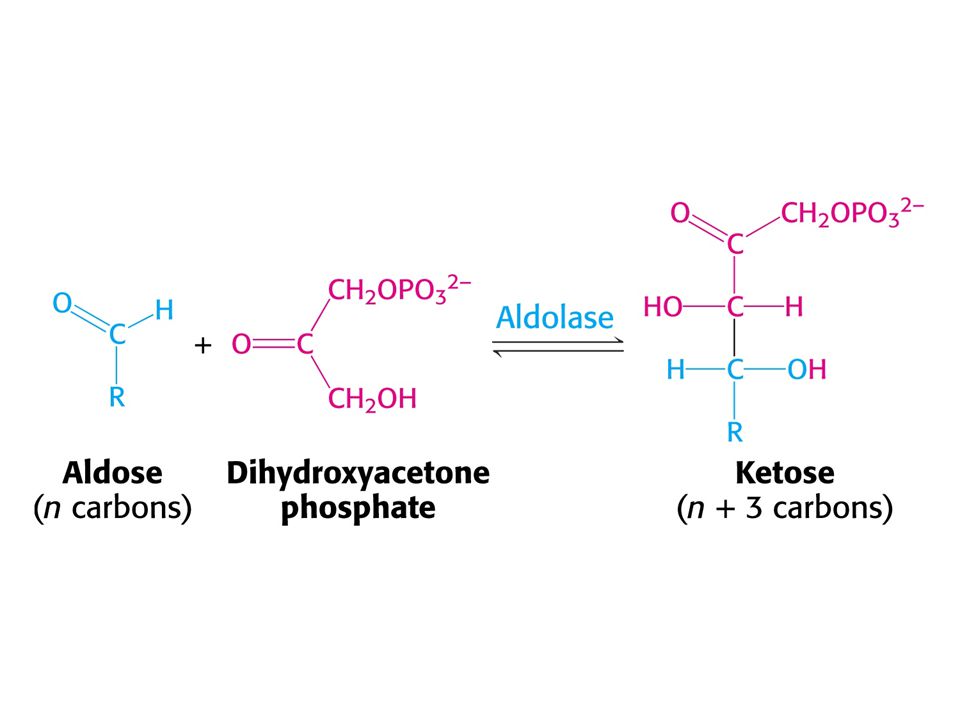
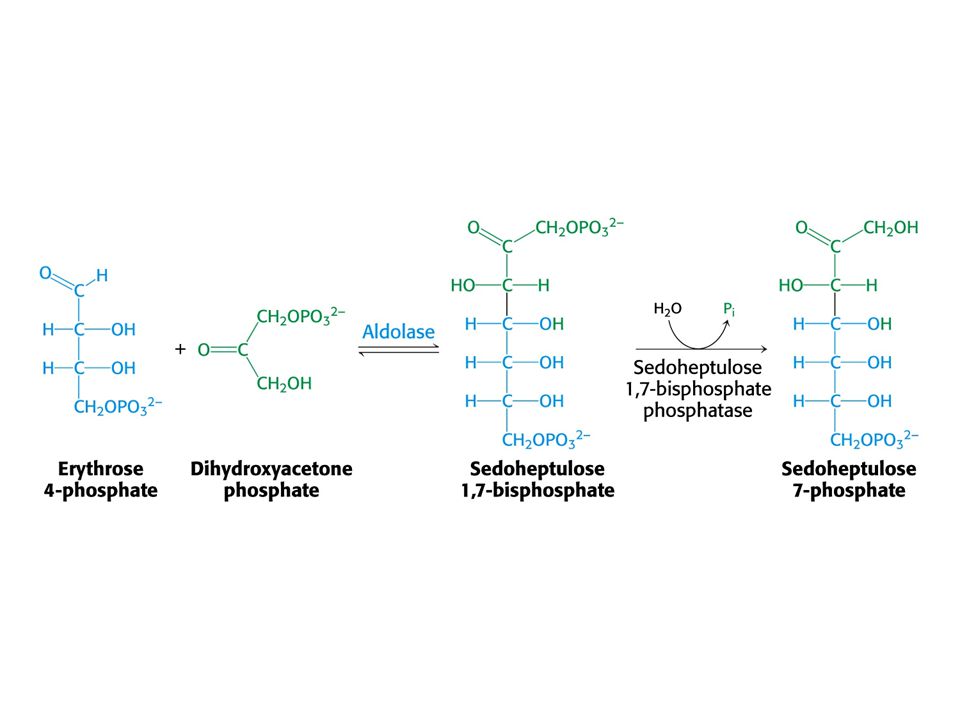
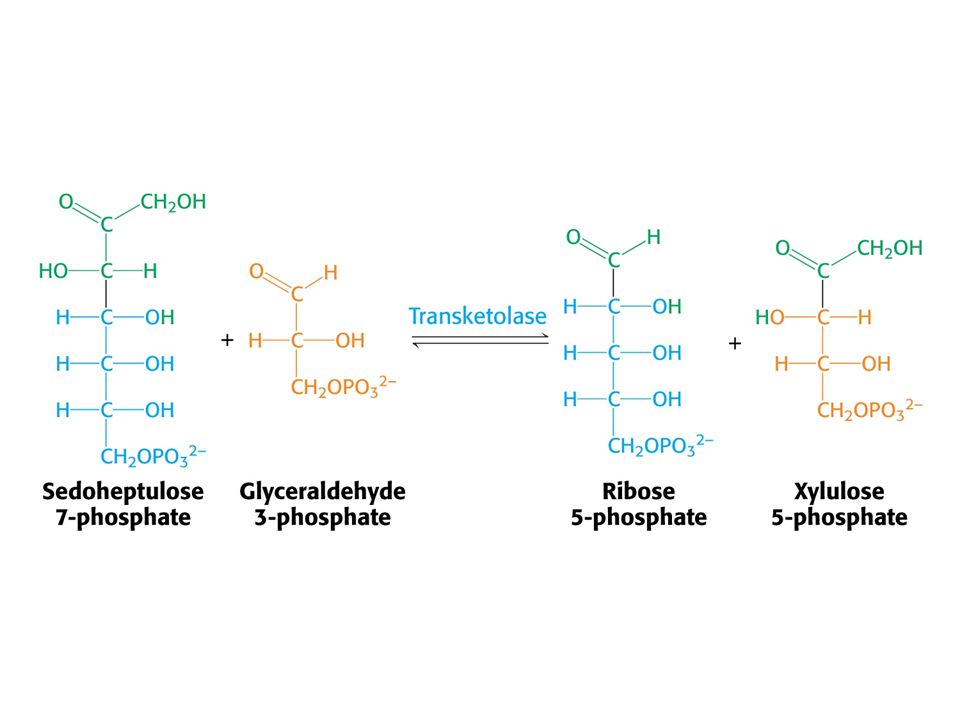
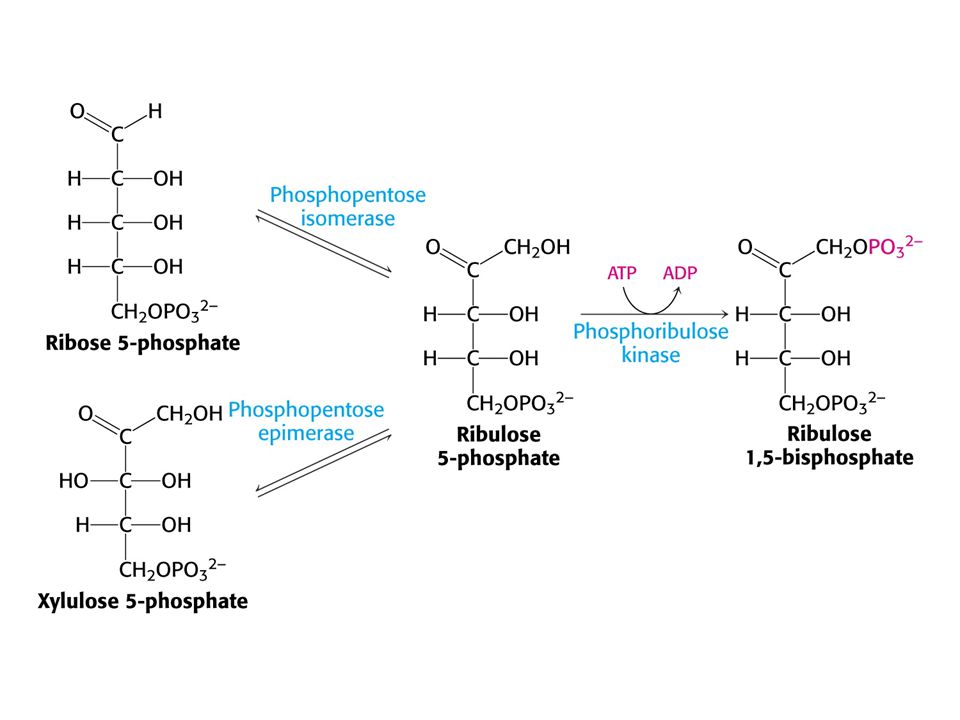
Tvorba hexosafosfátu.

31
Calvinův cyklus.

32
Stechiometrie Calvinova cyklu.
Rovnovážná rovnice Calvinova cyklu: 6 CO ATP + 12 NADPH + 12 H2O = C6H12O ADP + 18 Pi + 12 NADP+ + 6 H+

33
Škrob a sacharosa jsou hlavními skladovatelnými sacharidy rostlin.
Sacharosa je syntetizována v cytosolu. Rostliny nejsou schopny transportovat hexosafosfáty přes membránu chloroplastu. Existuje translokátor triosafosfátů, který je transportuje v synportu s fosfátem. Vytvořená fruktosa-6-fosfát se spojuje s UDP-glukosou. Po hydrolýze fosfátu se vzniklá sacharosa ukládá. Škrob se syntetizuje v chloroplastech, aktivující složkou je UDP-glukosa. Polymer obsahuje větvení a-1,6-glykosidových vazeb.

34
Syntéza sacharosa-6-fosfátu.

35
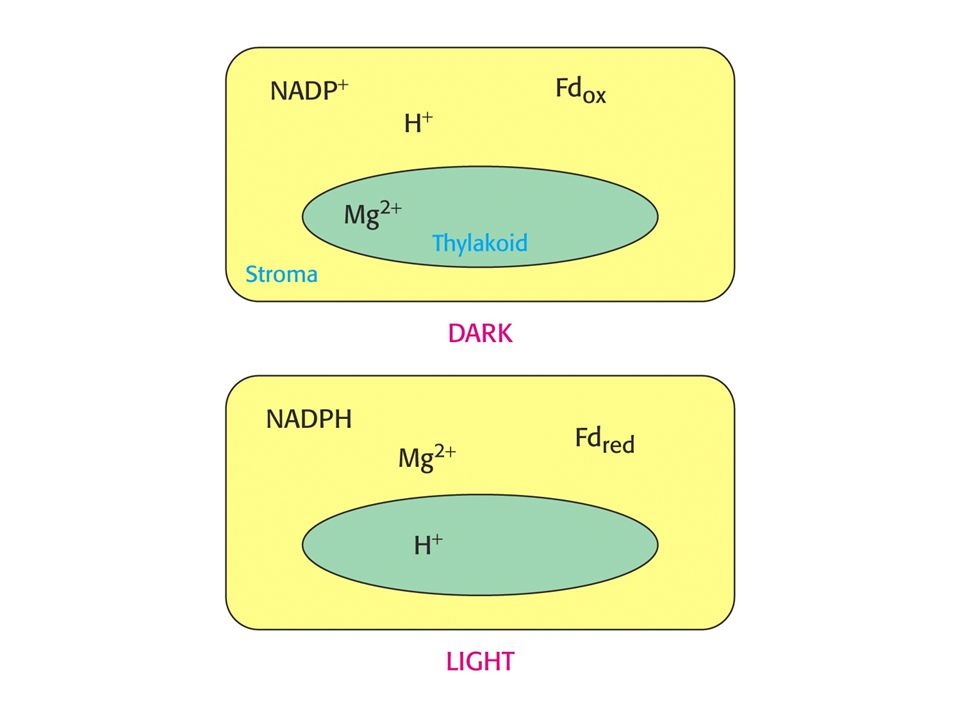
Regulace aktivit Calvinova cyklu.
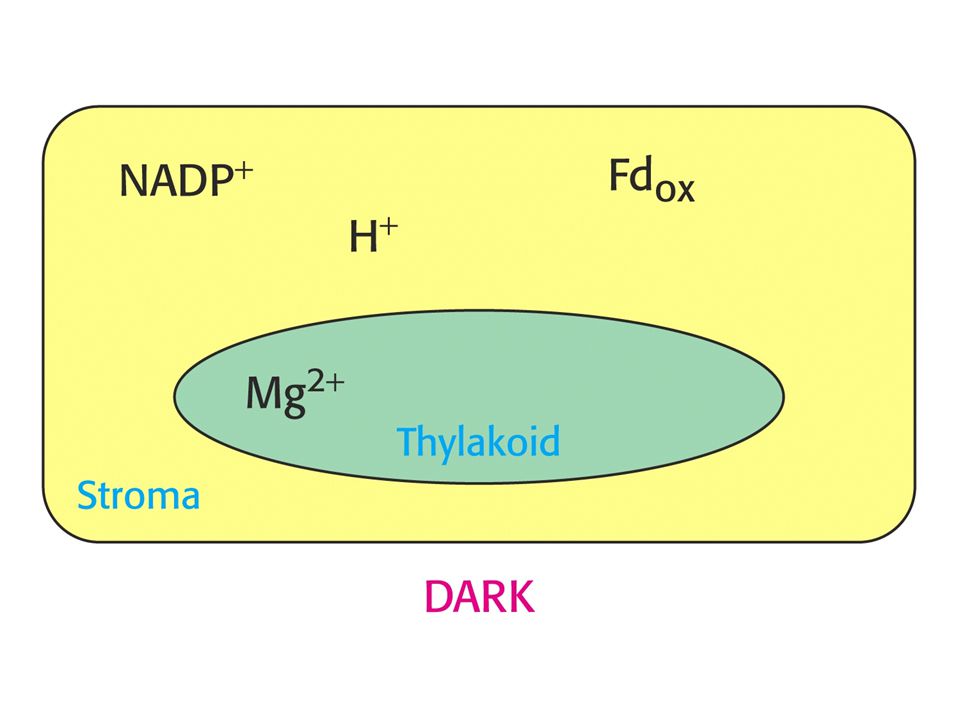
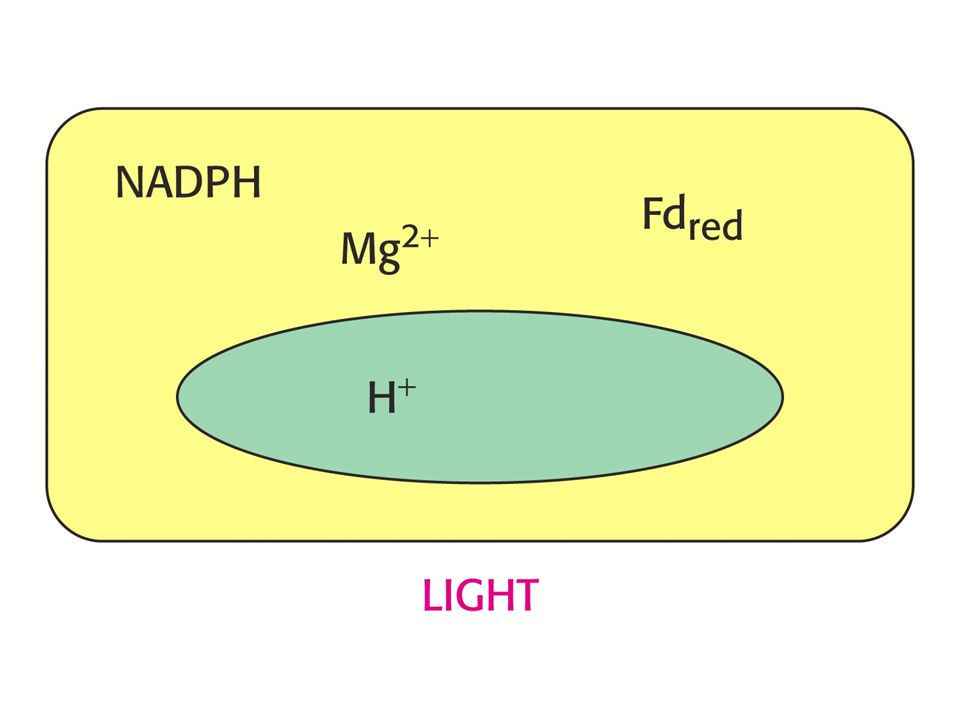
RUBISCO je aktivováno světlem vyvolanými změnami v koncentraci protonů a Mg2+. Tvorba karbamátu na Lys 201 rubisca je iniciována zvýšením pH a vysokou koncentrací Mg2+ ve stromatu. Obojí je spojeno se světlem. Protony jsou pumpovány ze stromatu do prostoru thylakoidů a koncentrace Mg2+ ve stromatu roste jako kompenzace odpumpovaných protonů.

36
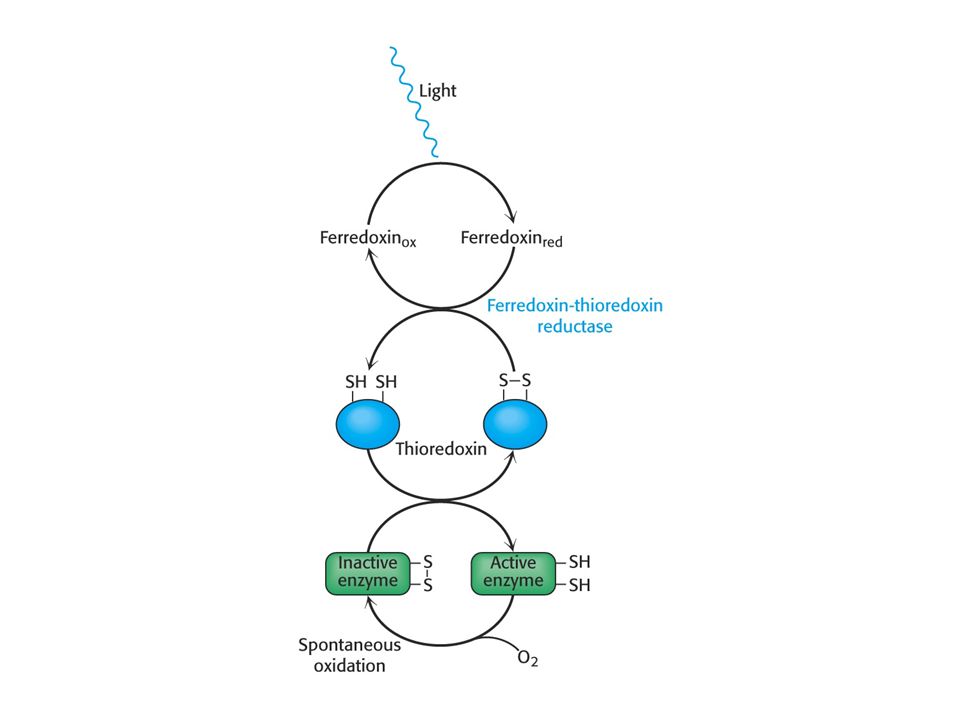
Thioredoxin je protein podílející se na tvorbě redukovaného ferredoxinu a NADPH. Thioredoxin je redukován redukovaným ferredoxinem. Existuje ve dvou formách mezi redukovanými (sulfhydrylovými) a oxidovanými (disulfidovými) skupinami. Redukovaná forma aktivuje celou řadu enzymů Calvinova cyklu.
 a oxidovanými (disulfidovými) skupinami. Redukovaná forma aktivuje celou řadu enzymů Calvinova cyklu.")
40
V OXIDOVANÉM STAVU a REDUKOVANÉM STAVU.
THIOREDOXIN V OXIDOVANÉM STAVU a REDUKOVANÉM STAVU. Redukovaný THIOREDOXIN aktivuje řadu enzymů metabolismu sacharidů a fenylalaninamoniumlyasu (biosyntéza fenylpropanoidů-lignin)
")
44
Svrchní epidermis Spodní Svazky cévní Podpůrné buňky Průduchy Houbový parenchym Palisádový

45
Svrchní epidermis Listový mezofyl Spodní Svazky cévní Podpůrné buňky Průduchy

46
C4 rostliny. Objasněno M.D.Hatchem a C.R.Slackem
Oxaloacetát a malát jsou sloučeniny, které přenáší oxid uhličitý z mezofylových buněk do buněk cévních svazků. Fosfoenolpyruvátkarboxylasa, NADPH-malátdehydrogenasa. Tvorba fosfoenolpyruvátu enzymem pyruvát-Pidikinasou.

47
Metabolismu C4.

48
Energeticky transport CO2:
CO2 v mezofylu + ATP + voda = CO2 v cévních svazcích + AMP + 2 Pi + H+ C4 + Calvinův cyklus: 6 CO ATP + 12 NADPH + 12 H2O = C6 H12 O ADP + 30 Pi + 12 NADP H+ Spotřebovává se o 12 molekul ATP více než u rostlin C3 Rostliny C3 převládají v mírném pásmu do teplot 28oC a C4 při teplotách vyšších.

49
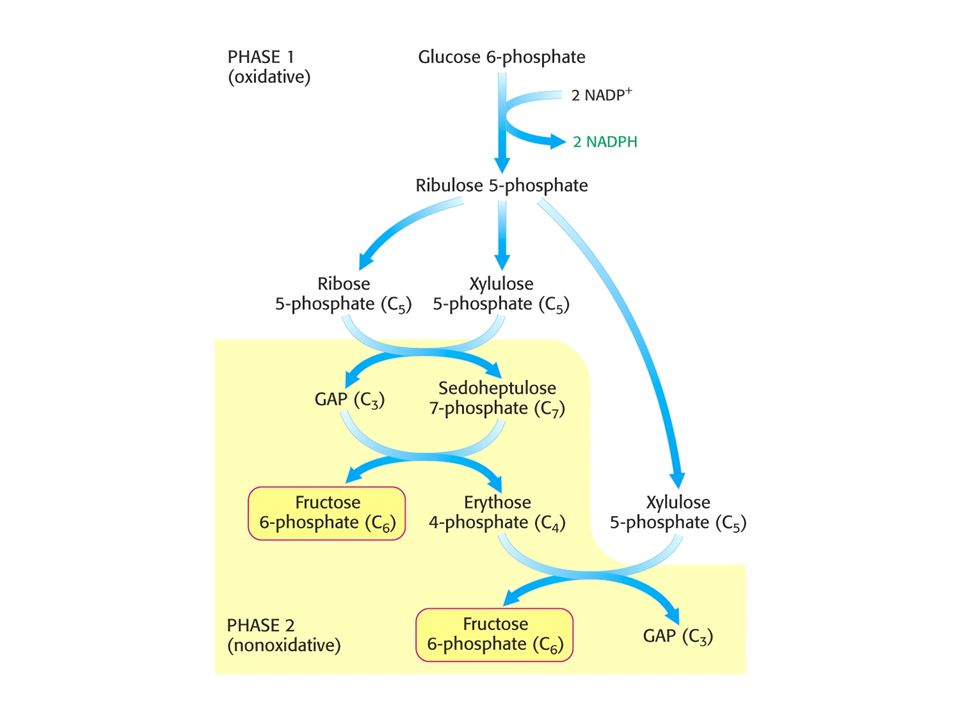
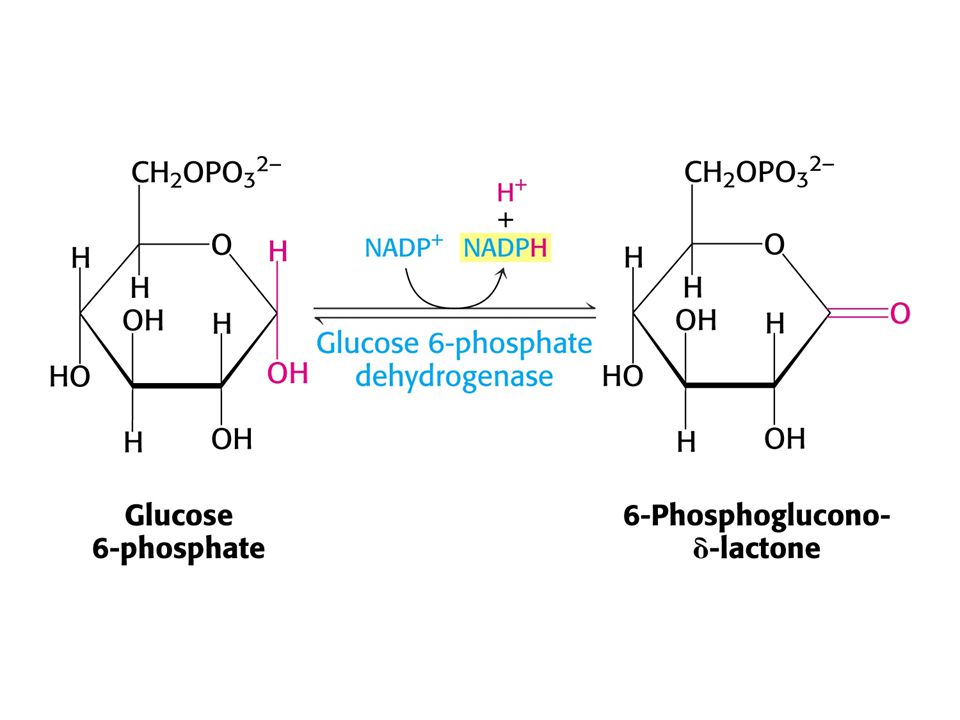
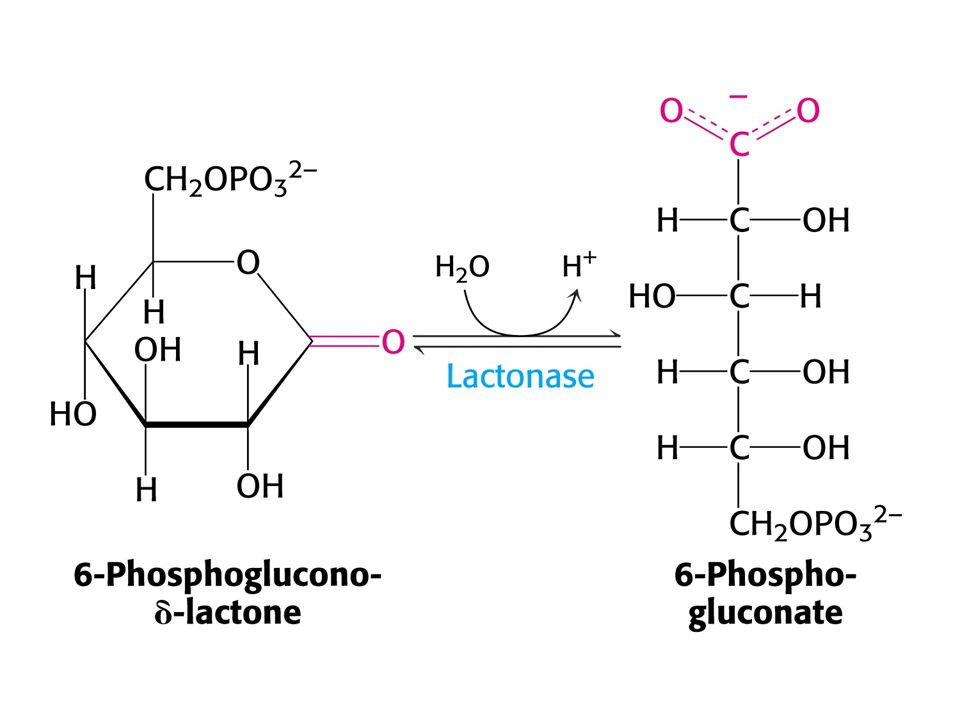
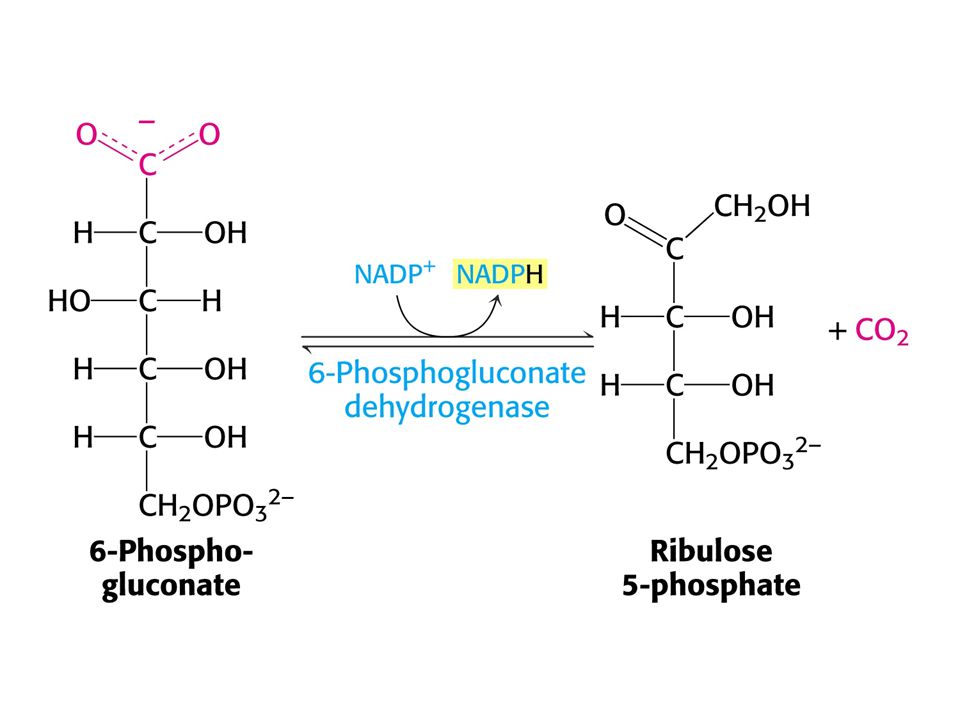
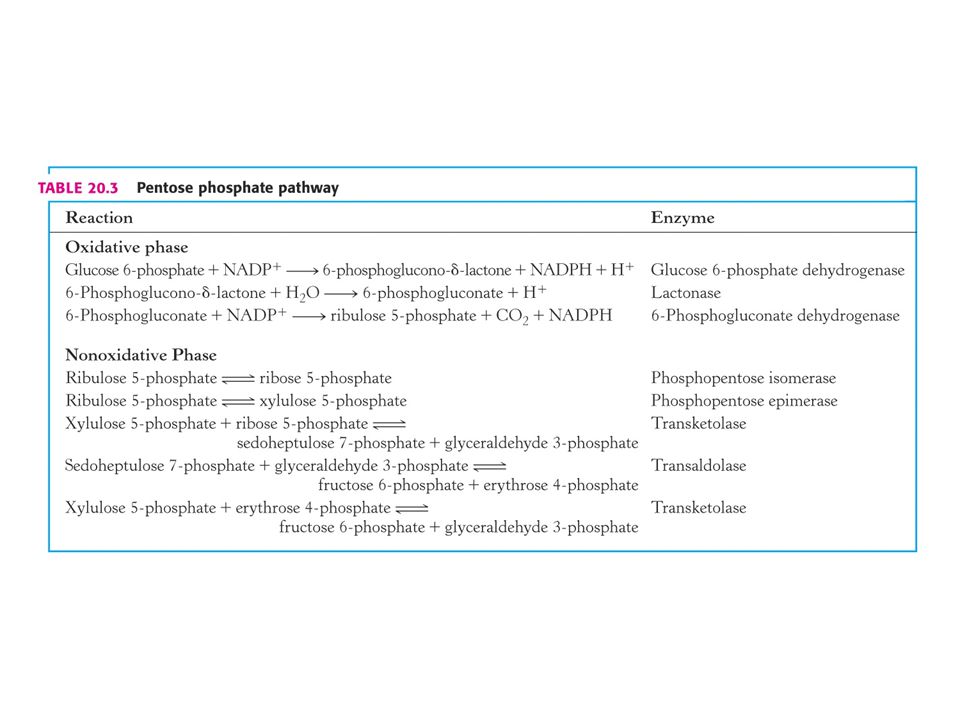
PENTOSAFOSFÁTOVÝ METABOLISMUS nebo pentosafosfátová dráha (PFM)
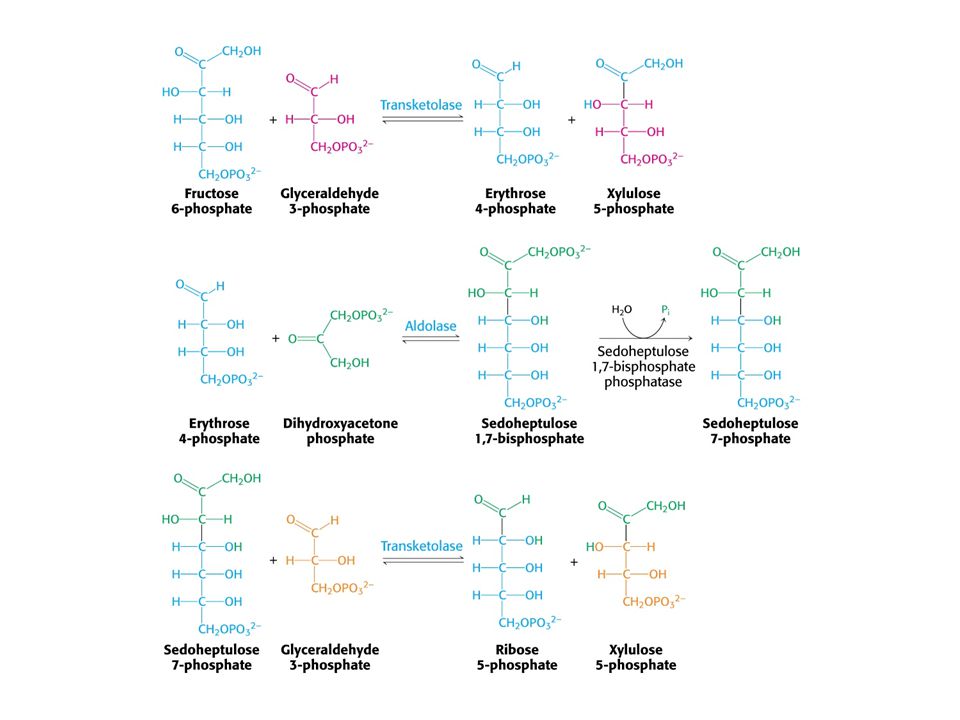
V buňkách poměr [NAD+] / [NADH] = znamená metabolické oxidace. Poměr [NADP+] / [NADPH] = 0, 01 –znamená metabolické redukce. Dvě části PFM: Obě v cytosolu. Oxidativní – dehydrogenasová Neoxidativní – přeměny tří-, čtyř-, pěti-, šesti- a sedmiuhlíkatých sacharidů. Výsledkem je tvorba pentos pro biosyntézu nukleotidů, případně přeměna nadbytku pentos na meziprodukty glykolýzy.

60
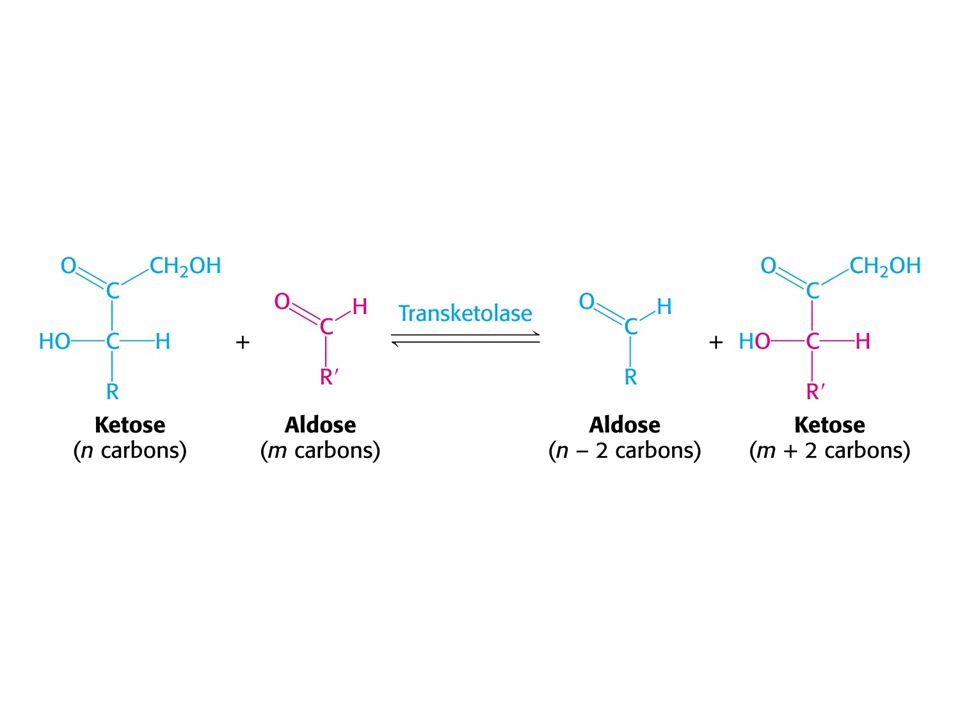
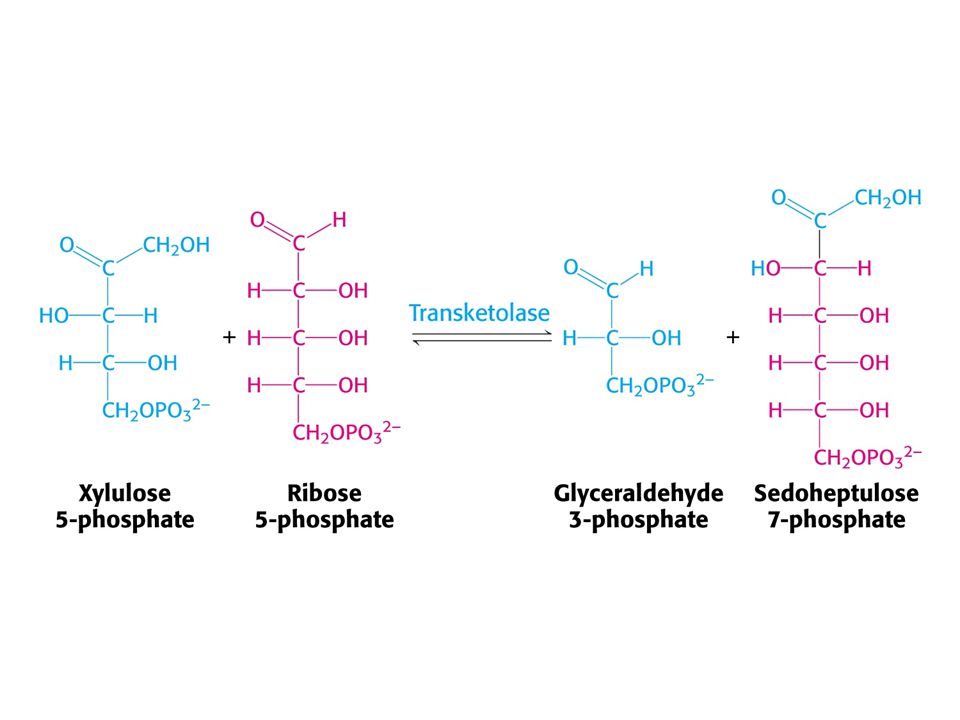
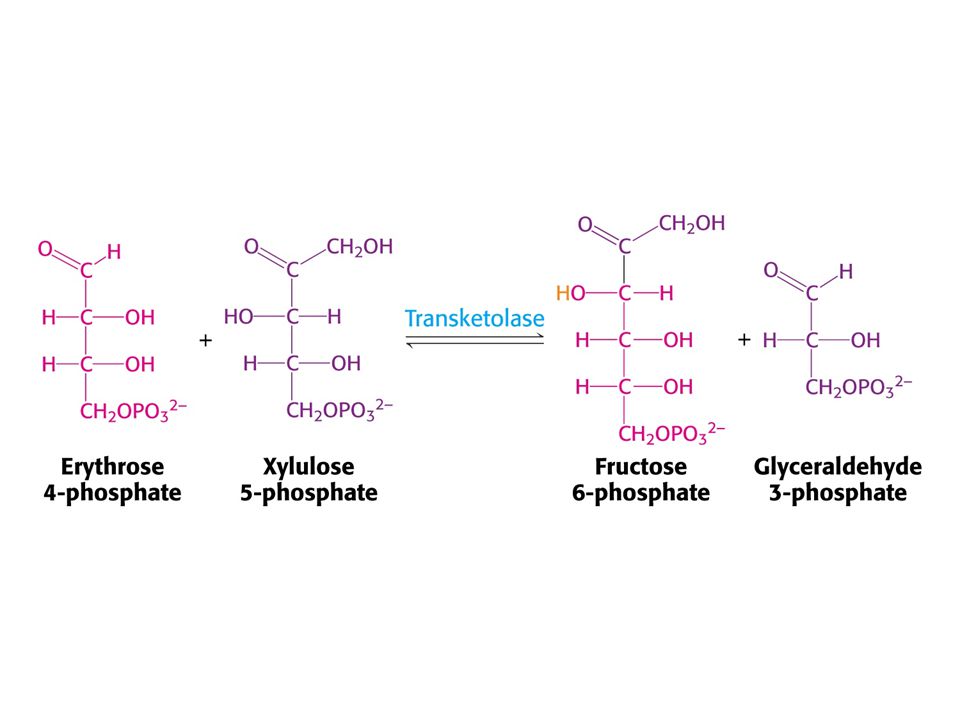
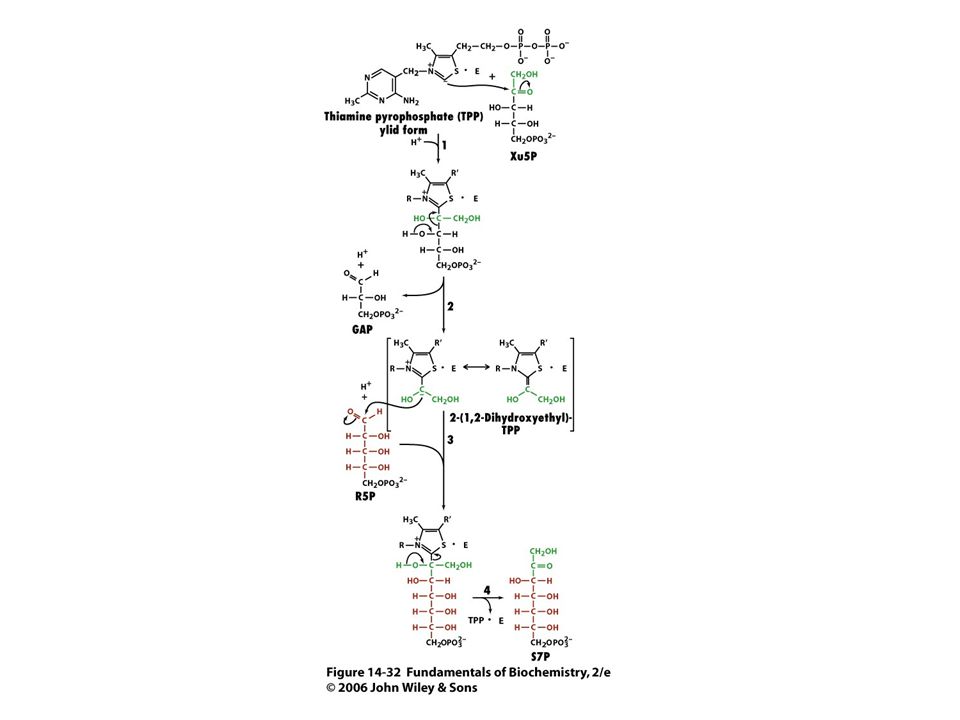
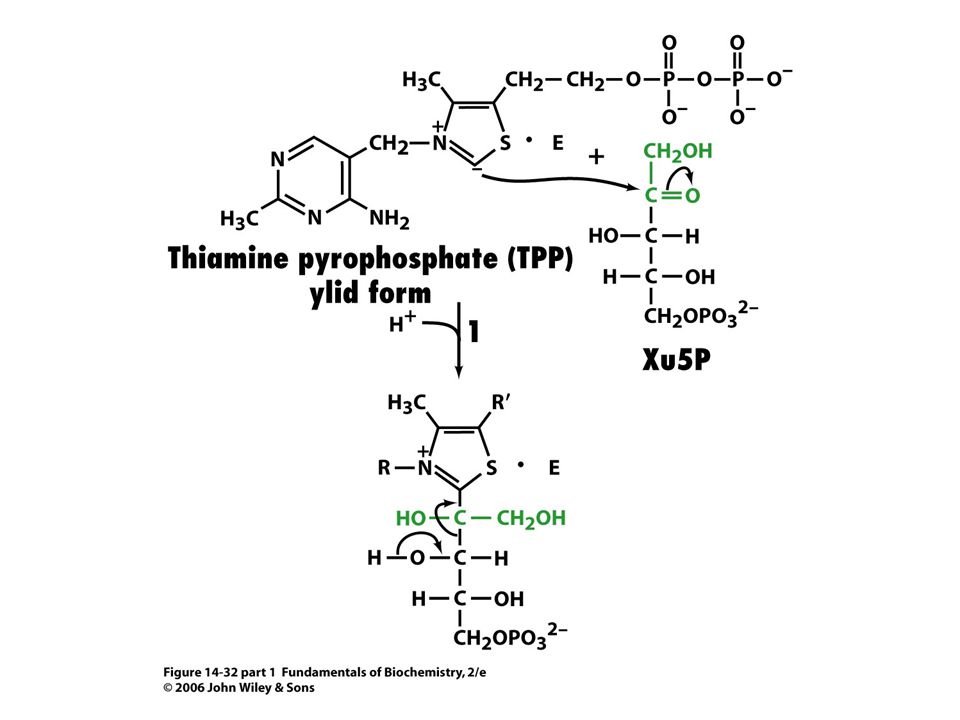
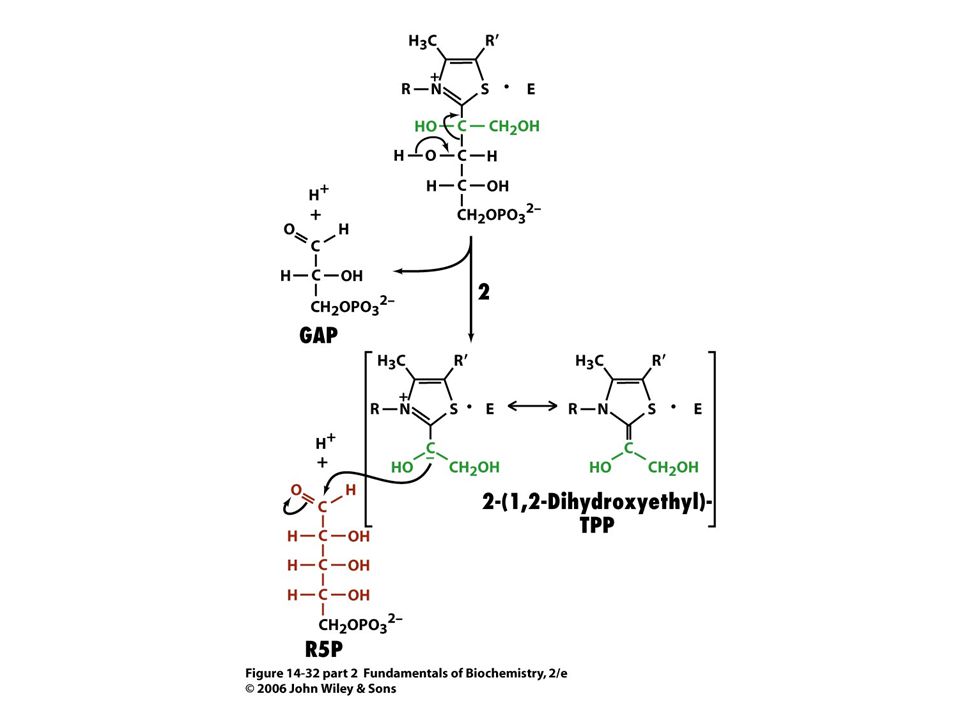
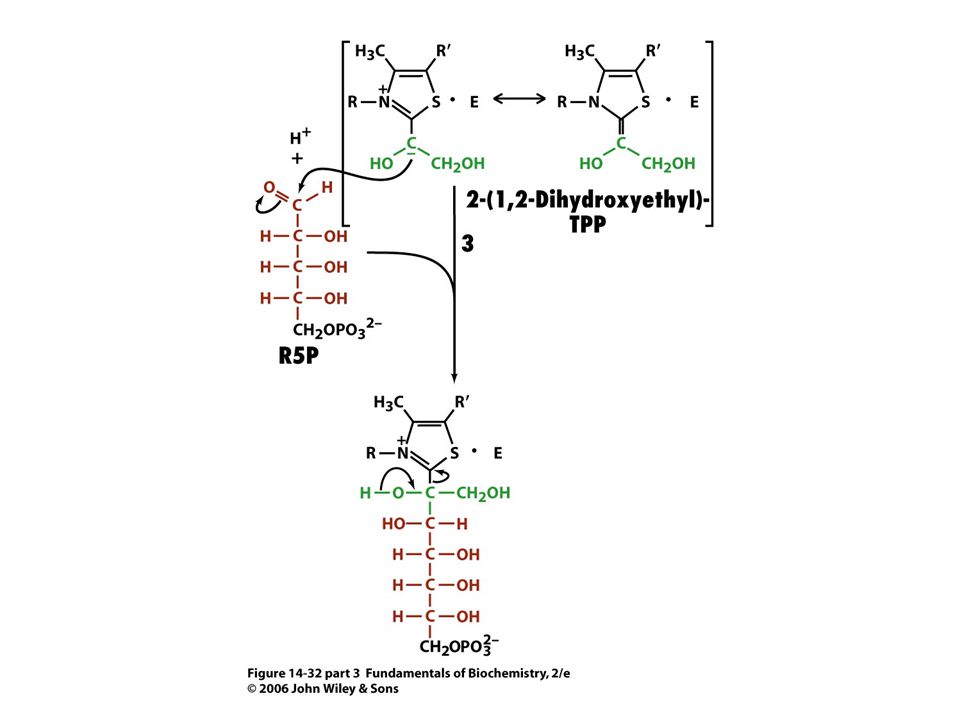
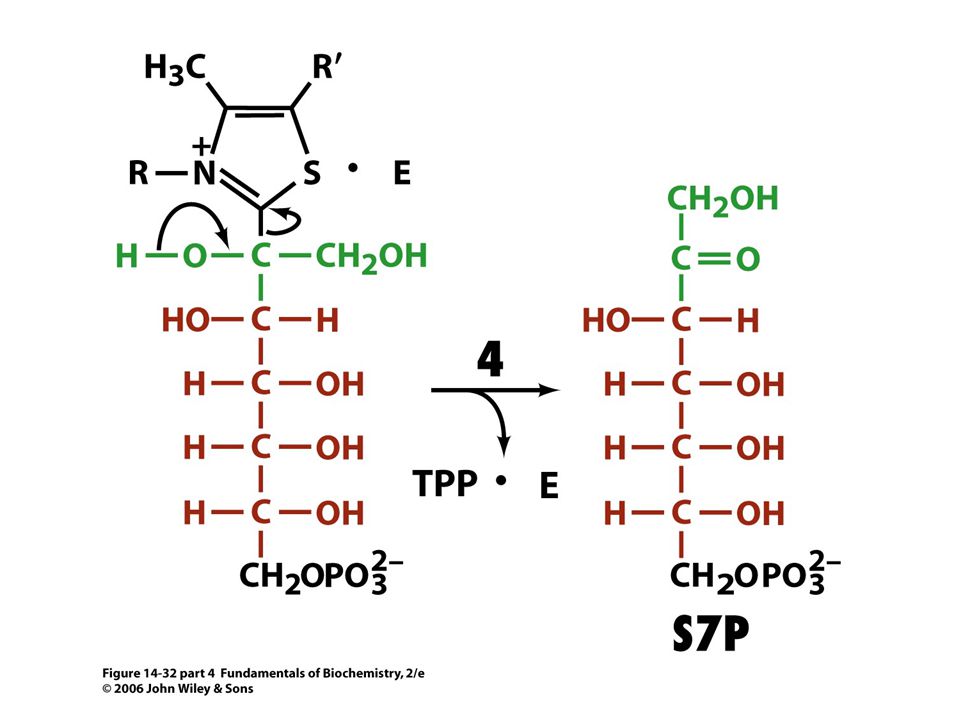
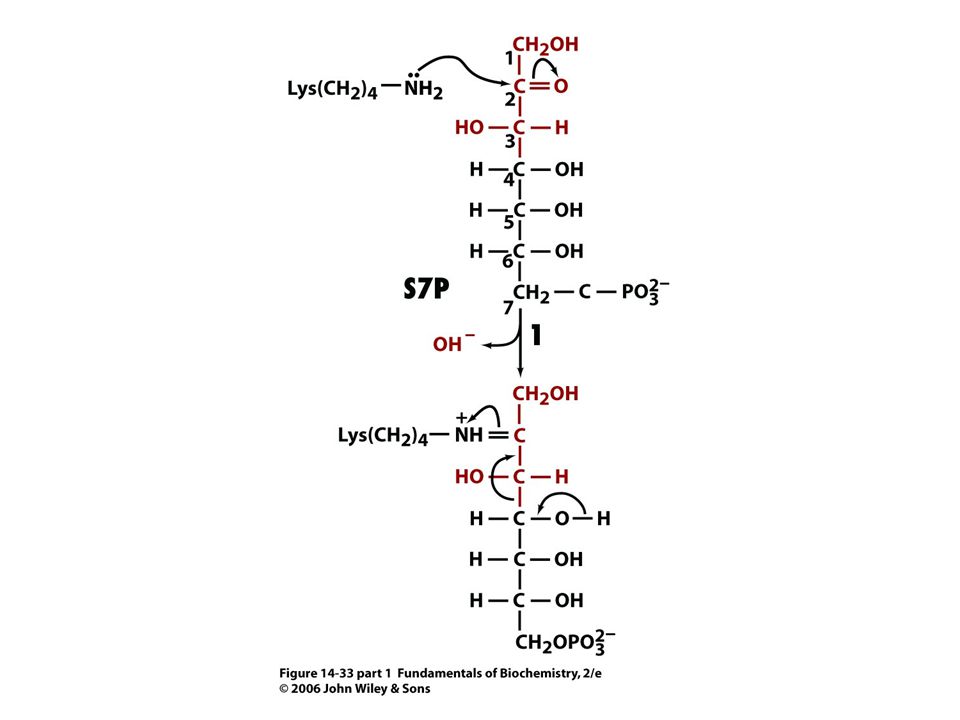
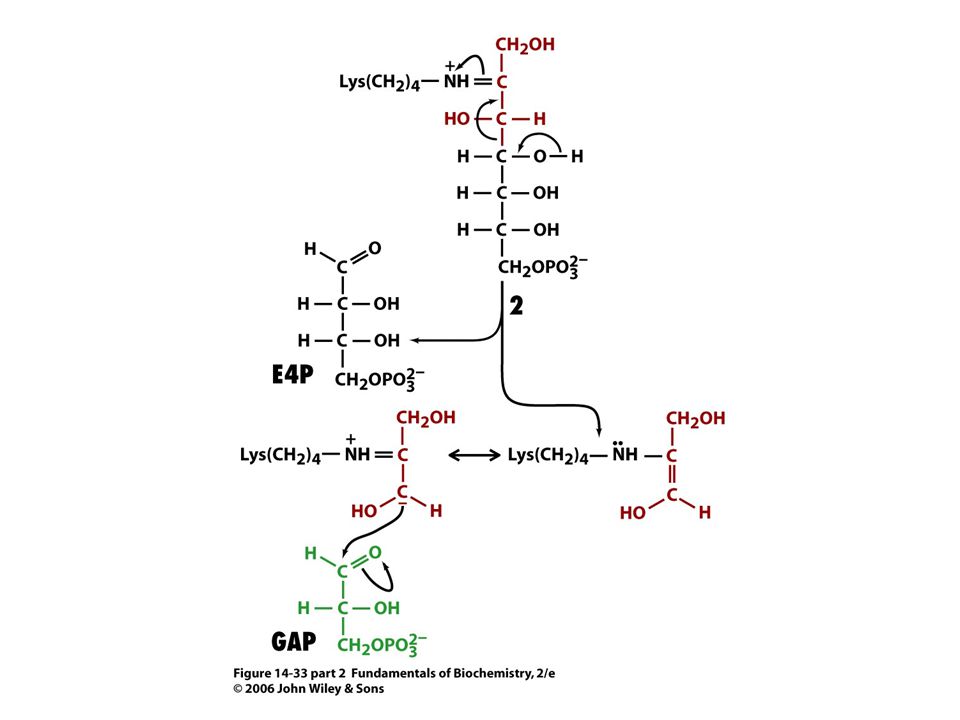
Mechanismus transketolasy

66
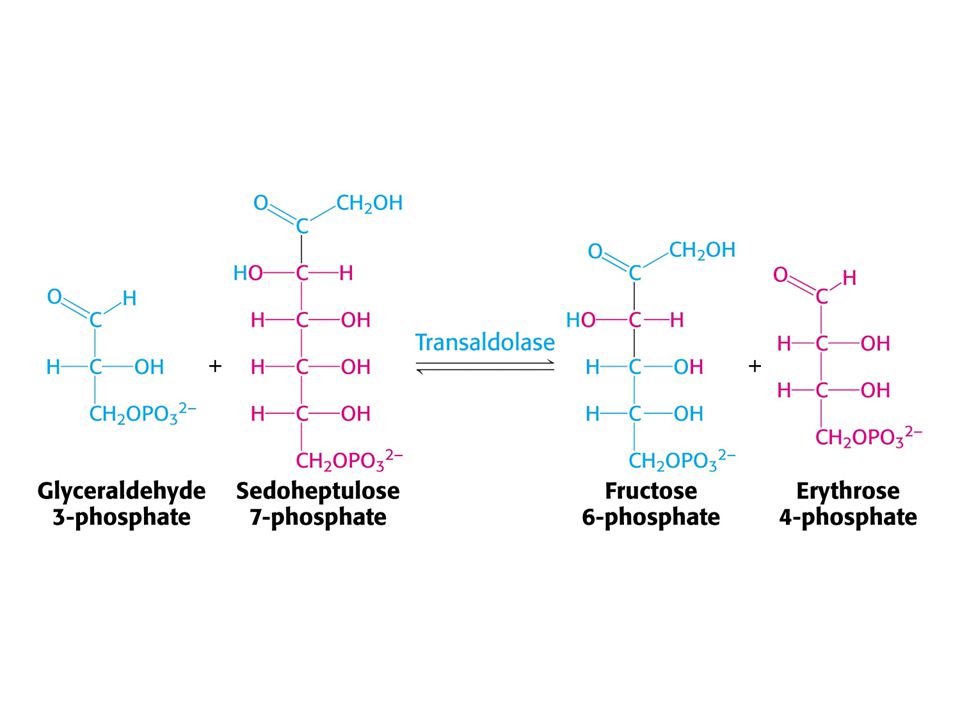
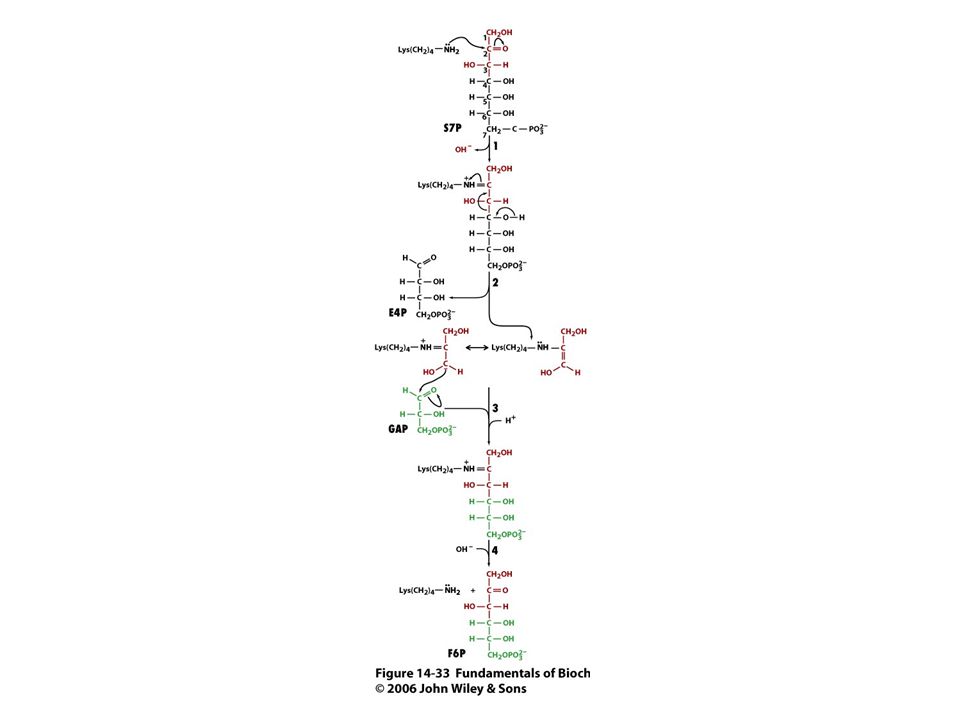
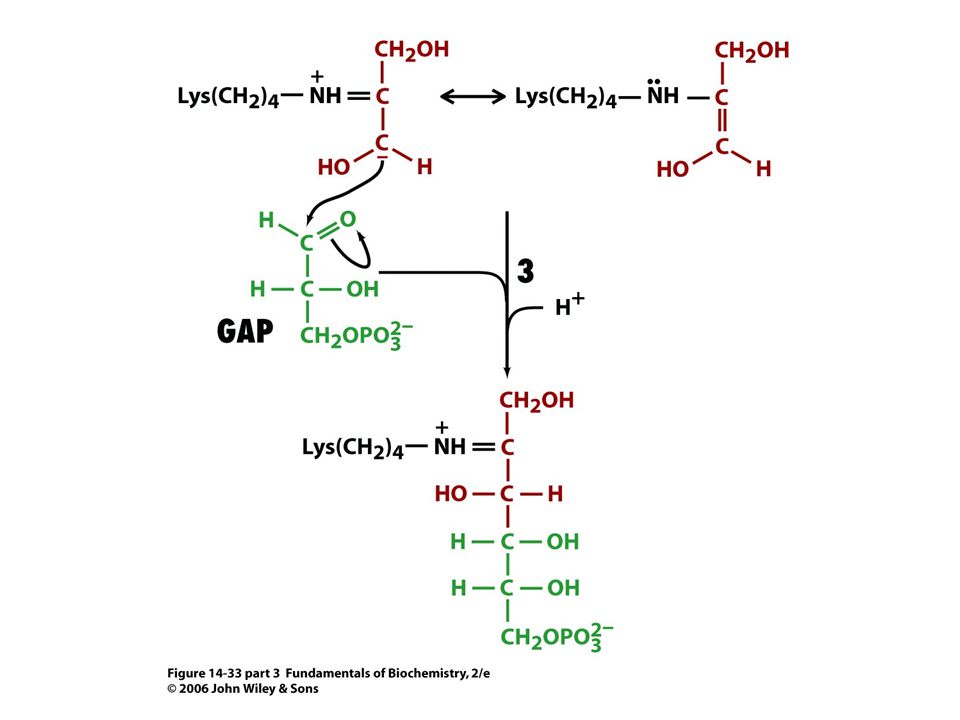
Mechanismus transaldolasy

72
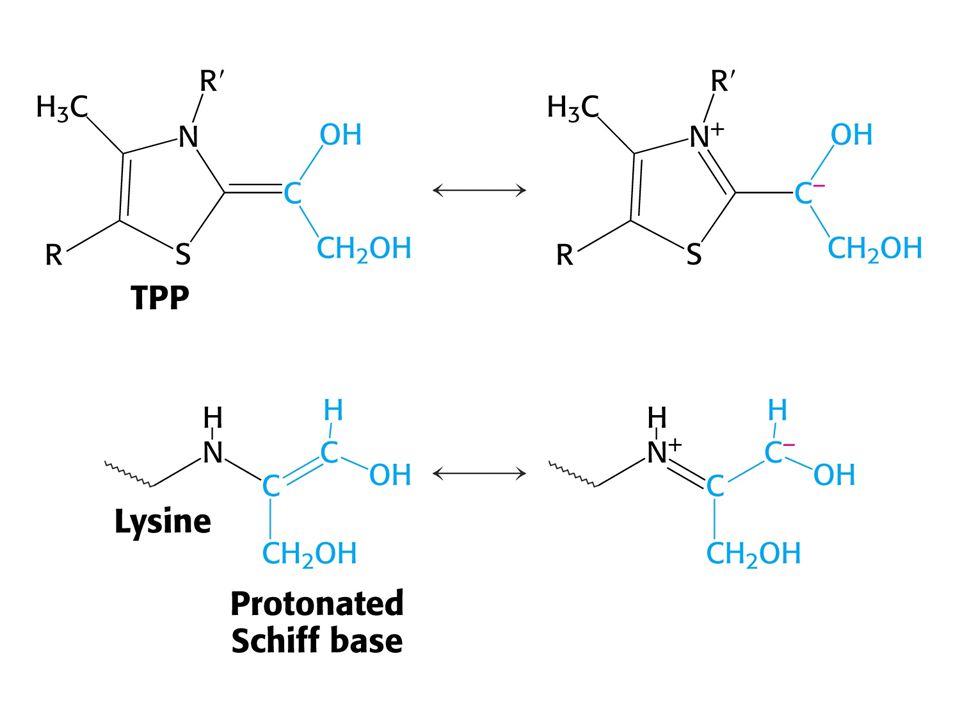
Karbanionty jako meziprodukty !
V obou případech se jedná o stabilizaci karbaniontu jako meziproduktu. V případě transketolasy je karbaniont stabilizován TPP. V případě transldolasy je karbaniont stabilizován protonovanou Schiffovou bazí.

74
Metabolismus glukosa-6-fosfátu cestou pentosafosfátů je koordinován s glykolýzou
Rychlost pentosafosfátové dráhy je kontrolována hladinou NADP+. Poměr NADP+/ NADPH v cytosolu jaterních buněk je 0, 014, zatímco poměr NAD+/NADH je za těchže podmínek 700. Hladina NADP+ je ovlivňována spotřebou NADPH pro biosyntézy.

76
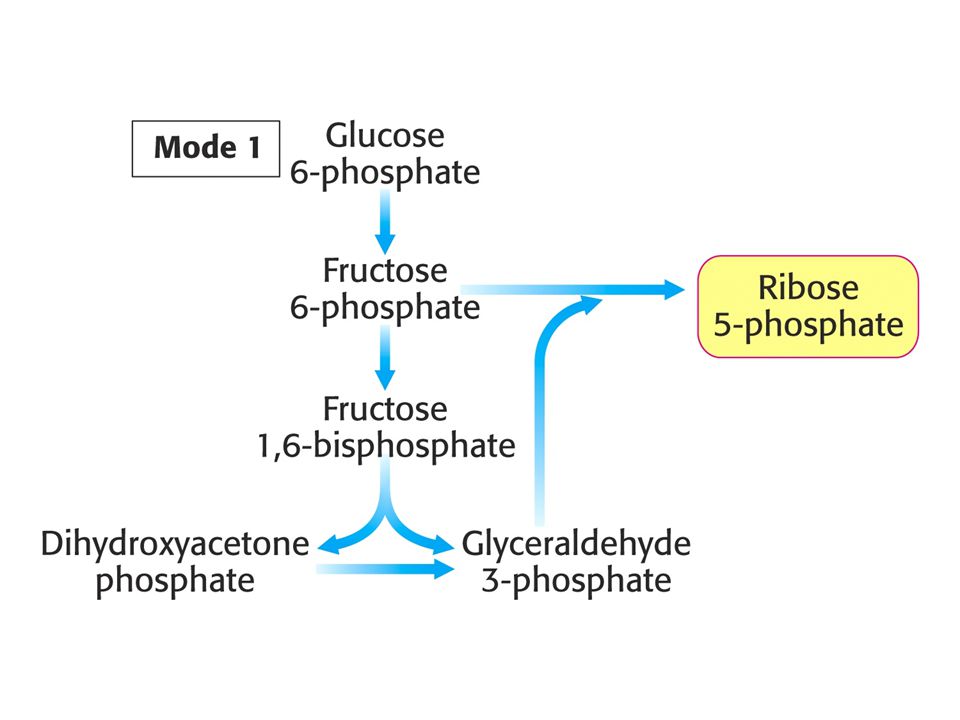
MODE 1 Potřeba ribosa-5-fosfátu je mnohem vyšší než potřeba NADPH.
Stechiometrie: 5 Glukosa-6-fosfát + ATP = 6 ribosa-5-fosfát + ADP + H+.

78
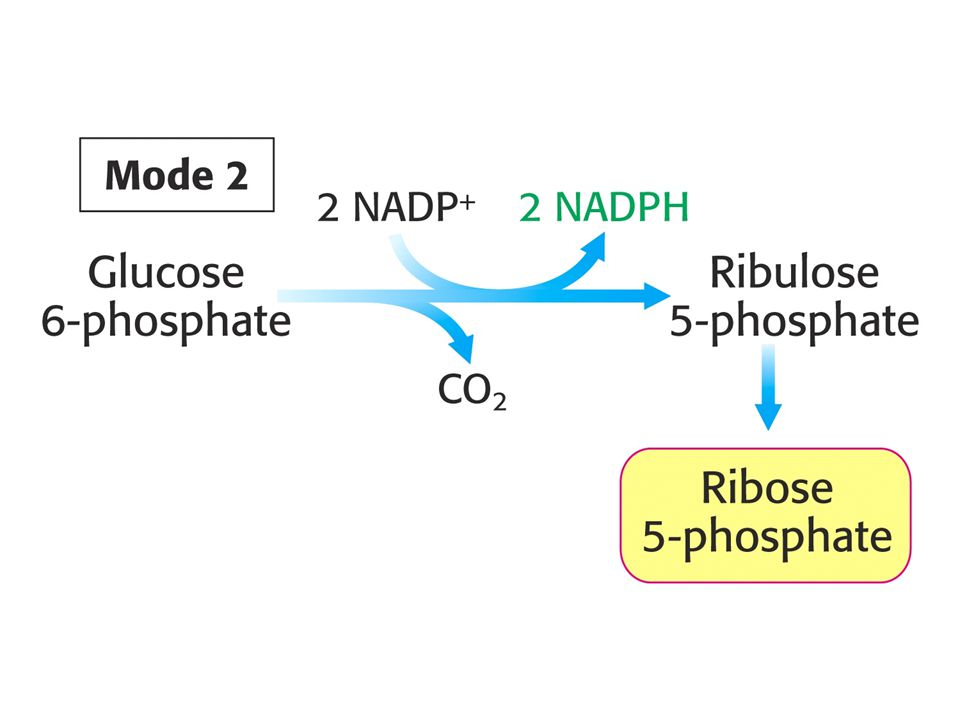
MODE 2 Potřeba NADPH a ribosa-5-fosfátu je vyrovnaná. Stechiometrie:
Glukosa-6-fosfát + 2 NADP+ + H2O = ribosa-5-fosfát + 2 NADPH + 2 H+ + CO2

80
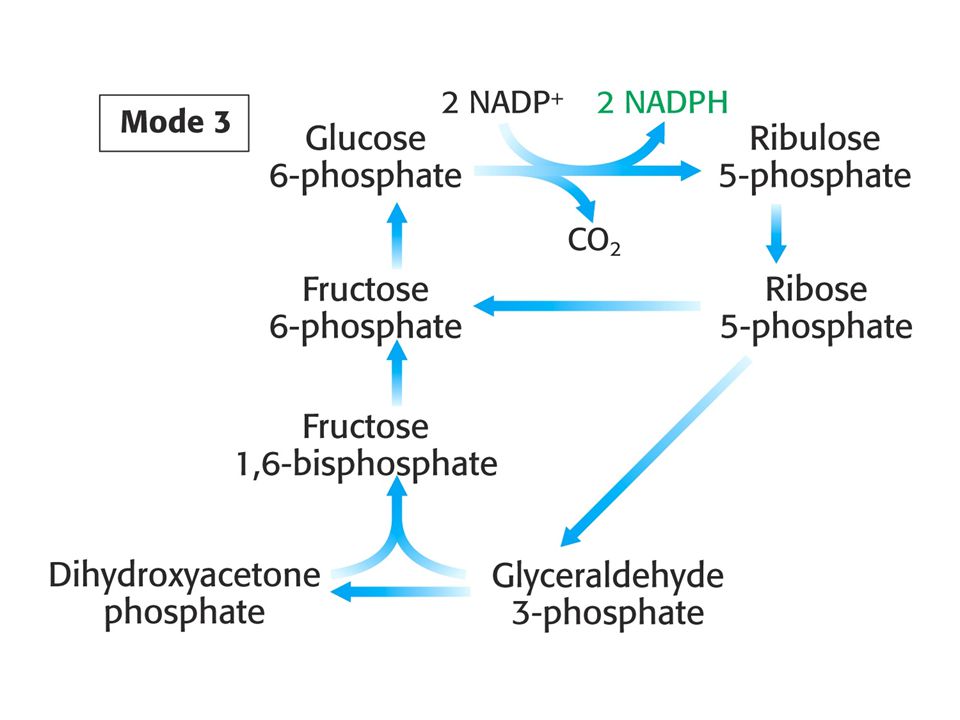
MODE 3 Je potřeba mnohem více NADPH než ribosa-5-fosfátu.
Stechiometrie tří dílčích článků: A) 6 Glukosa-6-fosfát + 12 NADP+ + 6 H2O = 6 ribosa-5-fosfát + 12 NADPH + 12 H+ + 6 CO2 B) 6 ribosa-5-fosfát = 4 fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát. C) 4 Fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát + H2O = 5 glukosa-6-fosfát + Pi Suma reakcí: Glukosa-6-fosfát + 12 NADP+ + 7H2O = 6 CO2 + + 12 NADPH + 12 H+ + Pi
 6 Glukosa-6-fosfát + 12 NADP+ + 6 H2O. = 6 ribosa-5-fosfát + 12 NADPH + 12 H+ + 6 CO2. B) 6 ribosa-5-fosfát = 4 fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát. C) 4 Fruktosa-6-fosfát + 2 glyceraldehyd-3-fosfát + H2O = 5 glukosa-6-fosfát + Pi. Suma reakcí: Glukosa-6-fosfát + 12 NADP+ + 7H2O = 6 CO NADPH + 12 H+ + Pi.")
82
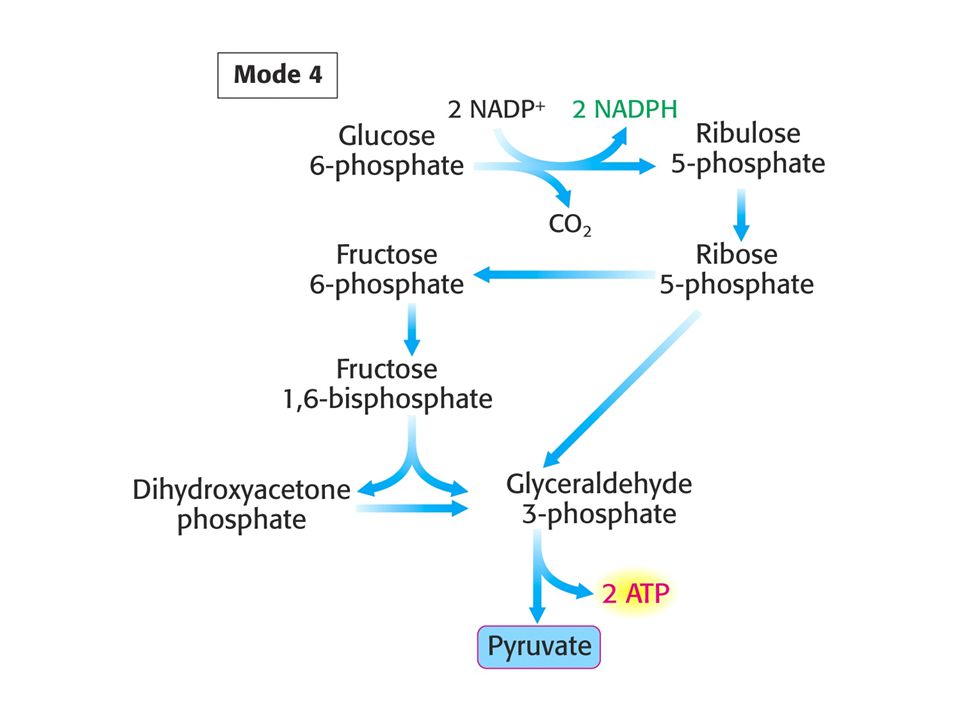
MODE 4 Je potřeba jak NADPH, tak ATP. Stechiometrie:
3 Glukosa-6-fosfát + NADP+ + 5 NAD+ + 5 Pi + 8 ADP = 5 pyruvát + 3 CO2 + 6 NADPH + 5 NADH + 8 ATP + 2 H2O + 8 H+ Pět ze šesti C atomů glukosy se objeví v pyruvátu.

84
Calvinův cyklus a pentosafosfátová dráha.
Calvinův cyklus začíná fixací CO2 a využívá NADPH k biosyntéze glukosy. Pentosafosfátová dráha začíná oxidací glukosy, odštěpuje se CO2 a tvoří se NADPH. Regenerační fáze Calvinova cyklu převádí C6 a C3 molekuly zpět na výchozí C5 molekuly (Ribulosa-1,5-bisfosfát) Pentosafosfátová dráha převádí C5 na C6 a C3 – meziprodukty glykolýzy. Příklad hospodárného a univerzálního chování živých organismů !
 Pentosafosfátová dráha převádí C5 na C6 a C3 – meziprodukty glykolýzy. Příklad hospodárného a univerzálního chování živých organismů !")
86
Glukosa-6-fosfátdehydrogenasa hraje klíčovou roli při ochraně před reaktivními kyslíkatými radikály
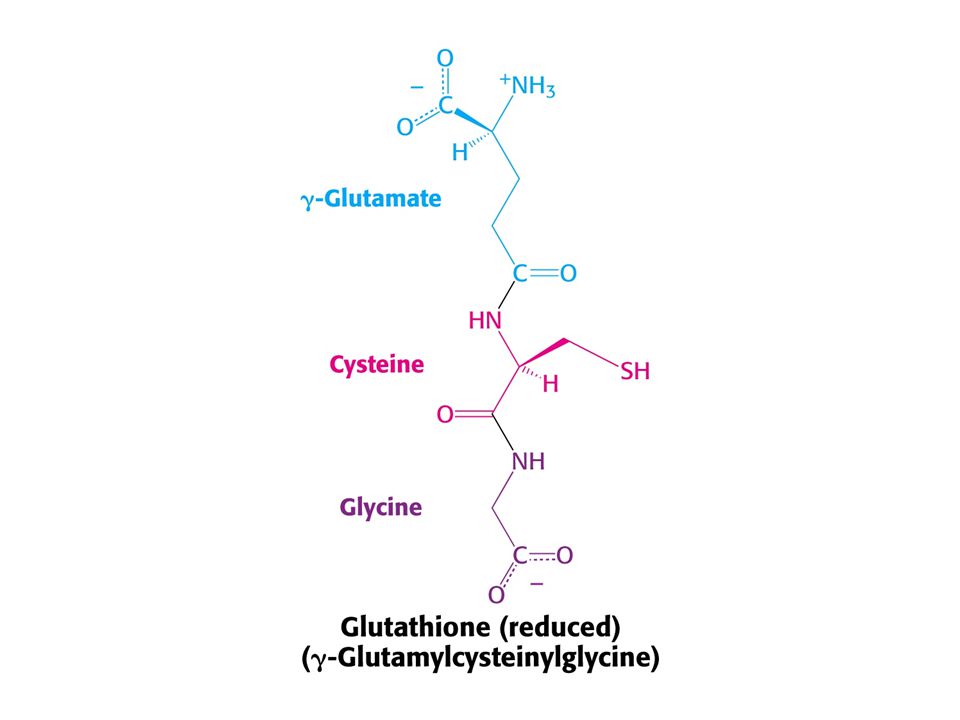
Látkou, která přímo udržuje reduktivní prostředí v buňkách je tripeptid GLUTATHION (zkratka GSH). GSH je udržován redukcí jeho oxidované formy NADPH !! Buňky s nízkou aktivitou glukosa-6-dehydrogenasy jsou citlivé na oxidativní stres. Zvláště červené krvinky, protože nemají mitochondrie, a tím i možnost alternativního zdroje redukční síly.
. GSH je udržován redukcí jeho oxidované formy NADPH !! Buňky s nízkou aktivitou glukosa-6-dehydrogenasy jsou citlivé na oxidativní stres. Zvláště červené krvinky, protože nemají mitochondrie, a tím i možnost alternativního zdroje redukční síly.")
88
Nedostatek glukosa-6-fosfátdehydrogenasy vede k hemolytické anemii.
Enzym katalyzující regeneraci GSH je glutathionreduktasa (flavoprotein). Je homologní s ferredoxin-NADP+-reduktasou (fotosyntéza). Redukovaný GSH je nutný pro normální strukturu a udržení hemoglobinu ve stavu Fe2+. Dále odstraňuje peroxidy a reaktivní kyslíkaté sloučeniny. Při nedostatku GSH dochází u hemoglobinu k jeho zesíťování, nejsou volné SH skupiny. Vytváří se agrgáty na buněčné membráně - Heinzova tělíska.
. Je homologní s ferredoxin-NADP+-reduktasou (fotosyntéza). Redukovaný GSH je nutný pro normální strukturu a udržení hemoglobinu ve stavu Fe2+. Dále odstraňuje peroxidy a reaktivní kyslíkaté sloučeniny. Při nedostatku GSH dochází u hemoglobinu k jeho zesíťování, nejsou volné SH skupiny. Vytváří se agrgáty na buněčné membráně - Heinzova tělíska.")
89
Dědičná odolnost Afričanů vůči falciparum malaria.
Parazit potřebuje ke svému množení redukovaný glutathion a produkty pentosafosfátové dráhy. Deficit glukosa-6-fosfátdehydrogenasy tak paradoxně chrání před malárií.

91
Úkoly: Calvinův cyklus a pentosafosfátový metabolismus.
1. Aldolasová reakce může probíhat opačným směrem jako enzymová aldolová kondenzace. Kolik různých produktů získáme, pokud uvažujeme, že enzym není stereospecifický ? 2. Bakteriální aldolasa netvoří Sciffovu bázi se substrátem. Obsahuje Zn2+ v aktivním místě. Jaký je mechanismus této aldolasové reakce ? 3. Arseničnan jako strukturní analog fosforečnanu může působit jako substrát řady reakcí namísto fosforečnanu. Estery arseničnanu, stejně jako estery fosforečnanu, jsou kineticky i termodynamicky nestálé a rychle hydrolyzují. Napište celkovou sumární rovnici převedení glukosy na pyruvát za přítomnosti ATP, ADP a NAD+ za účasti a) fosforečnanu, b) arseničnanu. c) Proč působí arseničnan jako jed ? 4) Porovnejte výtěžek ATP tří molekul glukosy vstupující do glykolýzy až po pyruvát s výtěžkem ATP tří molekul glukosy vstupujících do pentosafosfátového metabolismu přes tvorbu dvou fruktosa-6-fosfátů a jednoho glyceraldehyd-3-fosfátu, které jsou dále metabolizovány v glykolýze.
 fosforečnanu, b) arseničnanu. c) Proč působí arseničnan jako jed 4) Porovnejte výtěžek ATP tří molekul glukosy vstupující do glykolýzy až po pyruvát s výtěžkem ATP tří molekul glukosy vstupujících do pentosafosfátového metabolismu přes tvorbu dvou fruktosa-6-fosfátů a jednoho glyceraldehyd-3-fosfátu, které jsou dále metabolizovány v glykolýze.")
92
5. Glukosa-6-fosfát označena na uhlíku C2 prochází pentosafosfátovým metabolismem. Kde se označený uhlík objeví produktech PFM ? 6. Vysvětlete proč některé tkáně produkují oxid uhličitý za přítomnosti fluoridů, které inhibují glykolýzu. 7. Některé baktérie katabolizují glukosu tzv. Enter-Daudoroffovou drahou, což je varianta glykolýzy při které je glukosa-6-fosfát převedena na 6-fosfoglukonát (jako při pentosafosfátovém metabolismu) a poté na 2-keto-3-deoxyfosfoglukonát (KDPG). Na KDPG působí aldolasa. a) Napište struktury produktů aldolasové reakce. b) Napište jak jsou tyto reakční produkty dále metabolizovány glykolytickými enzymy. c) Jaký je výtěžek ATP v Enter-Daudoroffově dráze po stupeň pyruvátu ? Porovnejte s výtěžkem ATP při glykolýze.
 a poté na 2-keto-3-deoxyfosfoglukonát (KDPG). Na KDPG působí aldolasa. a) Napište struktury produktů aldolasové reakce. b) Napište jak jsou tyto reakční produkty dále metabolizovány glykolytickými enzymy. c) Jaký je výtěžek ATP v Enter-Daudoroffově dráze po stupeň pyruvátu Porovnejte s výtěžkem ATP při glykolýze.")
93
Řešení úkolů Calvinův cyklus a pentosafosfátový metabolismus:
1. Uhlíky C1 u glyceraldehyd-3-fosfátu a dihydroxyacetonfosfátu jsou achirální. Po kondenzaci se stanou chirálními v polohách 3 a 4 fruktosa-1,6-bisfosfátu. Vytvoří se tak 4 stereoizomery: fruktosa-1,6-bisfosfát, psikosa-1,6-bisfosfát, tagatosa-1,6-bisfosfát a sorbosa-1,6-bisfosfát. 2. Zinečnatý iont polarizuje karbonylový kyslík substrátu a stabilizuje enolátový meziprodukt reakce.

94
3. a) Glukosa + 2 NAD+ + 2 ADP + 2 Pi = 2 pyruváty + 2 NADH + 2 ATP + 2 H2O b) Glukosa + 2 NAD + 2 ADP + 2 AsO43- = 2 pyruvát + 2 NADH + 2 ADP-AsO H2O 2 ADP-AsO H2O = 2 ADP + AsO43- Celkově: Gukosa + 2 NAD = 2 pyruvát + 2 NADH c) Arseničnan je jed, protože odpojuje glykolýzu od tvorby ATP. Při glykolýze se neprodukuje energie. 4. Tři molekuly glukosy procházející glykolýzou poskytují 6 ATP. Tři molekuly glukosy procházející pentosafosfátovým metabolismem a následně glykolýzou poskytují 5 ATP. 5. Označený uhlík se objeví v polohách C1 a C3 fruktosa-6-fosfátu. 6. I když je metabolická cesta glukosy přes glykolýzu a následně citrátový cyklus blokována, glukosa může být oxidována pentosafosfátovou drahou za tvorby oxidu uhličitého. 7. a) Pyruvát + GAP b) Pyruvát jako produkt aldolasové reakce není dále metabolizován. GAP je převeden na pyruvát glykolytickými enzymy. c) Spotřebuje se jeden ATP pro převedení glukosy na glukosa-6-fosfát. Při fosfoglycerátkinasové reakci se vytvoří jeden ATP a druhý při pyruvátkinasové reakci. Čistým výtěžkem je jeden ATP na molekulu glukosy. Standardní glykolýza poskytuje 2 ATP.
 Glukosa + 2 NAD + 2 ADP + 2 AsO43- = 2 pyruvát + 2 NADH + 2 ADP-AsO H2O. 2 ADP-AsO H2O = 2 ADP + AsO43- Celkově: Gukosa + 2 NAD = 2 pyruvát + 2 NADH. c) Arseničnan je jed, protože odpojuje glykolýzu od tvorby ATP. Při glykolýze se neprodukuje energie. 4. Tři molekuly glukosy procházející glykolýzou poskytují 6 ATP. Tři molekuly glukosy procházející pentosafosfátovým metabolismem a následně glykolýzou poskytují 5 ATP. 5. Označený uhlík se objeví v polohách C1 a C3 fruktosa-6-fosfátu. 6. I když je metabolická cesta glukosy přes glykolýzu a následně citrátový cyklus blokována, glukosa může být oxidována pentosafosfátovou drahou za tvorby oxidu uhličitého. 7. a) Pyruvát + GAP. b) Pyruvát jako produkt aldolasové reakce není dále metabolizován. GAP je převeden na pyruvát glykolytickými enzymy. c) Spotřebuje se jeden ATP pro převedení glukosy na glukosa-6-fosfát. Při fosfoglycerátkinasové reakci se vytvoří jeden ATP a druhý při pyruvátkinasové reakci. Čistým výtěžkem je jeden ATP na molekulu glukosy. Standardní glykolýza poskytuje 2 ATP.")
Podobné prezentace










>")
















>")