Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Získání dat Sběr vzorků Izolace DNA
Výběr vhodného márkru (co už bylo publikováno pro srovnání, dostupnost, mutační rychlost, atd.)
")
2
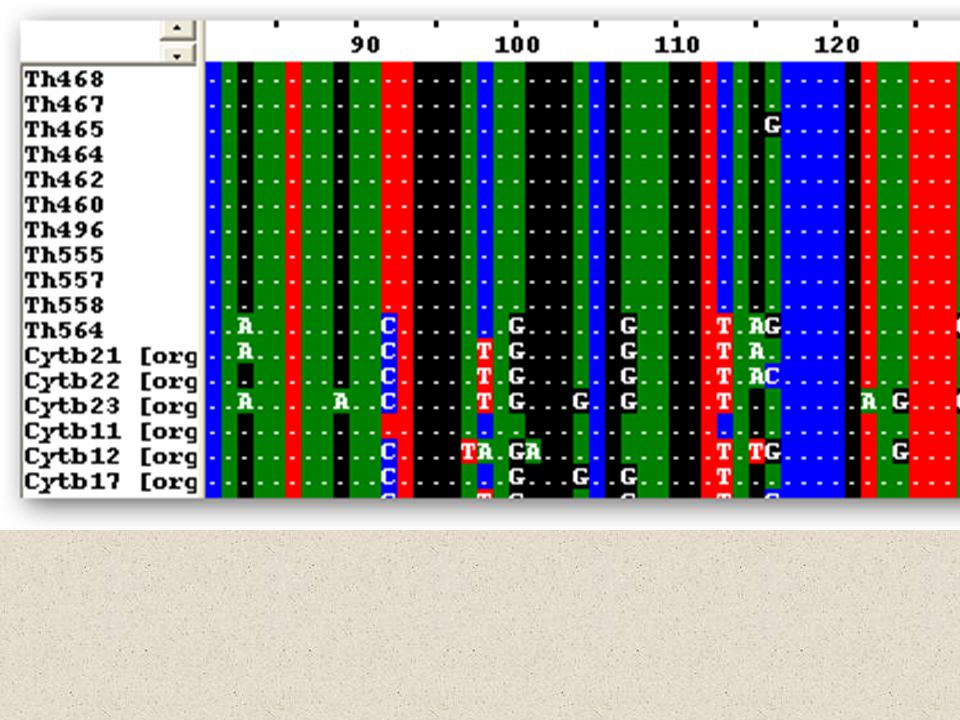
Velká vs. Malá fylogenetika
malé rozdíly mezi sekvencemi nízká podpora kládů často v rámci reproductive units – možnost introgrese

5
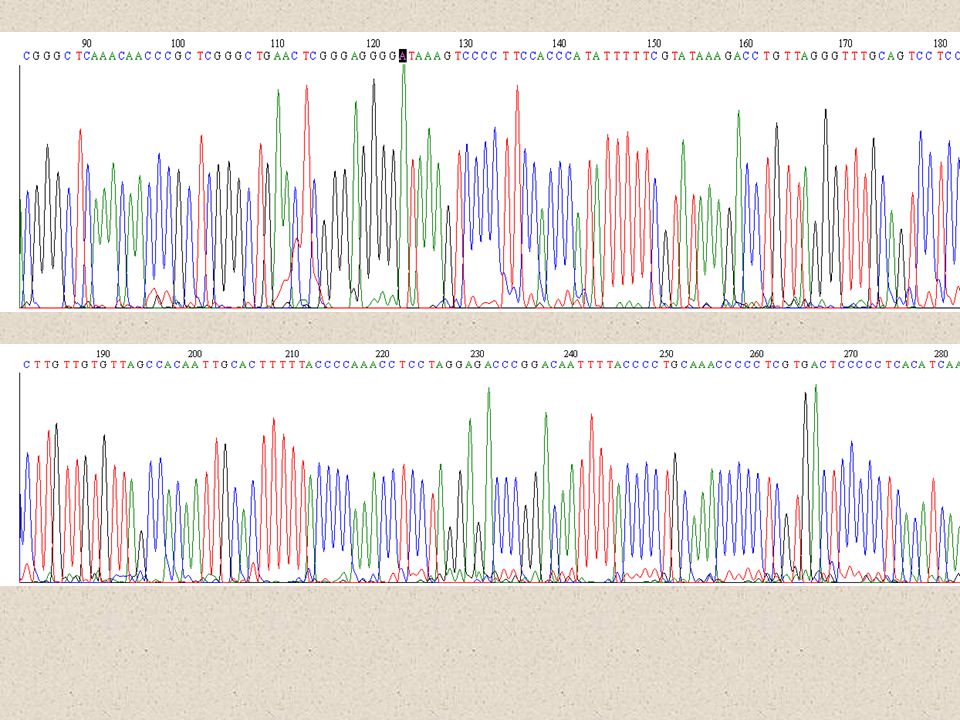
Alignment

6
Alignment

7
Alignment

8
Alignment

9
Alignment Homologizace pozic
Každý mismatch má jistou „penalty“, totéž platí pro otevření Gapu a pro prodlužování Gapu V jednoduchých případech možno okometricky Standardně se používá specializovaný software, např. CLUSTAL W.

10
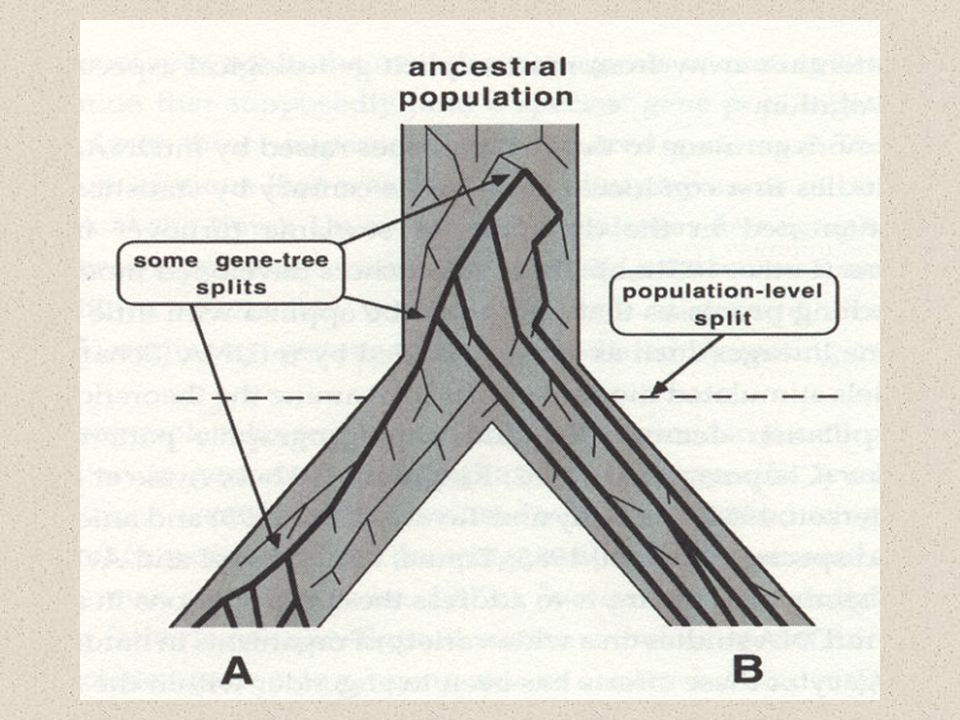
Velká vs. Malá fylogenetika
malé rozdíly mezi sekvencemi nízká podpora kládů často v rámci reproductive units – možnost introgrese nutno odlišovat „gene trees“ a „species trees“

12
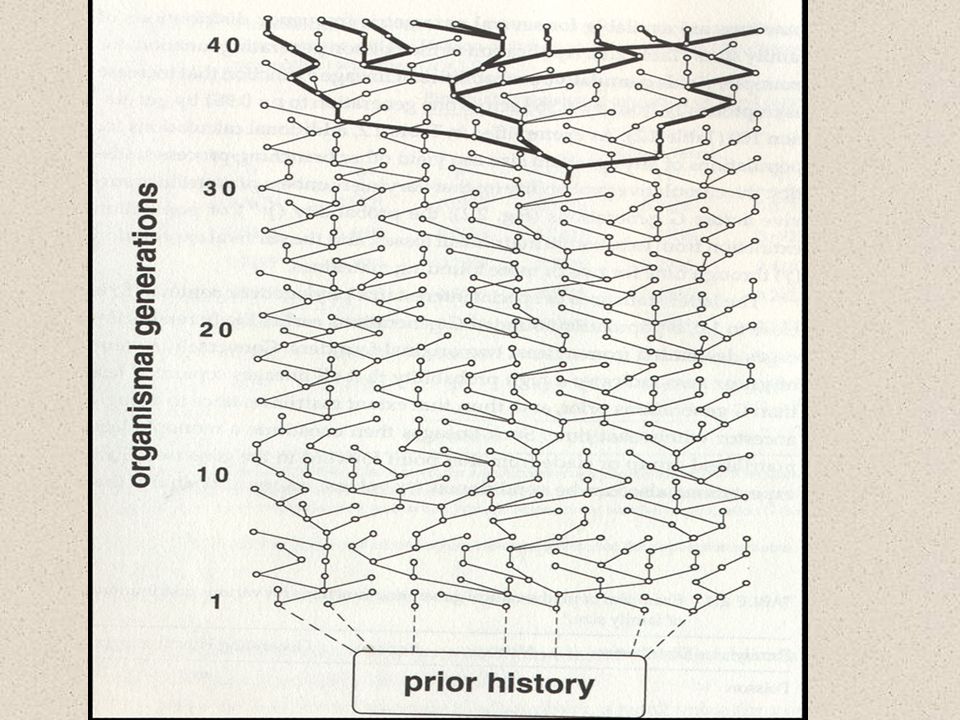
Koalescenční teorie proč jsou si alely v jednom druhu vzájemně bližší, než v jiném druhu? Jaký je vztah genetické variability k velikosti populace? A k populační historii? A ke geografické struktuře?

13
Koalescenční teorie odvození z pravděpodobnosti přežívání linií

15
Koalescenční teorie Pravděpododnost společného předka dvou jedinců: 1/2N Pravděpodobnost odlišných předků k jedinců: (1-1/2N)(1-2/2N)…(1-(k-1)/2N) Pravděpodobná koalescence: 1-Gkk = k(k-1)/4N
(1-2/2N)…(1-(k-1)/2N) Pravděpodobná koalescence: 1-Gkk = k(k-1)/4N.")
16
Koalescenční teorie Jak dlouho to trvá?
4N/k(k-1)+ 4N/(k-1)(k-2) +…+4N/2 4N (1-1/k)
+ 4N/(k-1)(k-2) +…+4N/2. 4N (1-1/k)")
17
Koalescenční teorie dvě náhodné kopie genu budou sdílet MRCA průměrně za 2N generací za 4N generací by měly všechny linie splynout

18
Koalescenční teorie

19
Koalescenční teorie - předpoklady
Stabilní populace Neutralita znaku Geografická homogeneita

20
Výpočet populační velikosti
Podle principu koalescenční teorie nutno znát mutační rychlost Populace se dá charakterizovat parmetrem Θ (Θ = 4Ne * u)
")
22
Výpočet populační velikosti Jaké překvapení
Census versus effective population size Vysvětlení se hledalo v historických změnách demografických parametrů v selekčním tlaku v rozdílných vlivech „life history“ na různé márkry: mtDNA vs. Y chromozóm vs. autozómy

23
Změny populační velikosti
Klimatické cykly, infekce atd. Vliv na dobu koalescence population bottlenecks během fragmentace i rekolonizace

24
Selekce Jak ji změřit? Negativní Pozitivní Ve prospěch heterozygotů
dN/dS poměr Negativní Pozitivní Ve prospěch heterozygotů Vliv na dobu koalescence

25
Vliv „life history“ Rozdílná migrace samců a samic
velká variance v reprodukční úspěšnosti

26
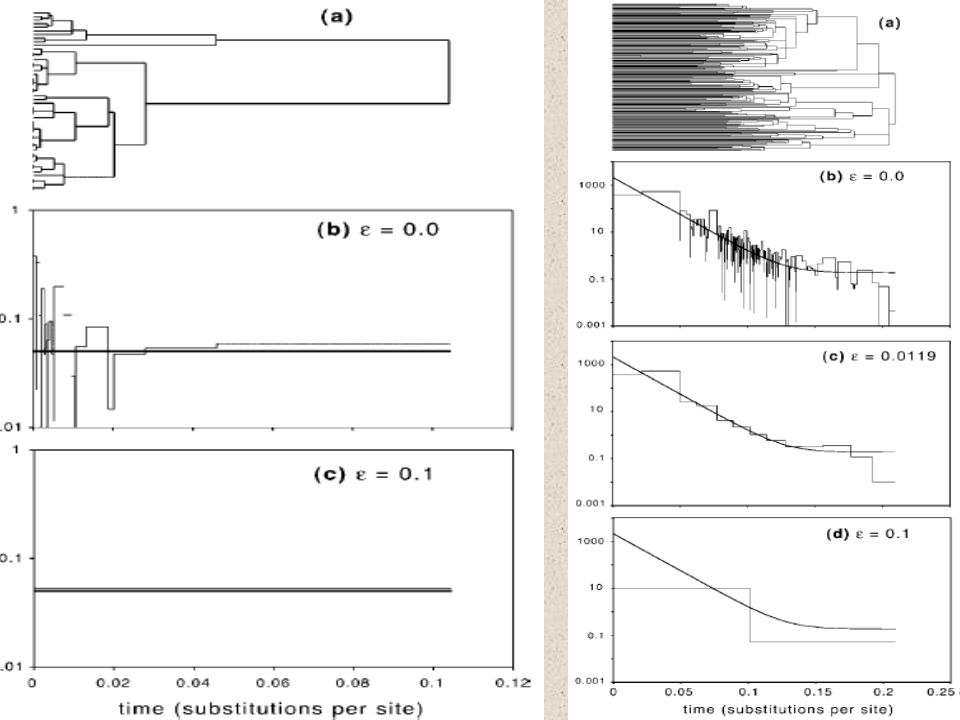
Odhad demografické historie
Mismatch distribution histogram četností mutačních rozdílů v jednotlivých dvojicích vzorků (DNAsp, Arlequin)
")
27
Mismatch distribution

28
Mismatch distribution

29
Odhad demografické historie
Mismatch distribution

30
Odhad demografické historie
Mismatch distribution

32
Odhad demografické historie
Lineages through time rozložení počtu genealogických linií v čase. (Genie)
")
34
Odhad demografické historie
Skyline plots zlepšný algoritmus Lineage Through Time. Počty linií jsou porovnávány v optimalizovaných časových intervalech (Genie)
")
36
Odhad demografické historie
h/p poměr a z něj odvozené indexy jako Tajima’s D, Fu’s Fs Tyto metody jsou založeny na poměru „starého“ vs. „mladého“ polymorfismu. V expandovaných populacích (převaha „mladého“) nabývají záporných hodnot (DNAsp, Arlequin)
 nabývají záporných hodnot. (DNAsp, Arlequin)")
37
Odhad demografické historie
Maximum Likelihood odhady populačních parametrů Porovnává se pravděpodobnost pozorované topologie Genetree s navrženými parametry populačního modelu (např. Θ a g, kde θt = θinitial –gt ) Parametry se postupně „vylepšují“ až se najdou ty, které maximalizují pst pozorování (Fluctuate, Lamarc, Sweep-Botleneck)
 Parametry se postupně „vylepšují až se najdou ty, které maximalizují pst pozorování. (Fluctuate, Lamarc, Sweep-Botleneck)")
38
Datování událostí Klasické molekulární hodiny
LRT test linearised tree ML test local rates v každém uzlu a optimalisace parametrů Malá divergence – nutno brát v potaz lineage sorting a stochasticitu akumulace mutací

39
Datování událostí Malá divergence – nutno brát v potaz lineage sorting a stochasticitu akumulace mutací tradiční Neiův přístup Moment methods ML methods

40
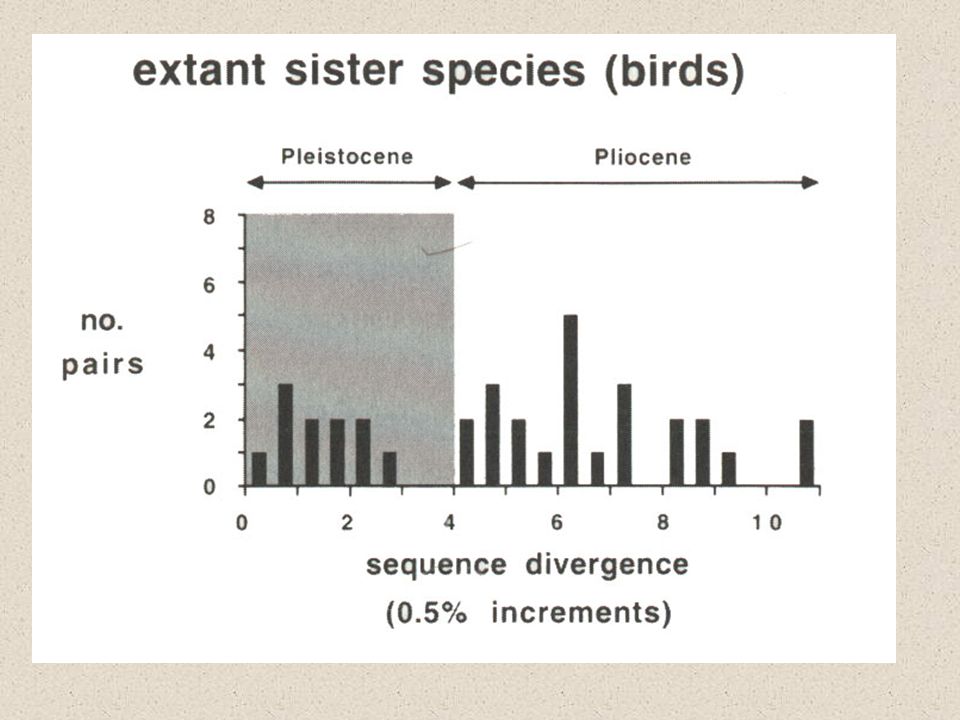
Vliv Pleistocénních glaciací na speciaci
mtDNA divergence predatuje Pleistocén druhy však do dob ledových vstupovaly už strukturovány

42
Complete glaciation of the shelf

43
And its impact on shelf and slope biota

44
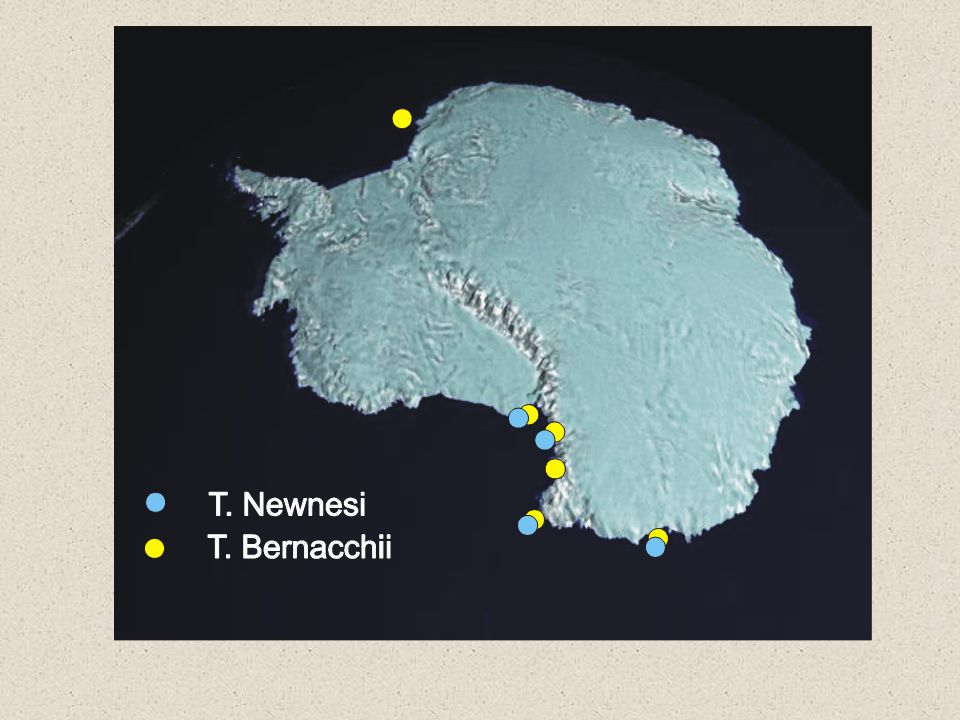
Notothenioidei-suitable model
two adaptive radiations first likely in relation to miocen coldering (cca 24 MY) second cca 3.5 MY- fam. Trematomidae
 second cca 3.5 MY- fam. Trematomidae.")
45
Trematomus project study the speciation mechanisms in the unique marine species-flock envision the geographical structuration of shelf fishes evaluate the role of climatic changes on the evolution of Antarctic biota appreciate the roles of intrinsic factors in shaping the population structure

50
Conclusions Glacial cycles apparently had significant impact on some but not all shelf organisms life history likely modifies the effects of these dramatic fluctuations presently we do not witness any ongoing speciation within studied species

Podobné prezentace













![Genetické algoritmy [GA]](/8/2469778/big_thumb.jpg "Genetické algoritmy [GA]>")








>")


 Stochastické.>")
