Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Julius Lukeš , Parazitologický ústav , 2. patro , č. 225 jula@paru .
cas cz

2
Nukleové kyseliny Deoxyribonukleová kyselina (DNA) obsahuje informaci přeložitelnou do aminokyselinové sekvence proteinů Tato informace je uspořádána ve formě jednotek označovaných jako geny Ribonukleová kyselina (RNA) je součástí buněčných mechanismů, které vybírají a spojují aminokyseliny do správné sekvence Centrální dogma zní: DNA RNA Protein DNA and RNA jsou polymery nukleotidových jednotek
 je součástí buněčných mechanismů, které vybírají a spojují aminokyseliny do správné sekvence. Centrální dogma zní: DNA RNA Protein. DNA and RNA jsou polymery nukleotidových jednotek.")
3
Ústřední dogma molekulární biologie
„Svět RNA“ Ústřední dogma molekulární biologie

4
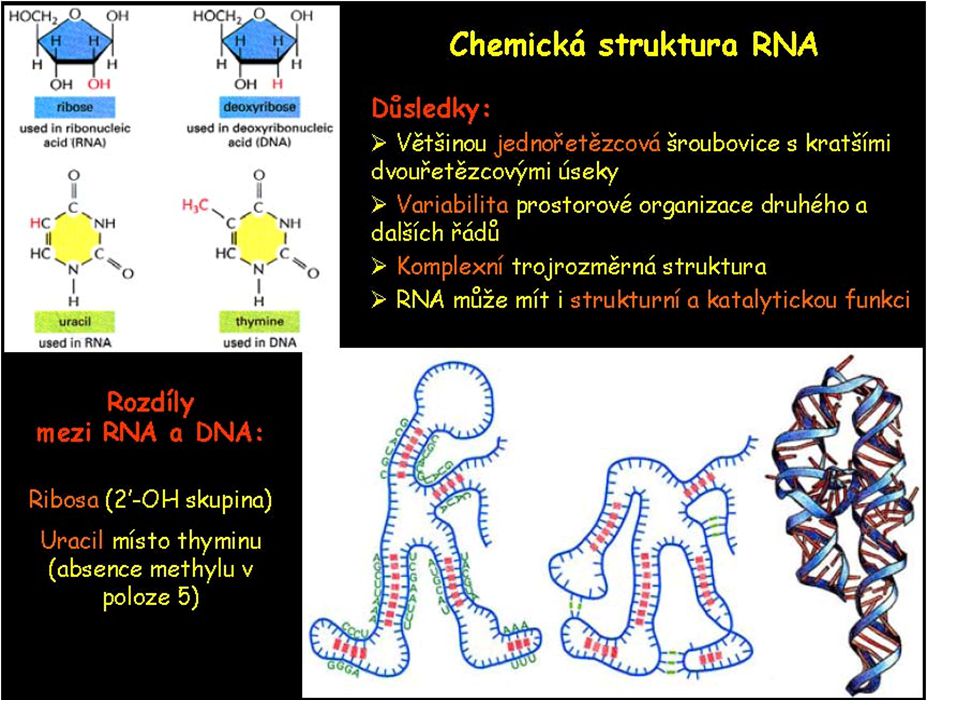
Existuje 5 hlavních bazí nukleových kyselin
A, G, T, C jsou přítomny v DNA A, G, U, C jsou přítomny v RNA

5
Všechny nukleotidy mají společnou strukturu

6
Nukleotidové podjednotky jsou vzájemně spojené fosfodiesterovou vazbou
- DNA nukleotid (nt) = fosfat + deoxyribosa (cukr) + base (A,G,C,T) Fig. 30.1
 = fosfat + deoxyribosa (cukr) + base (A,G,C,T) Fig")
8
DNA může procházet vratným oddělením vláken

9
Řada DNA molekul existuje v kruhové formě a její lokální rozvijení může způsobit superspiralizaci
supercoiled relaxed

10
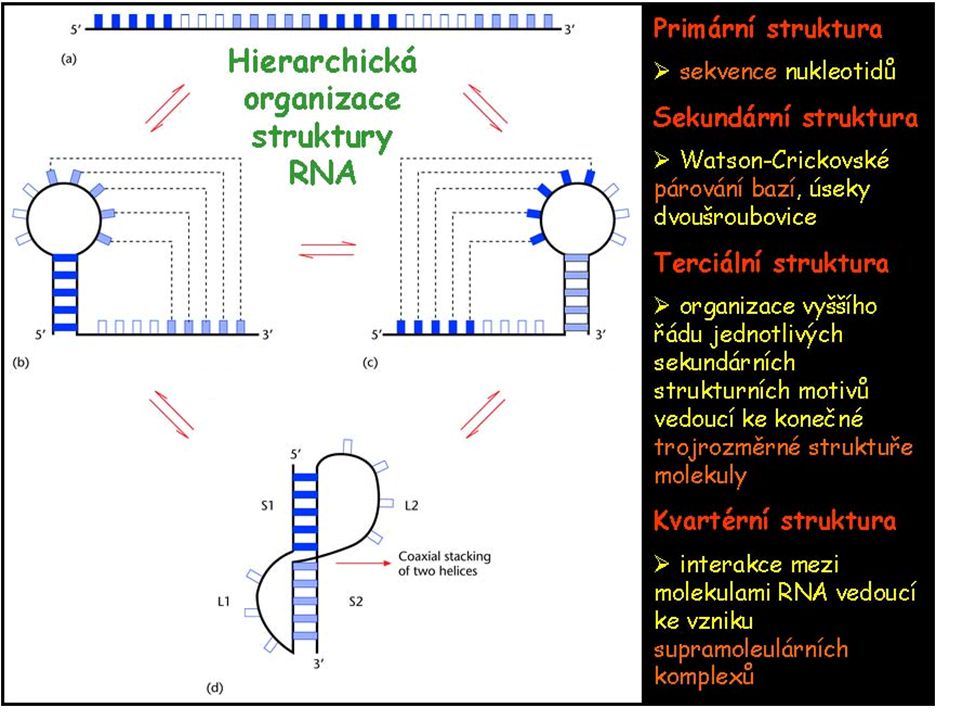
RNA molekuly se vyskytují v řadě konformací

13
Pro syntézu proteinů a nukleových kyselin platí řada společných pravidel
Proteiny a nukleové kyseliny jsou tvořené omezeným počtem různých monomerních stavebních kamenů Monomery jsou přidávány jeden po druhém Každý polypeptid a řetězec nukleové kyseliny má specifický počáteční bod, z něhož pokračuje růst jedním směrem k přesně stanovenému konci Počáteční produkt syntézy bývá často modifikován

14
Syntéza nukleových kyselin
Vlákna DNA i RNA jsou vytvářena kopírováním templátového vlákna DNA Vlákno nukleové kyseliny roste ve směru 5’ 3’ RNA polymerázy mohou samy začít syntézu vlákna, zatímco DNA polymerázy vyžadují primer(ové vlákno) Sekven(c)ování a) Syntéza DNA se řídí templátem a vyžaduje primer b) DNA polymeráza postupně přidává deoxynukleotidy ke 3' konci [5'-->3']
 Sekven(c)ování. a) Syntéza DNA se řídí templátem a vyžaduje primer. b) DNA polymeráza postupně přidává deoxynukleotidy ke 3 konci [5 -->3 ]")
15
Primer se váže do specifické oblasti
DNA polymeráza prodlužuje řetězec od 3’ konce primeru 5’ 5’ 3’

16
Sekvenování (pokrač.) c) dideoxynukleotidy - inhibitory ukončující řetězec d) enzymatické sekvenování: i) používá se směs dATP, dTTP, dGTP, dCTP a ddATP -> část molekul je ukončena na každém A [v případě ostatních tří nukleotidů se přidává ddTT nebo ddCTP nebo ddGTP] ii) produkty sekvenace jsou denaturovány a rozděleny gelovou elektroforézou iii) velikost DNA řetězců odráží pořadí bází
 používá se směs dATP, dTTP, dGTP, dCTP a ddATP -> část molekul je ukončena na každém A [v případě ostatních tří nukleotidů se přidává ddTT nebo ddCTP nebo ddGTP] ii) produkty sekvenace jsou denaturovány a rozděleny gelovou elektroforézou. iii) velikost DNA řetězců odráží pořadí bází.")
17
U dNTP může syntéza na 3’ konci pokračovat, zatímco u ddNTP to
dideoxyribonukleotidy ukončují syntézu DNA U dNTP může syntéza na 3’ konci pokračovat, zatímco u ddNTP to není možné, protože chybí 3’- OH skupina H di

18
5’ ddA A 5’ 3’ 5’ 5’ 3’ 5’ 5’ 3’ 5’ 5’ 3’

19
ATGCTTGCCAATT ddATP ddTTP ddGTP ddCTP
V souboru nasyntetizovaných molekul je v každé pozici vždy část z nich ukončena

21
Fig

22
Sekvence z automatického sekvenátoru

23
Illumina sequencing (Sequence method is not optimized for kDNA...)
23

24
sequencing by synthesis but each base contains fluorophore and terminator
24

25
fluorophore removed and 3’ block removed
25

26
Replikace DNA Replikace DNA je semikonzervativní, tzn. že každá nová molekula obsahuje staré vlákno, které slouží jako templát pro syntézu nového vlákna Je nezbytná asistence řady enzymů Dochází k málo chybám (~1/miliardu nukleotidů)
")
27
Proteiny se vyvijejí odlišnou rychlostí Fibrinopeptid 0.7 Hemoglobin 5
Cytochrom c Histon H

28
Rozdíl mezi semi-konzervativní a hypotetickou konzervativní replikací DNA

29
Hypotézy o replikaci DNA
1.při buněčném dělení dědí dceřinné buňky kopii genomu 2.konservativní replikace = 1 buňka dostane veškerou mateřskou DNA 3.dispersní replikace = nová a stará DNA jsou namíchány 4.semikonservativní replikace = každá buňka získá jedno nové a jedno staré vlákno Experiment Meselsona a Stahla (1953) 1. densitní centrifugace i) při vysokých centrifugačních silách bude roztok CsCl koncentrovanější směrem ke dnu zkumavky (=vyšší densita) ii) molekuly budou sedimentovat do oblastí odpovídajících jejich densitě iii) DNA vlákna s inkorporovaným 15N budou sedimentovat níže než STEJNÁ vlákna nesoucí 14N
 1. densitní centrifugace. i) při vysokých centrifugačních silách bude roztok CsCl koncentrovanější směrem ke dnu zkumavky (=vyšší densita) ii) molekuly budou sedimentovat do oblastí odpovídajících jejich densitě. iii) DNA vlákna s inkorporovaným 15N budou sedimentovat níže než STEJNÁ vlákna nesoucí 14N.")
30
Densitní značení DNA

31
Densitní centrifugace DNA
UV

32
2. Po kultivaci bakterií v médiu s 15N = těžká DNA byly přesunuty do média s 14N, aby viděli jak se změní densita DNA po replikaci 3. Jedna replikace -> střední densita DNA - tento fakt eliminuje konservativní replikaci

33
4. Replikace podruhé - intermediátní a lehká densita DNA- tento fakt eliminuje dispersivní replikaci

34
Replikace DNA začína ve specifických chromosomálních místech zvaných “replication origins”
Konsensus sekvence minimálního bakteriálního replikačního počátku (origin) Počátek replikace je v každém organizmu představován (1) jedinečným úsekem DNA s četnými krátkými “repeats”, (2) je rozeznáván multimerními “origin-binding” proteiny a (3) obvykle je A-T bohatý
 Počátek replikace je v každém organizmu představován (1) jedinečným úsekem DNA s četnými krátkými repeats , (2) je rozeznáván multimerními origin-binding proteiny a (3) obvykle je A-T bohatý.")
35
Replikace DNA Replikační origin: syntéza z něho běží oběma směry, čímž se tvoří bublina od replikační vidlice “replication fork” (= místo elongace DNA) (and DNA helicase)
 (and DNA helicase)")
36
Existuje řada “problémů”, které musí DNA polymeráza překonat, aby mohla kopírovat DNA
DNA polymerázy nedovedou rozvolnit DNA duplex, tak aby oddělily obě vlákna, která mají být kopírována Všechny známé DNA polymerázy dovedou pouze prodlužovat již existující vlákno DNA či RNA (tzv. “primer”) a nedovedou tudíž začít syntézu vlákna Obě vlákna v DNA duplexu mají opačnou chemickou polaritu, ale všechny DNA polymerázy katalyzují přidávání nukleotidů k 3-hydroxylovému konci rostoucího vlákna, které tudíž může růst pouze ve směru 5 k 3
 a nedovedou tudíž začít syntézu vlákna. Obě vlákna v DNA duplexu mají opačnou chemickou polaritu, ale všechny DNA polymerázy katalyzují přidávání nukleotidů k 3-hydroxylovému konci rostoucího vlákna, které tudíž může růst pouze ve směru 5 k 3")
37
K replikaci je nezbytný primer
DNA polymeráza nedovede začít sama replikovat DNA a vyžaduje tudíž, aby původní DNA obsahovala primer Primáza syntetizuje počáteční nukleotidy (RNA primer), na něž se DNA polymeráza váže RNA primer je později nahrazen nukleotidy DNA
, na něž se DNA polymeráza váže. RNA primer je později nahrazen nukleotidy DNA.")
38
Protein DnaA začíná replikaci u Escherichia coli

39
DnaB je helikáza E. coli, která dovede rozpustit duplex DNA

40
V replikační vidlici je jedno vlákno syntetizováno diskontinuálně z četných primerů

41
Vlákna DNA jsou antiparalelní
Nová DNA roste z 5’3’, jelikož DNA polymeráza připojuje nukleotidy pouze k 3’ konci vlákna DNA (leading strand) Okazakiho fragmenty, což jsou krátké úseky nově syntetizované DNA (dlouhé tisíce bází u prokaryot a stovky u eukaryot), jsou spojovány DNA ligázou za vzniku “lagging vlákna” DNA
 Okazakiho fragmenty, což jsou krátké úseky nově syntetizované DNA (dlouhé tisíce bází u prokaryot a stovky u eukaryot), jsou spojovány DNA ligázou za vzniku lagging vlákna DNA.")
42
3. Nový fragment lagging strandu je syntetizován
Syntéza leading a lagging vláken - pokračování 5’ 3’ lagging strand RNA primers leading strand 5’ 3’ 1. DNA je postupně rozvinována 3’ lagging strand 5’ leading strand RNA primers 3’ 2. leading strand je prodlužován 5’ 3. Nový fragment lagging strandu je syntetizován

43
Replikace DNA je postupná
3’ 5’ Poznámka: v tomto obrázku nejsou znázorněny RNA primery

44
Detail ligování fragmentů “lagging” vlákna, po němž se vytváří souvislé vlákno

45
Replikační vidlice

46
“Leading” a “lagging” vlákna jsou synthetizována najednou

47
Koordinace replikace “leading” & “lagging” vláken
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Koordinace replikace “leading” & “lagging” vláken

48
Souhrn proteinů Vedle primázy, DNA polymerázy a DNA ligázy jsou nezbytné 2 další proteiny Helikáza: zodpovědná za rozplétání DNA Single-strand binding proteiny (SSB): udržují původní komplementární vlákna oddělená
: udržují původní komplementární vlákna oddělená.")
49
Copyright © The McGraw-Hill Companies, Inc
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

50
Zásadní věc - chyby! Chybné zařazení nepárujících nukleotidů během syntézy je poměrně běžné a je opravováno samotnou DNA polymerázou mechanismem zvaným “mismatch repair” Mechanismem zvaným “Excision repair” se opravuje poškozená DNA post-replikačně (nesouvisí to s replikací)
")
51
Deaminace a depurinace
relativně častá deaminace může být důvodem proč není v DNA uracil

52
Pyrimidinový dimer další relativně častý typ poruchy

53
Mutace (změny v genetickém materiálu) mohou být:
Bodové mutace:chemické změny v pouze jediné nebo několika bázích v genu “Base-pair substitutions”: nahrazení jednoho nukleotidu jiným tiché měnící smysl (missence) nesmyslné (nonsense) “Insertion/Deletion”: změny v počtu nukleotidů Posun čtecího rámce (=Frame shift)
 nesmyslné (nonsense) Insertion/Deletion : změny v počtu nukleotidů. Posun čtecího rámce (=Frame shift)")
54
Poškození a oprava DNA a jejich funkce při karcinogenesi
Sekvence DNA může být změněna chybami při kopírování způsobenými DNA polymerázou během replikace, nebo látkami z vnějšího prostředí, jakými jsou např. chemické mutageny či radiace Pokud nejsou (rychle) opraveny, mohou tyto změny narušovat přirozenou funkci buňky Porucha DNA může být opravena různými procesy Karcinogeny způsobují změny v sekvenci DNA a poruchy a opravy DNA jsou tudíž důležitými aspekty při vzniku rakoviny Prokaryotické a eukaryotické opravné systémy jsou analogické
 opraveny, mohou tyto změny narušovat přirozenou funkci buňky. Porucha DNA může být opravena různými procesy. Karcinogeny způsobují změny v sekvenci DNA a poruchy a opravy DNA jsou tudíž důležitými aspekty při vzniku rakoviny. Prokaryotické a eukaryotické opravné systémy jsou analogické.")
55
Schematický model “proofreading” funkce DNA polymerázy

56
Funkce topoisomeráz při replikaci DNA
Molekuly DNA se v prostoru stáčí a ohýbají, což způsobuje změny topologie, např. “superkoily” Topoisomerázy jsou enzymy, které kontrolují topologii DNA a mají zásadní funkce během řady různých replikačních kroků V E. coli odstraňuje topo I negativní superspiralizaci DNA Topoisomerase II vnáší negativní superspiralizaci do DNA Stupeň superspiralizace DNA je určován protichůdnými aktivitami obou enzymů. Negativní superkoily jsou vnášené prostřednictvím topo II a jsou relaxované topo I

57
Rozdělení DNA topoisomerů SV40 obsahujících různý počet superkoilů

58
Topoisomerázy typu II mění topologii DNA tím, že přeštípnou a opět spojí dvouvlákno DNA

59
Topoisomerázy typu I relaxují DNA tím, že štípnou (“nicking”) a poté opětovně spojí jedno vlákno v duplexu DNA
 a poté opětovně spojí jedno vlákno v duplexu DNA")
60
Model katalytické aktivity topoisomerase II E. coli (DNA gyráza)
")
61
Replikované kruhové molekuly DNA jsou odděleny topoisomerázou typu II
Lineární dceřinné chromatidy jsou rovněž odděleny topoisomerázou II

62
Funkcí DNA gyrázy je odstraňovat pozitivní superkoily, které se tvoří před rostoucí replikační vidlicí

63
Je celá řada proteinů, které se účastní biogeneze DNA a RNA v buňce - DNázy, RNázy (endonukleázy a exonukleázy), restrikční enzymy (štípající s převisem, na tupo; 4katry až n-katry) atd. Budete probírat podrobněji v metodických přednáškách, protože využití vlastností těchto a jiných proteinů stojí de facto za obrovským pokrokem molekulární biologie

64
Problém: jak uložit tolik DNA do genomu?

65
DNA-replikace je rozsáhlá záležitost
DNA molekuly jsou enormně dlouhé, u člověka se jejich velikost pohybuje od do nt human chromosomes

66
Periodicita nukleozomu

67
Organizace nukleozomu
1.8 závitů (ca. 140 bp) DNA ovinutých kolem 8 histonů = [H2A + H2B]2 + [H3 + H4]2 Histon H1 (nepatří mezi “core” histony) se na DNA váže v místě vstupu a výstupu DNA z nukleozomu
 DNA ovinutých kolem 8 histonů = [H2A + H2B]2 + [H3 + H4]2. Histon H1 (nepatří mezi core histony) se na DNA váže v místě vstupu a výstupu DNA z nukleozomu.")
68
Struktura nukleosomu [H3 + H4]2 vytváří jádro s jedním heterodimerem [H2A + H2B] nad ním a dalším [H2A + H2B] pod ním
![Struktura nukleosomu [H3 + H4]2 vytváří jádro s jedním heterodimerem [H2A + H2B] nad ním a dalším [H2A + H2B] pod ním.](http://slideplayer.cz/slide/2991743/11/images/68/Struktura+nukleosomu+%5BH3+%2B+H4%5D2+vytv%C3%A1%C5%99%C3%AD+j%C3%A1dro+s+jedn%C3%ADm+heterodimerem+%5BH2A+%2B+H2B%5D+nad+n%C3%ADm+a+dal%C5%A1%C3%ADm+%5BH2A+%2B+H2B%5D+pod+n%C3%ADm..jpg "Struktura nukleosomu [H3 + H4]2 vytváří jádro s jedním heterodimerem [H2A + H2B] nad ním a dalším [H2A + H2B] pod ním.")
69
30 nm silné vlákno Mohou být stabilizovány:
histonem H1 (váže se ve vstupu a výstupu DNA z nukleozomu) histonovými konci
 histonovými konci.")
70
30 nm silné vlákno vytváří spirálu (solenoid)
6 nukleosomů/otáčku je stabilizováno histonem H1

71
Nukleosomy jsou komplexy histonů

72
Úrovně sbalení chromatinu viděné v elektronovém mikroskopu
Výsledek: DNA je sbalena do mitotického chromosomu, který je x kratší než celková délka DNA

73
DNA je kompaktně sbalena díky vazbě proteinů.
Nejvyšší strukturou je mitotický chromosom.

74
Remodelační komplexy jsou komplexy proteinů zpracovávající
ATP,které se pohybují po DNA a rozvolňují a vytváří nukleosomy a umožňují vazbu DNA-binding proteinů, např DNA a RNA polymeráz a transkripčních faktorů

75
Existují 2 typy chromatinu
heterochromatin = kompaktní chromatin euchromatin = rozvolněný chromatin

76
Acetylace histonů aktivuje transkripci
(HAT = histone acetyltransferáza)
")
77
Modifikace proteinů mění jejich funkci
Metylace - připojení CH3 k lysinu Fosforylace - připojení PO4- Acetylace - připojení CH3CO Např. acetylací lysinu nebo fosforylací serinu se mění celkový kladný náboj proteinu

78
Copyright © The McGraw-Hill Companies, Inc
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

79
Copyright © The McGraw-Hill Companies, Inc
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

80
Molekulární definice genu
Gen je takový úsek DNA, který je nutný pro syntézu funkčního polypeptidu Oblasti DNA, které kódují molekuly RNA jako tRNA a rRNA také považujeme za geny V eukaryotické DNA leží geny mezi velkými úseky nefunkční, nekódující DNA. Geny mohou rovněž obsahovat oblasti nekódující DNA

81
Genom vyšších eukaryot obsahuje hodně “nefunkční” DNA
U eukaryot nekoreluje velikost genomu s fylogenezí

82
Obsah DNA je v eukaryotickém genomu velmi variabilní
V průběhu evoluce se genom zmenšuje nebo expanduje?

83
Bakteriální operony vytvářejí polycistronickou mRNAs, zatímco většina eukaryotických mRNAs je monocistronická a obsahuje introny

84
V eukaryotických genomech se nalézají složité i jednoduché transkripční jednotky

85
Různé typy eukaryotické DNA nejsou nikdy přepisované

86
Mobilní DNA Mobilní sekvence (částečně opakované) se vyskytují v prokaryotických genomech, u vyšších rostlin a živočichů Tyto sekvence jsou dlouhé od stovek do tisíců bází Tyto sekvence jsou kopírované a integrované (=vkládané) do nových oblastí v genomu procesem transpozice Pro mobilní sekvence není známa žádná užitečná funkce
 do nových oblastí v genomu procesem transpozice. Pro mobilní sekvence není známa žádná užitečná funkce.")
87
Mobilní elementy se mohou pohybovat ve formě DNA či RNA

88
Obecná struktura bacteriálního transposonu

89
Mobilní elementy mají patrně velký význam během evoluce
Inzerce mobilního elementu do nebo poblíž transkripční jednotky může způsobit spontánní mutace Homologní rekombinace mezi mobilním DNA elementem může přispět ke genové duplikaci a jiným přestavbám, včetně duplikace intronů, rekombinace intronů s následnou tvorbou nových genů a změnami kontroly genové exprese

90
Funkční přestavby chromosomální DNA
Zatímco se zdá, že transposice mobilního elementu nemá žádný okamžitý vliv na organismus, různé typy přestavby DNA jsou pro něj výhodné a důležité Příkladem jsou inverze a delece úseků DNA a jejich amplifikace Tyto funkční přestavby probíhají u prokaryot i eukaryot

91
Organizace buněčné DNA do chromosomů
Většinou jsou bakteriální chromosomy kruhové s pouze jedním počátkem replikace Každý eukaryotický chromosom je tvořen jednou lineární molekulou DNA, která obsahuje několik počátků replikace (=origins of replication) Eukaryotická DNA je asocióvaná s histony, čímž se vytváří chromatin Mikroskopická pozorování počtu a velikosti eukaryotických chromosomů a způsob jejich barvení odhalily řadu důležitých aspektů jejich struktury
 Eukaryotická DNA je asocióvaná s histony, čímž se vytváří chromatin. Mikroskopická pozorování počtu a velikosti eukaryotických chromosomů a způsob jejich barvení odhalily řadu důležitých aspektů jejich struktury.")
92
Barvené chromosomy mají charakteristické proužkování

93
Počet chromosomů a jejich velikost a tvar během metafáze jsou druhově specifické

94
Jednotlivé chromosomy mají v jádře přesné místo

95
Pro replikaci a dědění eukaryotického chromosomu jsou nezbytné 3 funkční elementy
počátky replikace DNA centromera 2 konce (telomery)
")
96
Centromery jsou nezbytné pro stabilitu chromosomů během mitózy
jsou strukturně složité DNA v lidských centromerách je představována rodinou vysoce repetitivní, tandemově uspořádané ‘satelitní’ DNA, která měří na délku 300-5,000 kb

97
Jak fungují centromery?
Jsou místem tvorby kinetochorů, které umožňují přichycení sesterských chromatid k mikrotubulům, vybíhajícím z pólů dělícího vřeténka kinetochor je velký proteinový komplex, který zprostředkovává vazbu mezi vřeténkem a chromosomy

98
Konce DNA molekul - telomery:
umožňují úplnou replikaci chromosomálních konců chrání chromosomy před zkracováním a spojováním s jinými molekulami DNA.

99
Telomery (pokračování)
Všechny lidské metafázové chromosomy hybridizují s telomerickou repetitivní sekvencí (TTAGG)n
n.")
100
Telomery (pokračování)
DNA polymeráza nedovede syntetizovat úplné konce DNA molekuly Postupné zkracování po každé replikaci by mělo za následek ztrátu důležité informace Telomeráza dosyntetizovává mnoho kopií sekvence TTAGGG na konce DNA molekul (=telomery) Telomeráza se obvykle nachází pouze v zárodečných a pohlavních buňkách Její přítomnost v rakovinných buňkách může vést ke vzniku nádorů
 Telomeráza se obvykle nachází pouze v zárodečných a pohlavních buňkách. Její přítomnost v rakovinných buňkách může vést ke vzniku nádorů.")
101
Jak se telomery vytvářejí?
Telomeráza je komplex proteinu s RNA, který má reverzně transkripční aktivitu

102
Telomeráza si tudíž ve formě RNA sebou nosí svůj templát

103
Telomeráza (pokrač.)
")
104
Organelární DNA Mitochondrie obsahují jednu, ale většinou více molekul mtDNA Geny mtDNA vykazují cytoplasmatickou dědičnost a kódují rRNAs, tRNAs, a několik mitochondriálních proteinů Velikost a kódovací kapacita mtDNA je u jednotlivých organismů velmi odlišná Produkty mitochondriálních genů nejsou exportovány Mutace v mtDNA způsobují u lidí závažné genetické choroby Chloroplasty obsahují velkou kruhovou DNA kodující více než 100 proteins Apikoplast a jiné organely u prvoků mají svojí vlastní DNA

105
Lidská mtDNA

106
Mitochondriální genetické kódy se odlišují od standartního genetického kódu

108
Obecný postup klonování s plasmidovými vektory

109
Klonování pomocí plasmidů umožňuje isolaci cílových molekul DNA z směsných roztoků

110
LABORATORY OF MOLECULAR BIOLOGY OF PROTISTS
Kontakt: Julius Lukeš - Nabídka bakalářské práce v naší laborce (nástup v listopadu 2006) Zapojení v těchto projektech: Functional analysis by RNA interference of accessory factors in RNA editing Functional analysis by RNA interference of Fe-S cluster biosynthesis pathway Complementational analysis of putative mitochondrial proteins in various protozoans
 Zapojení v těchto projektech: Functional analysis by RNA interference of accessory factors in RNA editing Functional analysis by RNA interference of Fe-S cluster biosynthesis pathway. Complementational analysis of putative mitochondrial proteins in various protozoans")
111
Cytoplasmatická dědičnost mutantních mitochondrií

Podobné prezentace




>")






>")

>")








