Stáhnout prezentaci
1
e-mail: janecekv@fle.czu.cz
BIOLOGIE LESA II HSSL Genetika – tato verze je bez funkčních videosekvencí PRAHA, samostudium 8.3. Vladimír Janeček KDŠLD L357 tel

2
Podklady pro studium http://genetika.wz.cz/
Paule, L., 1992: Genetika a šĺachtenie lesných drevín. Príroda, 304 s. Pospíšil, J., Kobliha, J., 1988: Šlechtění lesních dřevin. VŠZ Brno, 135 s.

3
ŠLECHTĚNÍ LESNÍCH DŘEVIN
GENETIKA věda zabývající se dědičnou proměnlivostí živých organizmů ŠLECHTĚNÍ LESNÍCH DŘEVIN vědní obor nebo technologie aplikující poznatky z genetiky na zlepšení genetické konstituce lesních dřevin prakticky se jedná o člověkem řízenou evoluci

4
STRUČNÝ VÝVOJ GENETIKY
1865 J. G. Mendel ( ): Versuche über Pflanzenhybriden (Pokusy s rostlinnými kříženci) - Mendelovy zákony a mezialelární vztahy počátek 20. stol.: Hugo de Vries ( ), Erich Tschermak ( ), Carl Correns ( ) – znovuobjevení Mendelových zákonů 1903 Sutton, 1904 Boveri: Objev spojitosti zákonitého štěpení hybridů a dělení chromozómů při meióze 1906 William Bateson ( ) - genetika, heterozygot a homozygot 1909 Wilhelm Johansen ( ) - gen, genotyp a fenotyp 1923 Morgan –získání ověřených dokladů o lokalizaci genů a jejich možné rozdílné kombinaci 1953 James D. Watson a Francis H. Crick - strukturní model dvojšroubovice DNA současnost – rozluštění genetického kódu člověka, klonování zvířat, ….
: Versuche über Pflanzenhybriden (Pokusy s rostlinnými kříženci) - Mendelovy zákony a mezialelární vztahy. počátek 20. stol.: Hugo de Vries ( ), Erich Tschermak ( ), Carl Correns ( ) – znovuobjevení Mendelových zákonů Sutton, 1904 Boveri: Objev spojitosti zákonitého štěpení hybridů a dělení chromozómů při meióze William Bateson ( ) - genetika, heterozygot a homozygot Wilhelm Johansen ( ) - gen, genotyp a fenotyp Morgan –získání ověřených dokladů o lokalizaci genů a jejich možné rozdílné kombinaci James D. Watson a Francis H. Crick - strukturní model dvojšroubovice DNA. současnost – rozluštění genetického kódu člověka, klonování zvířat, ….")
5
Buňka Buňka jádro Jádro Chromozomy Ribozómy aaaaaaaaaaaaaa
Mitochondrie aaaaaaaaaaaaaa jádroeeeeeee

6
Buněčné jádro Chromozomy Jádro Jaderné póry

7
Chromozomy Vyskytují se v sadách
Nejčastější jsou diploidní organismy s 2 sadami, vyskytují se tedy v párech Gamety (pohlavní buňky) mají pouze 1 sadu
 mají pouze 1 sadu.")
8
Dělení chromozomů submetacentrický metacentrický subtelocentrický
se satelitem

9
Dělení chromozomů Chromosomy se dělí na chromosomy somatické (autosomy, které tvoří homologní páry a určují všechny vlastnosti organismu mimo pohlaví) a chromosomy pohlavní (gonosomy), které určují pohlaví jedince (ale nesou i jiné geny) a jsou heterologní (označení X a Y).
 a chromosomy pohlavní (gonosomy), které určují pohlaví jedince (ale nesou i jiné geny) a jsou heterologní (označení X a Y).")
10
Určení pohlaví Kombinací gonozomů (X a Y). Existují různé typy určení pohlaví: a) Savčí typ (drosophila): Je nejčastější - savci, plazi, obojživelníci, většina hmyzu a dvoudomých rostlin. Samičí pohlaví XX (samičí gameta vždy jen chromozóm X). Samčí pohlaví XY (samčí gameta nese chromozom X nebo Y, šance 50:50). b) Ptačí typ (abraxas): Vyskytuje se u ptáků, motýlů a některých ryb. Samice XY, samec XX. Navíc některé řády hmyzu nemají chromozom Y, samčí pohlaví je určeno přítomností jednoho chromozomu X. U jiných druhů hmyzu zase pohlaví jedince může být určeno vnějšími znaky (včela). Gonozomálně se dědí ovšem i další dědičné znaky uložené na gonosomech, navíc i některé choroby. Často uváděným příkladem je hemofilie = chorobná nesrážlivost krve. Chorobu podmiňuje recesivní alela z chromozomu X. Možnosti jsou muž: XY-zdravý, xY-nemocný; žena XX-zdravá, Xx-přenašečka, xx-nemocná.
 Savčí typ (drosophila): Je nejčastější - savci, plazi, obojživelníci, většina hmyzu a dvoudomých rostlin. Samičí pohlaví XX (samičí gameta vždy jen chromozóm X). Samčí pohlaví XY (samčí gameta nese chromozom X nebo Y, šance 50:50). b) Ptačí typ (abraxas): Vyskytuje se u ptáků, motýlů a některých ryb. Samice XY, samec XX. Navíc některé řády hmyzu nemají chromozom Y, samčí pohlaví je určeno přítomností jednoho chromozomu X. U jiných druhů hmyzu zase pohlaví jedince může být určeno vnějšími znaky (včela). Gonozomálně se dědí ovšem i další dědičné znaky uložené na gonosomech, navíc i některé choroby. Často uváděným příkladem je hemofilie = chorobná nesrážlivost krve. Chorobu podmiňuje recesivní alela z chromozomu X. Možnosti jsou. muž: XY-zdravý, xY-nemocný; žena XX-zdravá, Xx-přenašečka, xx-nemocná.")
11
Chromozomy Chromosome Jádro Buňka Jaderné Chromozom Telomera
Chromatidy Chromosome Telomera Centromera Buňka Telomera Jaderné Histony Nukleotidové báze Nukl DNA (dvojšroubovice)
")
12
Základní počty chromozomů
Rod / Druh Betula pendula 14 Populus tremula 19 Salix Picea abies 12 Pinus silvestris Sequoia sempervirens 11 Tilia 41 Taxus

13
Karyotyp Druhově specifický stav souboru chromozómů somatické buňky co do počtu, velikosti a morfologie chromozómů Počet chromozómů nesouvisí s velikostí organizmu ani s jeho evoluční vyspělostí Příbuzné druhy mohou mít podobné, i když poněkud odlišné chromozómy. Je možné studovat evoluci karyotypů

14
Karyotyp-člověk Cytogenetic map of human chromosomes numbered according to the Paris classification of Note: Chromosome size decreases as chromosome number increases (Ex: Chr 1 is larger than Chr 6)
")
15
DNA Picture spins!

16
Struktura DNA nosná kostra DNA homopolymerní řetězec cukr-fosfát
Molekula DNA má tvar pravotočivé dvojšroubovice. Obě vlákna, skládající se z řetězce cukr (2-deoxyribóza) a fosfát a dále z připojených nukleotidových bází, jsou vzájemně komplementárně spojena vodíkovými můstky mezi párovými bázemi, a sice adenin (A) s thyminem (T) a guanin (G) s cytosinem (C). A a T jsou spojeny dvěma vodíkovými můstky, zatímco C a G třemi. Na 1 otáčku připadá cca 10 nukleotidových párů. Haploidní genom člověka obsahuje 3,000,000,000 párů bází, které jsou rozděleny mezi 22 párů autozómů a 1 pár gonozómů.
 a fosfát a dále z připojených nukleotidových bází, jsou vzájemně komplementárně spojena vodíkovými můstky mezi párovými bázemi, a sice adenin (A) s thyminem (T) a guanin (G) s cytosinem (C). A a T jsou spojeny dvěma vodíkovými můstky, zatímco C a G třemi. Na 1 otáčku připadá cca 10 nukleotidových párů. Haploidní genom člověka obsahuje 3,000,000,000 párů bází, které jsou rozděleny mezi 22 párů autozómů a 1 pár gonozómů.")
17
Páry bází DNA Na 1 otáčku připadá cca 10 nukleotidových párů
The binding of two nucleotides forms a base pair. In DNA, cytosine and guanine are bound together by 3 hydrogen bonds, whereas adenine and thymine are bound by 2 hydrogen bonds. Therefore, the link between cytosine and guanine is much stronger than the link between adenine and thymine.

18
Charakteristiky DNA Organizmus Velikost genomu a Počet genů
Kódující DNA % Escherichia coli 5 4000 100 Kvasinky 12 6000 50 Picea abies 30000 35000 <3 Člověk 3000 50000 <2 a 106 párů bází u haploidní sady (čísla orientační)
")
19
DĚDIČNOST přenos genetické informace z rodičů na potomstvo (a tedy z generace na generaci) PROMĚNLIVOST = VARIABILITA odchylnost od průměrné hodnoty znaku, od normálu absence uniformity

20
GEN základní jednotka většiny typů dědičnosti; sekvence nukleotidových bází (nebo lépe tripletů) okupující určitý lokus na chromozómu a nesoucí informaci, která po transkripci a translaci kóduje syntézu jednoho polypeptidového řetězce (+ výjimky) triplet = sekvence 3 nukleotidů DNA (a jejich komplementárního protipólu mRNA)
 triplet = sekvence 3 nukleotidů DNA (a jejich komplementárního protipólu mRNA)")
21
Geny a Gen je možné definovat jako úsek DNA (sekvence nukleotidových bází, eventuelně tripletů), které po transkripci a translaci kódují syntézu specifického proteinu nebo RNA. Geny se skládají z: kódujících sekvencí (nazývají se exony) nekódujících sekvencí (introny) regulační části (promotor, místo počátku transkripce) koncových sekvencí Geny mohou mít různou délku (od cca 1000 až po několik set tisíc párů bází)
, které po transkripci a translaci kódují syntézu specifického proteinu nebo RNA. Geny se skládají z: kódujících sekvencí (nazývají se exony) nekódujících sekvencí (introny) regulační části (promotor, místo počátku transkripce) koncových sekvencí. Geny mohou mít různou délku (od cca 1000 až po několik set tisíc párů bází)")
22
RNA v eukaryotních buňkách se vyskytuje RNA v několika formách:
mediátorová RNA transferová RNA ribozómová RNA

23
Proteosyntéza = exprese genů
Proces, při kterém dochází k produkci proteinů na základě genetické informace uložené v DNA Je možné jej rozdělit do 2 dílčích částí: 1) Transkripce Nejprve je přepsána informace z DNA do m-RNA. Jedno vlákno dvoušroubovice DNA je při transkripci využito jako šablona, podle které RNA-polymerázy (syntetizující enzymy) vytvářejí m-RNA. Tato m-RNA následně přechází z jádra do cytoplazmy. Přitom prochází různými úpravami včetně vystřihání nekódujících sekvencí (intronů) a následného spojení exonů. Kódující m-RNA může být popsána jednotkami 3 nukleotidů, které se nazývají kodony (triplety).
 Transkripce. Nejprve je přepsána informace z DNA do m-RNA. Jedno vlákno dvoušroubovice DNA je při transkripci využito jako šablona, podle které RNA-polymerázy (syntetizující enzymy) vytvářejí m-RNA. Tato m-RNA následně přechází z jádra do cytoplazmy. Přitom prochází různými úpravami včetně vystřihání nekódujících sekvencí (intronů) a následného spojení exonů. Kódující m-RNA může být popsána jednotkami 3 nukleotidů, které se nazývají kodony (triplety).")
24
DNA Transkripce (přepis) matrice DNA do m-RNA Vzniká primární transkript m-RNA, následně podléhá úpravám (zejména vystři-hání nefunkčních intronů) Přechází z jádra do cytoplazmy DNA

25
Kodony Kodon (triplet) = skupina tří sousedních nukleotidů na mediátorové (informační) RNA, které svým pořadím určují kvalitu AK a její polohu v peptidickém řetězci
 = skupina tří sousedních nukleotidů na mediátorové (informační) RNA, které svým pořadím určují kvalitu AK a její polohu v peptidickém řetězci.")
26
tRNA Transferové tRNA jsou sestaveny z jednoduchého polynukleotidového řetězce, cca 80 nukleotidů Svým tvarem připomínají jetelový list Funkce tRNA je přenos jednotlivých aminokyselin, z nichž se polypeptidy skládají

27
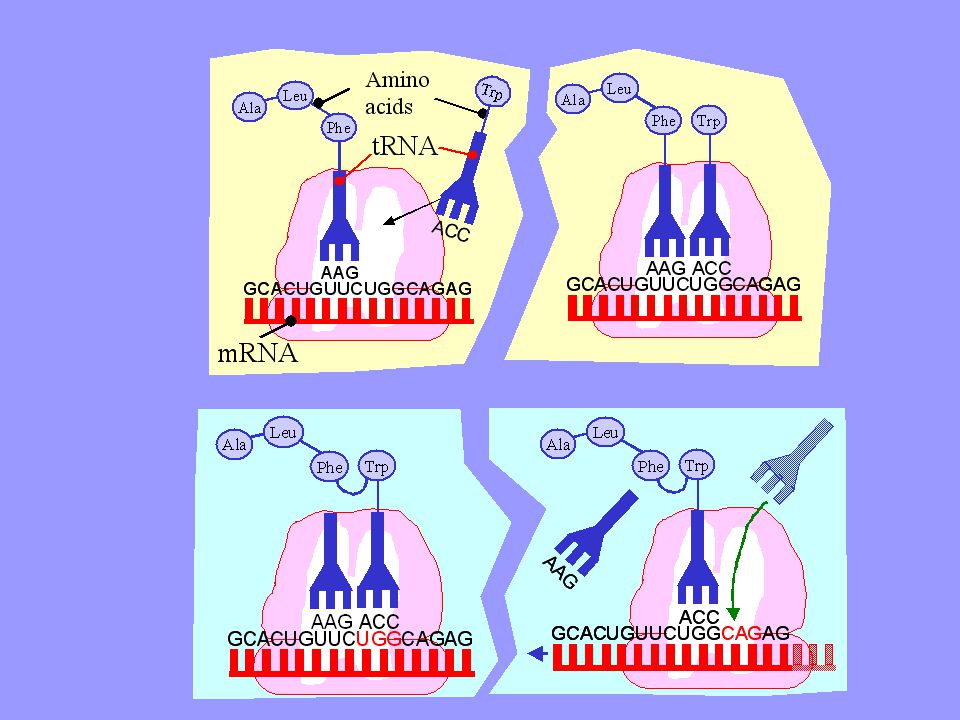
2) Translace = překlad proces překladu genetické informace z jazyka nukleotidů (kodonů) do pořadí aminokyselin v polypeptidovém řetězci Molekuly transferové t-RNA se svými antikodony (specifické sekvence 3 nukleotidů) vážou na komplementární kodony na mRNA. Translace (syntéza proteinů) začíná v místě iniciačního kodónu AUG (tomu odpovídá tRNA nesoucí AK methionin) a dále pokračuje ve smyslu kodónů AK se k sobě vážou za vzniku polypeptidu probíhá na ribozómech, které se přitom postupně pohybují podél řetězce m-RNA (po jednotlivých kodónech) Ukončení translace je zakódováno v kodónech UAA, UGA a UAG – po dosažení této sekvence dojde k uvolnění vytvořeného polypeptidu
 vážou na komplementární kodony na mRNA. Translace (syntéza proteinů) začíná v místě iniciačního kodónu AUG (tomu odpovídá tRNA nesoucí AK methionin) a dále pokračuje ve smyslu kodónů AK se k sobě vážou za vzniku polypeptidu. probíhá na ribozómech, které se přitom postupně pohybují podél řetězce m-RNA (po jednotlivých kodónech) Ukončení translace je zakódováno v kodónech UAA, UGA a UAG – po dosažení této sekvence dojde k uvolnění vytvořeného polypeptidu.")
29
Exprese genů = Proteosyntéza

30
Řetězec aminokyselin

31
Genetický kód DNA přenáší informaci do m-RNA ve formě kódu definovaného sekvencemi nukleotidových bází. - ribozómy se pohybují podél molekuly m-RNA a čtou její sekvence po 3 nukleotidech najednou (kodon). - kodón na m-RNA se páruje s komplementárním antikodónem na t-RNA každá AK je specifikována určitým kodónem (nebo více kodóny) na m-RNA RNA je konstruována ze 4 nukleotidů, existuje 64 možných kodónů (43) tři z nich specifikují ukončení polypeptidového řetězce (nekódují AK), nazývají se STOP kodóny pak na 61 kodónů připadá 20 různých aminokyselin.
. - kodón na m-RNA se páruje s komplementárním antikodónem na t-RNA - každá AK je specifikována určitým kodónem (nebo více kodóny) na m-RNA - RNA je konstruována ze 4 nukleotidů, existuje 64 možných kodónů (43) - tři z nich specifikují ukončení polypeptidového řetězce (nekódují AK), nazývají se STOP kodóny pak na 61 kodónů připadá 20 různých aminokyselin.")
32
The Genetic Code 2 A = adenin G = guanin C = cytosin T = thymin
U = uracil Ala: Alanin Cys: Cystein Asp: Asparagová kys. Glu: Glutamová kys. Phe: Fenylalanin Gly: Glycin His: Histidin Ile: Isoleucin Lys: Lysin Leu: Leucin Met: Methionin Asn: Asparagin Pro: Prolin Gln: Glutamin Arg: Arginin Ser: Serin Thr: Threonin Val: Valin Trp: Tryptofan Tyr: Tyrosin

33
Vlastnosti genetického kódu
Tripletový Základní jednotkou je trojice nukleotidů – triplet, kodón, která při translaci kóduje 1 AK Nepřekrývající se Záleží na začátku čtení (tj. od kterého nukleotidu se začne, neboť jinak by se změnil smysl informace), výjimka: viry Univerzální Smysl čtení tripletů platí pro veškeré živé organizmy Nadbytečný Většina z 20 AK je kódována více než 1 tripletem, u různých organizmů a různých typů NK může triplet kódovat odlišné kyseliny
, výjimka: viry. Univerzální. Smysl čtení tripletů platí pro veškeré živé organizmy. Nadbytečný. Většina z 20 AK je kódována více než 1 tripletem, u různých organizmů a různých typů NK může triplet kódovat odlišné kyseliny.")
34
Buněčný cyklus Cyklická reprodukce buněk od jednoho dělení ke druhému, zahrnuje 4 fáze: G1-fáze: Nastává po dokončení předešlého dělení a vzniku dceřiných buněk Probíhá syntéza RNA a proteinů, buňka v této fázi roste S-fáze (= syntetická) Probíhá replikace DNA, chromozómy jsou „zdvojeny“ vzniklé sesterské chromatidy jsou spojeny v místě centromery + syntéza RNA a proteinů (zejména těch, které tvoří chromatin – histony) jako v G1
 Probíhá replikace DNA, chromozómy jsou „zdvojeny vzniklé sesterské chromatidy jsou spojeny v místě centromery. + syntéza RNA a proteinů (zejména těch, které tvoří chromatin – histony) jako v G1.")
35
G2-fáze Příprava pro vlastní dělení buňky pokračuje růst, intenzívně se tvoří RNA a proteiny M-fáze Vlastní dělení buňky na 2 geneticky identické dceřiné buňky Zahrnuje 4 fáze: Profáze Metafáze Anafáze Telofáze

36
Mitóza 1) Profáze Rozpuštění jaderné membrány a jadérek, vznikají 2 centrioly - vzniká dělící vřeténko (mikrofilamenta, mikrotubuly), z chromatinu a jadérek vznikají pentlicovité chromozomy. (Touto dobou je již dávno po S fázi a veškerý genetický materiál je tudíž znásobený. Chromozomy jsou zdvojené, jsou ale stále spojeny v centroméře, než budou v anafázi roztrženy). 2) Metafáze Chromozomy se seřazují do rovníkové (ekvatoriální) roviny. Dělící vřeténko se navazuje na centromery chromozomů. Chromozomy zůstávají spojeny jen v centromerách. 3) Anafáze Roztržení chromozómů v centromerách zkracováním mikrotubulů dělícího vřeténka. Chromozomy putují k pólům buňky. 4) Telofáze Zánik dělícího vřeténka, despiralizace chromozomů, vzniká jaderná membrána a jadérka.
 Profáze Rozpuštění jaderné membrány a jadérek, vznikají 2 centrioly - vzniká dělící vřeténko (mikrofilamenta, mikrotubuly), z chromatinu a jadérek vznikají pentlicovité chromozomy. (Touto dobou je již dávno po S fázi a veškerý genetický materiál je tudíž znásobený. Chromozomy jsou zdvojené, jsou ale stále spojeny v centroméře, než budou v anafázi roztrženy). 2) Metafáze Chromozomy se seřazují do rovníkové (ekvatoriální) roviny. Dělící vřeténko se navazuje na centromery chromozomů. Chromozomy zůstávají spojeny jen v centromerách. 3) Anafáze Roztržení chromozómů v centromerách zkracováním mikrotubulů dělícího vřeténka. Chromozomy putují k pólům buňky. 4) Telofáze Zánik dělícího vřeténka, despiralizace chromozomů, vzniká jaderná membrána a jadérka.")
37
Meióza Meiotické neboli redukční dělení dává za vznik haploidních buněk (pohlavní buňky). Jejím cílem je tedy zajistit, aby buňka získala pouze polovinu genetického materiálu. Má 2 fáze, a to 1. a 2. meiotické dělení. 1. Meiotické dělení Homologní chromozomy tvoří v ekvatoriální rovině tzv. tetrády. Může mezi nimi dojít k rekombinaci genetického materiálu (crossing-over). Chromozomy nejsou roztrhávány, k pólům buňky putují celé sady. Na každém pólu tak zůstane vlastně 2krát jedna polovina gen. kódu. 2. Meiotické dělení Navazuje na první meiotické dělení. Mezi nimi již NEDOCHÁZÍ k další replikaci DNA. Probíhá téměř stejně jako normální mitóza. Výsledkem jsou tedy 4 dceřinné buňky, každá s jednou polovinou genetické výbavy.
. Jejím cílem je tedy zajistit, aby buňka získala pouze polovinu genetického materiálu. Má 2 fáze, a to 1. a 2. meiotické dělení. 1. Meiotické dělení Homologní chromozomy tvoří v ekvatoriální rovině tzv. tetrády. Může mezi nimi dojít k rekombinaci genetického materiálu (crossing-over). Chromozomy nejsou roztrhávány, k pólům buňky putují celé sady. Na každém pólu tak zůstane vlastně 2krát jedna polovina gen. kódu. 2. Meiotické dělení Navazuje na první meiotické dělení. Mezi nimi již NEDOCHÁZÍ k další replikaci DNA. Probíhá téměř stejně jako normální mitóza. Výsledkem jsou tedy 4 dceřinné buňky, každá s jednou polovinou genetické výbavy.")
38
Meióza

39
Od prarodičů k vnukům Přesně polovina genů od každého rodiče
V průměru jedna čtvrtina genů prarodičů, nikoliv úplně přesně

40
Klasifikace dědičnosti (Jinks 1967)
Genom chromozómy geny Genotyp plastidy plastogeny kinetostomy kinetogeny Plazmon mitochondrie chondriogeny centrioly centriogeny genotyp = celkový dědičný materiál buňky genom = celkový dědičný materiál uložený v chromozómech plazmon = celkový dědičný materiál uložený mimo chromozómy, tj. tj.v cytoplazmě

41
Podvojné založení dědičnosti
Znak je u diploida založen 2 formami genu – alelami (od otce a od matky), které jsou uloženy na příslušných lokusech homologních chromozómů, mohou být dominantní a recesivní Výjimka: dědičnost znaků vázaných na pohlaví Alely mohou být na homolo-gických chromozómech: Stejné kvality – homozygot v daném znaku Různé kvality – heterozygot v daném znaku A lokus B lokus A a
, které jsou uloženy na příslušných lokusech homologních chromozómů, mohou být dominantní a recesivní. Výjimka: dědičnost znaků vázaných na pohlaví. Alely mohou být na homolo-gických chromozómech: Stejné kvality – homozygot v daném znaku. Různé kvality – heterozygot v daném znaku. A lokus. B lokus. A. a.")
42
Mendelovy zákony a fenotypová pravidla
Pravidlo o uniformitě hybridů F1 generace a identitě reciprokých křížení Pravidlo o zákonitém štěpení v potomstvu hybridů F2 – princip náhodné segregace genů do gamet Pravidlo o volné (nezávislé) kombinovatelnosti vloh příklad dihybridní křížení) Pravidlo o čistotě gamet
 kombinovatelnosti vloh příklad dihybridní křížení) Pravidlo o čistotě gamet.")
43
Schématické znázornění monohybridního křížení
parentální generace: červenokvětý x bělokvětý P: BB x bb gamety v parentální generaci B x b F1: Bb křížení v rámci potomstva Bb x Bb F2: BB : 2Bb : bb genotypový štěpný poměr A a AA Aa aa

44
Při studiu dědičnosti Mendel použil hrách setý
Dominantní homozygot Recesivní homozygot Je samosprašný Sledoval pouze alternativní znaky Křížil čisté linie, tj. homozygoty v daných znacích (7 znaků, např. tvar semen, barva květů) P: AA x aa F1: Aa F2: AA : 2Aa : aa
 P: AA x aa. F1: Aa. F2: AA : 2Aa : aa.")
45
Schématické znázornění dihybridního křížení
P: AABB x aabb - matka vytváří gamety AB, otec ab F1: AaBb - v potomstvu se objevují pouze heterozygoti v obou znacích Následným křížením v rámci potomstva dochází k vyštěpování genotypů v přesných, tzv. genotypových štěpných poměrech ♀\♂ AB Ab aB ab AABB AABb AaBB Aabb AAbb AaBb aaBB aaBb aabb

46
Podmínky platnosti Mendelových zákonů
Jen pro monogenní znaky Jen pro úplnou dominanci alel Při úplné náhodnosti křížení Příslušné geny musí sídlit na autozómech, event. na homologických úsecích gonozómů

47
U lesních dřevin platí Mendelovy zákony jen v omezené míře

48
Vztahy mezi alelami 1 genu
Úplná dominance Dominantní alela svým účinkem převládá a stačí k úplnému projevu znaku (AA i Aa mají stejný fenotypový projev > aa) Neúplná dominance Dominantní alela u heterozygota plně nepře-krývá svým účinkem alelu recesivní, na fenotypu se projeví obě nestejnou intenzitou
 Neúplná dominance. Dominantní alela u heterozygota plně nepře-krývá svým účinkem alelu recesivní, na fenotypu se projeví obě nestejnou intenzitou.")
49
Kodominance U heterozygota dominují obě alely společně, jejich produkty jsou na sobě nezávislé (ve fenotypu se projeví obě alely plným projevem) Superdominance Znak u heterozygota je vyjádřen silněji než u obou homozygotů Toto může být mnohdy výhodné, a sice díky přítomnosti obou alel AA < Aa > aa

50
Inbreeding a heterozní efekt
křížení mezi blízkými příbuznými stupně uniformita v rámci populace – redukce heterozygotů o 50% v každé následné generaci ztráta vitality – projevení recesivity snížená produkce semen snížená adaptační schopnost Heterozní efekt převaha hybridů v porovnání s rodičovskými druhy

51
Použité zdroje http://genetika.wz.cz/
Dag Lindgren, Tomáš Funda - Základy obecné genetiky – PP prezentace Paule, L., 1992: Genetika a šĺachtenie lesných drevín. Príroda, 304 s.