Stáhnout prezentaci
1
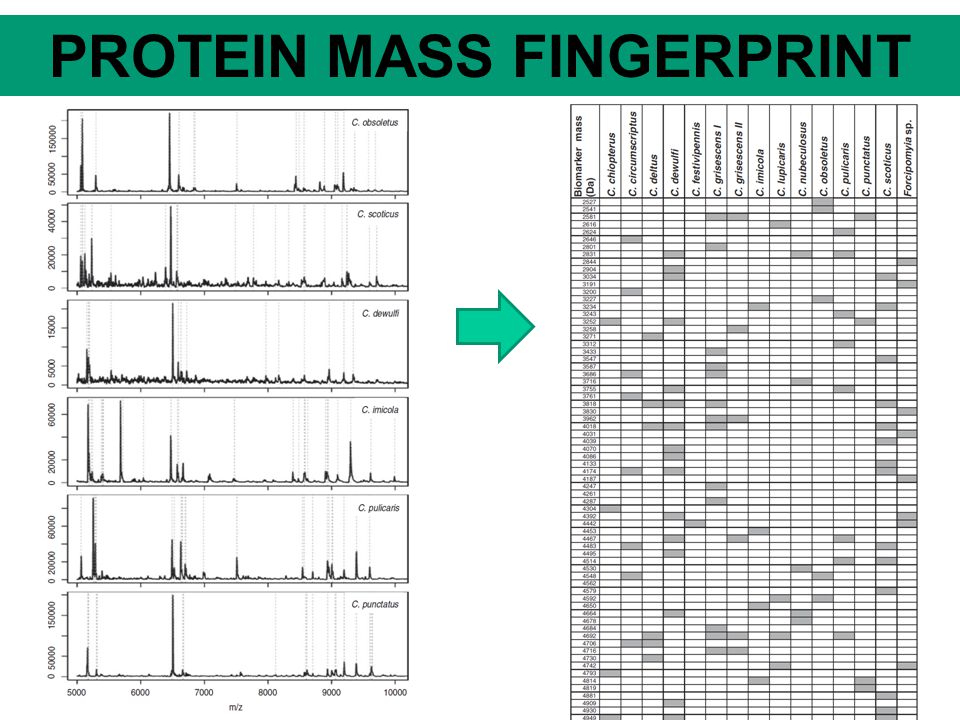
PROTEIN MASS FINGERPRINT

3
DNA/RNA MASS FINGERPRINT

4
SNPs Single Nucleotide Polymorphism Polymorfimus DNA, kdy se jedinci nebo druhy liší v jedné nukleotidové záměně AAGCCTA AAGCTTA V tomto případě mluvíme o alelách C a T. Téměž všechny SNPy mají jen 2 alely. Genom dvou lidí se liší zhruba v 3 mil. bazí (ne všechno jsou SNP).
..")
5
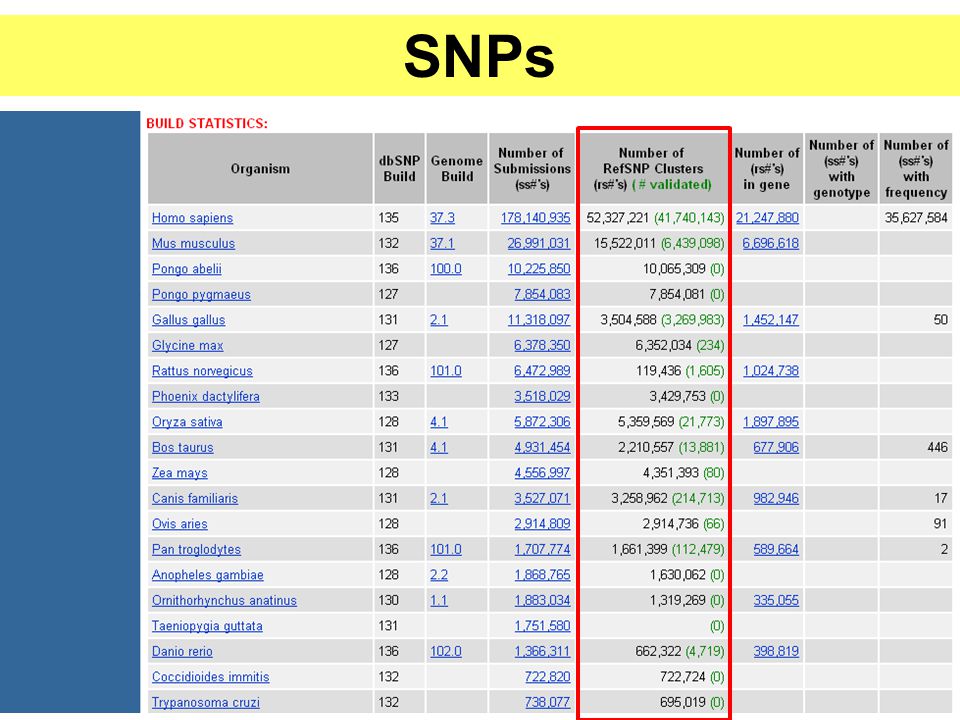
SNPs

7
SNPs genotypizace Molecular beacon Hybridizační metody Enzymatické metody Metody založené na fyzikálních vlastnostech DNA Hybridizační metody

8
SNPs – hybridizační metody 906 600 lidských SNPs

9
SNPs – enzymatické metody Primer extension – např. Infinium (Illumina) http://www.illumina.com/technology/ infinium_hd_assay.ilmn
 infinium_hd_assay.ilmn.")
10
SNP – OSTATNÍ METODY + + + _ _ _ denaturacePCRrenaturace vertikální elektroforéza vizualizace Single-Strand Conformation Polymorphism

11
SROVNÁNÍ METOD Fingerprinting (“otisk prstu DNA”) – souhrné označení pro metody poskytující velice variabilní elektroforetické vzory – VNTR RFLP, RAPD, AFLF. Čistě distanční Znakové Proteinové DNA Protein mass fingerprinting

12
SROVNÁNÍ METOD MetodaDNA hybri Dizace Mass fingerpti nting Mikro satelity RFLF (VNTR) SINERAPDAFLPSNP Počet lokusů VšechnyMnohoJedenMnohoJedenMnoho Jeden / mnoho Repliko vatelnost RůznáVysoká Různávysoká Povaha znaků DistanceKodomin antní Kodomi nantní? Vzácná událost Domi nantní Kodomi nantní? Kodo minantní RozlišeníStřednívysokéVysoké NízkéStředníVysoké Jedno duchost provedení TěžkéJedno Duché TěžkéJedno duché TěžkéJedno duché Střední- Doba trvání StředníKrátkáDlouháKrátkáDlouháKrátká -

13
ODHAD GENETICKÉ VZDÁLENOSTI

14
DISTANCE Z PODOBNOSTI VZORŮ RAPD/RFLP Koeficient genetické vzdálenosti dle. Nei a Li 1979, PNAS 76, 1979 Pro každou dvojici (x, y) spočteme všechny fragmenty (M x, M y ) a dále fragmenty vyskytující se v obou elektroforetogramech ( M xy ) Vypočteme podíl shodných fragmentů I = 2M xy /(M x + M y ) a z něj distanci D= 1- I X Y
 spočteme všechny fragmenty (M x, M y ) a dále fragmenty vyskytující se v obou elektroforetogramech ( M xy ) Vypočteme podíl shodných fragmentů I = 2M xy /(M x + M y ) a z něj distanci D= 1- I X Y.")
15
DISTANCE Z FREKVENCE ALEL Rogersova vzdálenost (pro všechny alely i lokusu v populacích X A a X B ) D= (0,5 Σ(x Ai - x Bi ) 2 ) 0,5 Vzdálenost Cavali-Svorza a Edwardse (1967) (pro všechny alely u lokusu v populacích X a Y) V případě, že použijeme více lokusů bude celková vzdálenost průměrem vzdáleností lokusů. Reynoldsova distance (1983) nebo Neiova distance (1972, 1978) berou v poraz biologické procesy.
 nebo Neiova distance (1972, 1978) berou v poraz biologické procesy..")
16
VÝPOČET „p“ p = ΔTm. 0,01 (0,015) p = podíl rozdílných nukleotidů mezi sekvencemi 2 taxonů Odhad p z reasociační kinetiky DNA-DNA hybridizace p = n d /n AATGTAGGAATCGC ACTGAAAGAATCGC p = 3/14 = 0,21
 p = podíl rozdílných nukleotidů mezi sekvencemi 2 taxonů Odhad p z reasociační kinetiky DNA-DNA hybridizace p = n d /n AATGTAGGAATCGC ACTGAAAGAATCGC p = 3/14 = 0,21.")
17
FREKVENCE SUBSTITUCÍ JE VYŠŠÍ NEŽ „p“ K = 12, p = 3 Jednoduchá substituce Vícenásobná substituce Zpětná substituce CTCT TCTC GTCTCT Koincidence ACTGAACGTAACGC Vidíme jen 3 rozdíly (p=3/14), ale ve skutečnosti došlo ke dvanácti substitucím (D = 12/14).
, ale ve skutečnosti došlo ke dvanácti substitucím (D = 12/14).")
18
Skutečný počet substitucí na jednu pozici Počet pozorovaných rozdílů 0,75 DNA 0,95 PROTEINY SUBSTITUČNÍ SATURACE p

19
Sekvence A Sekvence B ut Substituční rychlostČasový interval Sekvence A - AATGTAGGAATCGC Sekvence B - ACTGAAAGAATCGC ODHAD POČTU SUBSTITUCÍ

20
A G C T u/3 u = substituční rychlost u/3 = rychlost substituce za jednu konkrétní bázi (např. A -> G) u/3 Rychlost substituce za kteroukoli bázi i sebe sama 4/3 u Očekávaný počet substitucí za čas t 4/3 ut Jukes-Cantor příklad korekce na mnohonásobné substituce
 u/3 Rychlost substituce za kteroukoli bázi i sebe sama 4/3 u Očekávaný počet substitucí za čas t 4/3 ut Jukes-Cantor příklad korekce na mnohonásobné substituce.")
21
Poissonovo rozdělení Rozdělení počtu výskytu málo pravděpodobných jevů Pravděpodobnost, že dojde právě ke k událostem je f(k, ℷ ) = ( ℷ k e - ℷ )/k! ∞ 25 let → 20 substitucí…pro 5 let ℷ =4 ℷ =4 Pravděpodobnost, že k žádné události nedojde (k=0) je e - ℷ
 je e - ℷ.")
22
A G C T u/3 Pravděpodobnost, že za čas t k žádné události nedojde e -4/3 ut Pravděpodobnost, že za čas t k události dojde 1- e -4/3 ut Pravděpodobnost, že dojde ke konkrétní události P (C|A) = 1/4 (1- e -4/3 ut ) Pravděpodobnost, že dojde ke změně p = 3/4 (1- e -4/3 ut ) u/3 Jukes-Cantor příklad korekce na mnohonásobné substituce
 = 1/4 (1- e -4/3 ut ) Pravděpodobnost, že dojde ke změně p = 3/4 (1- e -4/3 ut ) u/3 Jukes-Cantor příklad korekce na mnohonásobné substituce")
23
Sekvence A Sekvence B ut Substituční rychlostČasový interval Sekvence A - AATGTAGGAATCGC Sekvence B - ACTGAAAGAATCGC p = počet neshodných míst/ délka sekvence 1-p = „sequence identity“ ODHAD POČTU SUBSTITUCÍ

24
A G C T u/3 Pravděpodobnost, že dojde ke změně p = 3/4 (1- e -4/3 ut ) Korigovaný počet substitucí D = ut = -3/4 ln(1- 4/3 p) u/3 Jukes-Cantor příklad korekce na mnohonásobné substituce V(D) = (p(1 -p))/(L(1 - 4/3 p) 2 ) L = délka sekvence Rozptyl D:
 Korigovaný počet substitucí D = ut = -3/4 ln(1- 4/3 p) u/3 Jukes-Cantor příklad korekce na mnohonásobné substituce V(D) = (p(1 -p))/(L(1 - 4/3 p) 2 ) L = délka sekvence Rozptyl D:")
25
A G C T u/3 Korigovaný počet substitucí D = ut = -3/4 ln(1- 4/3 p) u/3 Jukes-Cantor příklad Příklad naší sekvence: D= -3/4 ln(1- 4/3 * 0,21) D= 0,246
 u/3 Jukes-Cantor příklad Příklad naší sekvence: D= -3/4 ln(1- 4/3 * 0,21) D= 0,246")
26
Jukes-Cantor Obecně použitelné odvození 4/3 ut u/3 -u u/3 -u u/3 -u A G Pravděpodobnost, že dojde ke konkrétní události P (C|A) = 1/4 (1- e -4/3 ut ) CT A G C T
 = 1/4 (1- e -4/3 ut ) CT A G C T")
27
Jukes-Cantor Součet řádku je 1 4/3 ut u/3 -u u/3 -u u/3 -u A GCT A G C T Suma = 1

28
Jukes-Cantor Nekonečně vzdálené sekvence se podobají z 1/4 Skutečný počet substitucí na jednu pozici Počet pozorovaných rozdílů 0,75 DNA p Sekvence A Sekvence B ut=∞ 4/3 ut

29
Jukes-Cantor Je symetrický u/3 -u u/3 -u u/3 -u A GCT A G C T Sekvence A Sekvence B

30
Jukes-Cantor Je symetrický u/3 -u u/3 -u u/3 -u A GCT A G C T Sekvence A Sekvence B

31
Kimura 2P Kimura 2 parametrový DALŠÍ MODELY A G C T α ββ β β α

32
Kimura 2P Kimura 2 parametrový DALŠÍ MODELY A G C T α ββ β β α

33
A G C T α ββ β β α Kimura 2P Kimura 2 parametrový DALŠÍ MODELY D = 0,5 ln(a) + 1/4 ln(b) a = 1/(1 - 2P - Q) b = 1/(1 -2Q) P – podíl transic Q – podíl transverzí V(D) = [a 2 P + c 2 Q -(aP +cQ) 2 ]/L c = (a + b)/2 L = délka sekvence Rozptyl D:
![A G C T α ββ β β α Kimura 2P Kimura 2 parametrový DALŠÍ MODELY D = 0,5 ln(a) + 1/4 ln(b) a = 1/(1 - 2P - Q) b = 1/(1 -2Q) P – podíl transic Q – podíl transverzí V(D) = [a 2 P + c 2 Q -(aP +cQ) 2 ]/L c = (a + b)/2 L = délka sekvence Rozptyl D:](http://images.slideplayer.cz/10/2697234/slides/slide_33.jpg "A G C T α ββ β β α Kimura 2P Kimura 2 parametrový DALŠÍ MODELY D = 0,5 ln(a) + 1/4 ln(b) a = 1/(1 - 2P - Q) b = 1/(1 -2Q) P – podíl transic Q – podíl transverzí V(D) = [a 2 P + c 2 Q -(aP +cQ) 2 ]/L c = (a + b)/2 L = délka sekvence Rozptyl D:")
34
A G C T α ββ β β α Kimura 2P Kimura 2 parametrový DALŠÍ MODELY D = 0,5 ln(a) + 1/4 ln(b) a = 1/(1 - 2P - Q) b = 1/(1 -2Q) Příklad naší sekvence: P=2/14=0,14 Q=1/14=0,07 a = 1/(1 – 2*0,14 – 0,07) = 1,54 b = 1/(1 -2*0,07) = 1,16 D = 0,5 ln(1,54) + 1/4ln(1,16)=0,254
 + 1/4 ln(b) a = 1/(1 - 2P - Q) b = 1/(1 -2Q) Příklad naší sekvence: P=2/14=0,14 Q=1/14=0,07 a = 1/(1 – 2*0,14 – 0,07) = 1,54 b = 1/(1 -2*0,07) = 1,16 D = 0,5 ln(1,54) + 1/4ln(1,16)=0,254")
35
DALŠÍ MODELY Rovnovážné frekvence nukleotidů π A π C π G π T F84

36
A G C T α εβ δ γ ζ GTR General time reversible + DALŠÍ MODELY Rovnovážné frekvence nukleotidů π A π C π G π T Parametry: frekvence (rychlost) záměn ( αβγδεζ ) a frekvence nukleotidů ( π A π C π G π T ) se odvozují z analyzovaných sekvencí.
 záměn ( αβγδεζ ) a frekvence nukleotidů ( π A π C π G π T ) se odvozují z analyzovaných sekvencí.")
37
RŮZNÉ MODELY FUNGUJÍ RŮZNĚ Skutečný počet substitucí Odhad počtu substitucí

38
d xy = -ln (det F xy ) Sekvence A Sekvence B F xy = [ ] 0,2490,0060,0270,009 0,0030,1660,0010,018 0,0270,0060,2560,004 0,0060,0210,0090,194 Alignment 900 pozic d xy = -ln (det F xy ) = -ln (0.002) = 6,216 LogDet distance
![d xy = -ln (det F xy ) Sekvence A Sekvence B F xy = [ ] 0,2490,0060,0270,009 0,0030,1660,0010,018 0,0270,0060,2560,004 0,0060,0210,0090,194 Alignment 900 pozic d xy = -ln (det F xy ) = -ln (0.002) = 6,216 LogDet distance](http://images.slideplayer.cz/10/2697234/slides/slide_38.jpg "d xy = -ln (det F xy ) Sekvence A Sekvence B F xy = [ ] 0,2490,0060,0270,009 0,0030,1660,0010,018 0,0270,0060,2560,004 0,0060,0210,0090,194 Alignment 900 pozic d xy = -ln (det F xy ) = -ln (0.002) = 6,216 LogDet distance")
39
JAKÉ DISTANCE POUŽÍVAT? Modely s větším počtem parametrů (GTR) jsou flexibilnější a většinou přesnější než metody jednoduché. Potřebují však velké množství parametrů a distance jimi vypočítané mají větší rozptyl. Pro kratší úseky proto dávají horší výsledky. Simulace ukázala, že pro D< 0,5 dávají poměrně přesné výsledky i nejjednoduší metody (Jukes-Cantor, Kimura). Pro větší distance (a dostatečně dlouhé sekvence) je lépe použít složitější modely (GTR).
 jsou flexibilnější a většinou přesnější než metody jednoduché. Potřebují však velké množství parametrů a distance jimi vypočítané mají větší rozptyl. Pro kratší úseky proto dávají horší výsledky. Simulace ukázala, že pro D< 0,5 dávají poměrně přesné výsledky i nejjednoduší metody (Jukes-Cantor, Kimura). Pro větší distance (a dostatečně dlouhé sekvence) je lépe použít složitější modely (GTR)..")
















 – maximální zisk 10 bodů - k ruce můžete mít jakékoli materiály - kalkulačka.>")



