Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
KLONOVÁNÍ a GENOVÉ INŽENÝRSTVÍ

2
KLONOVÁNÍ a GENOVÉ INŽENÝRSTVÍ
úvodní přednáška PCR techniky mutageneze in vitro cDNA a genomové knihovny sekvencování genomů transgenoze rostlin genové čipy transgenoze živočichů genová terapie klonovací strategie a expresní systémy seminář – nástroje molekulární biologie v praxi

3
Získávání bioinformací (dobrovolný seminář)
NCBI (National Center for Biological Information) TIGR (The Institute for Genomic Research) SignalP, TargetP (predikce buněčné lokalizace proteinů) BIOEDIT (freeware pro manipulaci s DNA a proteiny) BLAST (basic local alighment and search tool) GENEVESTIGATOR (sledování exprese genů na úrovni celých genomů in silico)
 TIGR (The Institute for Genomic Research) SignalP, TargetP (predikce buněčné lokalizace proteinů) BIOEDIT (freeware pro manipulaci s DNA a proteiny) BLAST (basic local alighment and search tool) GENEVESTIGATOR (sledování exprese genů na úrovni celých genomů in silico)")
4
LITERATURA Molecular Biotechnology Molecular Cloning 1,2,3
Glick R a Pasternak JJ Molecular Biotechnology 3th edition, ASM Press Ltd, Washington, USA, 2003 Sambrook J a kol. Molecular Cloning 1,2,3 3th edition, Cold Spring Harbor Laboratory Press, New York, USA 2001 Rosypal S Úvod do Molekulární Biologie díl čtvrtý třetí vydání, Prof.RNDr. Stanislav Rosypal, DrSc, Brno 2002 Brown TA; překlad Fellner M a kol. Klonování genů a analýza DNA 1 české vydání, Olomouc UP 2007

5
Jeden gen jeden enzym (protein/polypeptid)
Centralní dogma DNA RNA Protein DNA do DNA REPLIKACE DNA do RNA TRANSKRIPCE RNA do Protein TRANSLACE RNA do DNA REVERZNÍ TRANSKRIPCE Fenotyp Jeden gen jeden enzym (protein/polypeptid)
")
6
AAGCTCGACTAGCGCTAGCCTACCTAGCTCTCCCCTTCC
Centralní dogma 5’ C AAA 3’ 3’ N Gen 9,200 bp zralá RNA 1,870 bp Protein 386 aa CDS 1158 bp (coding sequence) AAGCTCGACTAGCGCTAGCCTACCTAGCTCTCCCCTTCC (kodující sekvence ORF (open reading frame) a nepřekládané regiony – 5´-UTR, 3´-UTR)
 AAGCTCGACTAGCGCTAGCCTACCTAGCTCTCCCCTTCC. (kodující sekvence ORF (open reading frame) a nepřekládané regiony – 5´-UTR, 3´-UTR)")
7
Struktura genu Introny a Exony Promotor Enhancer a Silencer
Počátek transkripce Transkripční faktory Signální sekvence Introny a Exony: Kodující sekvence (ORF) mnoha eukaryotických genů je přerušená sekvencí známou jako introny. Introny jsou úseky DNA jejichž transkripty nejsou přítomný ve zralé mRNA. Exony jsou části sekvence přítomné ve zralé mRNA a kodují produkt eukaryotického genu.
 mnoha eukaryotických genů je přerušená sekvencí známou jako introny. Introny jsou úseky DNA jejichž transkripty nejsou přítomný ve zralé mRNA. Exony jsou části sekvence přítomné ve zralé mRNA a kodují produkt eukaryotického genu.")
8
Místo počátku transkripce: místo kde začíná syntéza mRNA (pozice 0, -30bp od TATA boxu) .
Promotor: Oblast genu (5’-konec) upstream od místa počátku transkripce sloužící jako templát pro navázání transkripční jednotky a iniciaci transkripce. Inducibilní promotor – je závislý na přítomnosti induktoru (nízkomolekulární látka). Konstitutivní promotor – nezávislý (provozní „house keeping“ geny). Enhancer: Část sekvence genu která interaguje s transkripčními faktory a tak zvyšuje účinnost transkripce. Je umístěna upstream nebo downstream od kódující sekvence a může být stovky bází daleko od kódující sekvence kterou kontroluje. Silencer: Opak enhanceru. Transkripční faktory: Proteiny interagující se specifickou sekvenci DNA, většinou ovlivňující terciální strukturu DNA a tak regulují transkripční jednotku. Jsou buď aktivátory nebo represory viz výše. Koaktivátory: Proteiny interagující s transkripčními faktory (ale ne s DNA) regulující transkripci. Signální sekvence: kóduje signální peptid, který předurčuje zacílení proteinu na buněčné úrovni a následně se odštěpuje.
 upstream od místa počátku transkripce sloužící jako templát pro navázání transkripční jednotky a iniciaci transkripce. Inducibilní promotor – je závislý na přítomnosti induktoru (nízkomolekulární látka). Konstitutivní promotor – nezávislý (provozní „house keeping geny). Enhancer: Část sekvence genu která interaguje s transkripčními faktory a tak zvyšuje účinnost transkripce. Je umístěna upstream nebo downstream od kódující sekvence a může být stovky bází daleko od kódující sekvence kterou kontroluje. Silencer: Opak enhanceru. Transkripční faktory: Proteiny interagující se specifickou sekvenci DNA, většinou ovlivňující terciální strukturu DNA a tak regulují transkripční jednotku. Jsou buď aktivátory nebo represory viz výše. Koaktivátory: Proteiny interagující s transkripčními faktory (ale ne s DNA) regulující transkripci. Signální sekvence: kóduje signální peptid, který předurčuje zacílení proteinu na buněčné úrovni a následně se odštěpuje.")
9
DNA DNA DNA polymerasy blíže přednáška techniky PCR

10
DNA RNA Transkripce (RNA syntéza): přenos informace z DNA do RNA (nestabilní) prostřednictvím enzymu RNA Polymerasy (DNA-dependentní RNA Polymerasa). Syntéza RNA je komplementární k templátovému DNA duplexu a je identická s netemplátovým vláknem duplexu (5´-3´). Netemplátový řetězec je proto nazýván kódující řetězec a je identický s mRNA s výjimkou záměny U za T. Typy RNA: messenger RNA (mRNA, polyA+ RNA). RNA která je přepisována do polypeptidů (1-5%). ribozomální RNA (rRNA). strukturní složka ribozómů. (90%) transferová RNA (tRNA). přenašeč aminokyselin do místa syntézy proteinu. (5-10%) málá jaderná, jadérková, cytoplazmatická RNA (snRNA/sno/scRNA). sestřih mRNA
: přenos informace z DNA do RNA (nestabilní) prostřednictvím enzymu RNA Polymerasy (DNA-dependentní RNA Polymerasa). Syntéza RNA je komplementární k templátovému DNA duplexu a je identická s netemplátovým vláknem duplexu (5´-3´). Netemplátový řetězec je proto nazýván kódující řetězec a je identický s mRNA s výjimkou záměny U za T. Typy RNA: messenger RNA (mRNA, polyA+ RNA). RNA která je přepisována do polypeptidů (1-5%). ribozomální RNA (rRNA). strukturní složka ribozómů. (90%) transferová RNA (tRNA). přenašeč aminokyselin do místa syntézy proteinu. (5-10%) málá jaderná, jadérková, cytoplazmatická RNA (snRNA/sno/scRNA). sestřih mRNA.")
11
Postranskripční úpravy mRNA
Čepička (capping). modifikovaný guanozin (m7G) je přidáván na 5'-konec většiny mRNA. Stabilita mRNA a podílí se na vazbě k ribozomu. Polyadenylace bp dlouhá sekvence polyadenozinu je přidávána na 3'-konec většiny eukaryotické pre-mRNA. PolyA konec není kodován v DNA. Sestřih intronů a exonů. introny jsou vyštěpeny a exony spojeny dohromady ve struktuře zvané spliceozom. Y pyrimidinová báze
. modifikovaný guanozin (m7G) je přidáván na 5 -konec většiny mRNA. Stabilita mRNA a podílí se na vazbě k ribozomu. Polyadenylace bp dlouhá sekvence polyadenozinu je přidávána na 3 -konec většiny eukaryotické pre-mRNA. PolyA konec není kodován v DNA. Sestřih intronů a exonů. introny jsou vyštěpeny a exony spojeny dohromady ve struktuře zvané spliceozom. Y pyrimidinová báze.")
12
Postranskripční úpravy mRNA

13
RNA protein Translace (syntéza proteinů): Translace je proces přeměny informace z RNA do aminokyselinového řetězce (polypeptidu) probíhající v cytoplasmě. Zralá mRNA je transportována do cytoplasmy k ribozomům (mnohdy vázané na endoplasmatickou retikulární síť). Ribozóm se skládá ze strukturní rRNA a 80 různých proteinů, které tvoří malou a velkou podjednotku. Iniciace: Zachycení mRNA a nalezení start kodonu AUG, který koduje methionin. (M je odštěpen během postranslačních úprav nebo společně se signálním peptidem) Elongace: Přidávání různých AK v aktivované formě vázané na tRNA a vytváření peptidové vazby mezi amino a karboxylovou skupinou. Terminace: STOP kodón (UAA, UAG nebo UGA) Posttranslační úpravy: Fosforylace, glykosylace, organelové cílení a odštěpení signálního peptidu, složení do aktivního stavu (vytvoření terciální a kvartérní struktury).
: Translace je proces přeměny informace z RNA do aminokyselinového řetězce (polypeptidu) probíhající v cytoplasmě. Zralá mRNA je transportována do cytoplasmy k ribozomům (mnohdy vázané na endoplasmatickou retikulární síť). Ribozóm se skládá ze strukturní rRNA a 80 různých proteinů, které tvoří malou a velkou podjednotku. Iniciace: Zachycení mRNA a nalezení start kodonu AUG, který koduje methionin. (M je odštěpen během postranslačních úprav nebo společně se signálním peptidem) Elongace: Přidávání různých AK v aktivované formě vázané na tRNA a vytváření peptidové vazby mezi amino a karboxylovou skupinou. Terminace: STOP kodón (UAA, UAG nebo UGA) Posttranslační úpravy: Fosforylace, glykosylace, organelové cílení a odštěpení. signálního peptidu, složení do aktivního stavu. (vytvoření terciální a kvartérní struktury).")
14
RNA DNA Reverzní transkripce: Proces vytvoření DNA z RNA templátu za pomocí RNA-dependentní DNA polymerasy – Reverzní transkriptasa (RT). RT je přítomná pouze v RNA virech avian myeloblastosis virus (AMV), Moloney murine leukemia virus (MMLV), nebo human immunodeficiency virus (HIV). Aplikace: tvorba cDNA (komplementární DNA): přesná DNA kopie mRNA. DNA sekvence proteinu bez přerušení introny. cDNA knihovny RT PCR
, Moloney murine leukemia virus (MMLV), nebo human immunodeficiency virus (HIV). Aplikace: tvorba cDNA (komplementární DNA): přesná DNA kopie mRNA. DNA sekvence proteinu bez přerušení introny. cDNA knihovny. RT PCR.")
15
Hybridizace Dva řetězce DNA mohou být k sobě komplementární.
mRNA syntetizována z genu je identická k jednomu vláknu svého genu a komplementární k druhému vláknu. Komplementární řetězce mohou být odděleny (denaturace) a znovu spojeny (renaturace – hybridizace) na různém stupni specifity, mohou vznikat hybridní formace DNA-DNA, DNA-RNA, nebo RNA-RNA. Methody využívající těchto vlastnosti: Southern blot: Hybridizace DNA na DNA (v pevné fázi) Northern blot: Hybridizace DNA na RNA (v pevné fázi) RNAse protection assay: Hybridizace RNA na RNA (v roztoku) In situ hybridizace DNA-DNA nebo DNA-RNA (ve tkání, či v buňce, např. FISH) izolace mRNA: vazba polyA konce na polyT sekvenci navázanou na pevný matrix. Stringence: podmínky zaručující stupeň specifity (komplementarity)
 a znovu spojeny (renaturace – hybridizace) na různém stupni specifity, mohou vznikat hybridní formace DNA-DNA, DNA-RNA, nebo RNA-RNA. Methody využívající těchto vlastnosti: Southern blot: Hybridizace DNA na DNA (v pevné fázi) Northern blot: Hybridizace DNA na RNA (v pevné fázi) RNAse protection assay: Hybridizace RNA na RNA (v roztoku) In situ hybridizace DNA-DNA nebo DNA-RNA (ve tkání, či v buňce, např. FISH) izolace mRNA: vazba polyA konce na polyT sekvenci navázanou na pevný matrix. Stringence: podmínky zaručující stupeň specifity (komplementarity)")
16
Strigence hybridizace
teplota ▲ koncentrace soli ▼ Denaturační činidla (formamid) za snížené teploty udržují vysokou strigenci
 za snížené teploty udržují vysokou strigenci.")
17
Princip Southern a Northern blotu
Hybridizační sondy jsou značené: chemiluminiscenčně BIOTIN detekce AVIDIN-HRP, DIGOXYGENIN fluorescenčně FLUORESCEIN, RHODAMIN radioaktivně dAT32P, 3H Příprava sond: 5´ a 3´koncové značení – nízká citlivost pomocí RANDOM HEXAMERů a DNA polymerasy (Klenowův fragment) PCR nick translace – DNasa I a DNA polymerasa in vitro transkripce (RNA próby) FISH fluorescent in situ hybridization
 PCR. nick translace – DNasa I a DNA polymerasa. in vitro transkripce (RNA próby) FISH fluorescent in situ hybridization.")
18
příklad nukleotidu značeného digoxygeninem

19
Příprava hybridizačních sond
značení pomocí náhodných hexameru značení pomocí nick translation (posun jednořetězcového zlomu)
")
20
RESTRIKČNÍ ENDONUKLEASY (restrikční enzymy II třídy)
rozpoznávají specifické sekvence dsDNA a štípou jí ve stejném místě rozpoznávací místo je 4-8 bp dlouhé štípou dlouhé fragmenty dsDNA na kratší štípou vždy znovu opakovatelným způsobem pocházejí především z bakterií ochrana před cizorodou DNA především bakteriofágovou modifikačně-restrikční enzymový systém (RE + specifická modifikační metylasa) Přes 120 rozpoznávacích míst dodnes popsaných více než 800 RE
 Přes 120 rozpoznávacích míst. dodnes popsaných více než 800 RE.")
21
RESTRIKČNÍ MíSTA MAJÍ BOD SYMETRIE
RE mohou na DNA vytvářet TUPÉ nebo LEPIVÉ konce restrikční sekvence jsou většinou symetrické. nukleotidy vytvářejí PALINDROM Palindrom: sekvence se čte stejně v obou směrech jak ve 5’ 3’ tak ve směru 5’ 3’ na komplementárním řetězci. EcoRI TUPÉ KONCE – nespecifické spojení LEPiVÉ KONCE – specifické spojení

22
Existují RE se stejnou restrikční sekvencí lišící se pouze typem vzniklých konců (5´- či 3´- lepivé) KpnI vs. Asp718 - izoschizomery Lepivé konce: každý DNA fragment (jakéhokoli původu) tvoří štěpením stejnou RE dvě jednovláknové identické sekvence (čtené ve směru 5’ - 3’). Kompatibilní konce: BamHI GˇGATCC BglII AˇGATCT
 tvoří štěpením stejnou RE dvě jednovláknové identické sekvence (čtené ve směru 5’ - 3’). Kompatibilní konce: BamHI GˇGATCC BglII AˇGATCT.")
23
dvě odlišné DNA mající stejné rozpoznávací místo pro danou RE se mohou vzájemně spojit v rekombinantní DNA molekulu rekombinantní DNA molekula: je nová kombinace nukleotidů v DNA která se nevyskytuje volně v přírodě

24
Počet nukleotidů v restrikčním místě dané RE determinuje průměrnou délku fragmentů DNA získaných naštípaním danou RE Pravděpodobnost že dané restrikční místo bude nalezeno v genomu: 44 = 256 bp 46 = 4096 bp 48 = bp GTAC průměrná vzdálenost mezi restrikčními sekvencemi v DNA = 4n n = # bází v restrikčním místě Působení RE je velmi citlivé na iontovou sílu roztoku a charakter přítomných solí. Pokud nejsou zachovány specifické podmínky pro danou RE ta může mít tzv. star aktivitu STAR AKTIVITA – nespecifické náhodné štěpení

25
T4 DNA ligasa Mung Bean nukleasa
enzym izolovaný z T4 bakteriofága infikujícího E.coli, katalyzuje tvorbu fosfodiesterové vazby mezi 5´fosfátovou skupinou jednoho a 3´OH skupinou sousedního nukleotidu. reakce vyžaduje energii jednoho ATP používá se ke spojování lepivých i tupých konců dvou restrikčních fragmentů DNA, připojování různých adaptorových sekvencí k DNA a cirkulizaci DNA Reakce se provádí většinou za snížené teploty (15-16°C) aby se snížila kinetická energie molekul a zabránilo se tak nekomplementárnímu spárování lepivých konců. zápor: nízká aktivita enzymu – dlouhý reakční čas. Další enzymy používané v molekulární biologii: Alkalická fosfatasa Mung Bean nukleasa
 aby se snížila kinetická energie molekul a zabránilo se tak nekomplementárnímu spárování lepivých konců. zápor: nízká aktivita enzymu – dlouhý reakční čas. Další enzymy používané v molekulární biologii: Alkalická fosfatasa. Mung Bean nukleasa.")
26
KLONOVÁNÍ Molekulární klonování:
multi-krokový proces který vytvoří kolekci definovaných fragmentů dané DNA pomocí restrikčních endonukleas spojení vybraných DNA fragmentů (většinou jednoho genu) se speciálním nosičem – vektor pomocí T4 DNA ligasy přenos vzniklého konstruktu do živých buněk (často baktérie) mnohonásobné namnožení vložených DNA fragmentů replikaci v živé buňce – DNA KLONY Buněčné klonování: vytváření geneticky identických buněk (organismů) v živé přírodě přirozené: kolonie baktérii řízkování rostlin jednovaječná dvojčata umělé ►
 se speciálním nosičem – vektor pomocí T4 DNA ligasy. přenos vzniklého konstruktu do živých buněk (často baktérie) mnohonásobné namnožení vložených DNA fragmentů replikaci v živé buňce – DNA KLONY. Buněčné klonování: vytváření geneticky identických buněk (organismů) v živé přírodě přirozené: kolonie baktérii. řízkování rostlin. jednovaječná dvojčata. umělé ►")
27
multiple cloning site (polylinker)
KLONOVACÍ VEKTORY Bakteriální a kvasinkové – PLASMIDY přirozené součástí bakteriálních buněk nesoucí vlastní genetickou informaci ve formě dsDNA (rezistence k antibiotikům, metabolismus nezvyklých substrátů…) 1-100kb replikují se nezávisle na baktérii vlastní plasmidové vektory vznikly úpravou přirozených „high copy“ plasmidy kopií v buńce pBR (Bolivar a Rodriguez) 4.36 kb menší než je přirozeně se vyskytující v E.coli ori vlastní počátek replikace rezistence k Amp a Tc unikátní restrikční místa - kapacita pro inzertovou DNA 1-5 kb MCS multiple cloning site (polylinker) ori
 1-100kb. replikují se nezávisle na baktérii. vlastní plasmidové vektory vznikly úpravou přirozených. „high copy plasmidy kopií v buńce pBR (Bolivar a Rodriguez) 4.36 kb menší než je přirozeně se vyskytující v E.coli. ori vlastní počátek replikace. rezistence k Amp a Tc. unikátní restrikční místa. - kapacita pro inzertovou DNA 1-5 kb. MCS. multiple cloning site (polylinker) ori.")
28
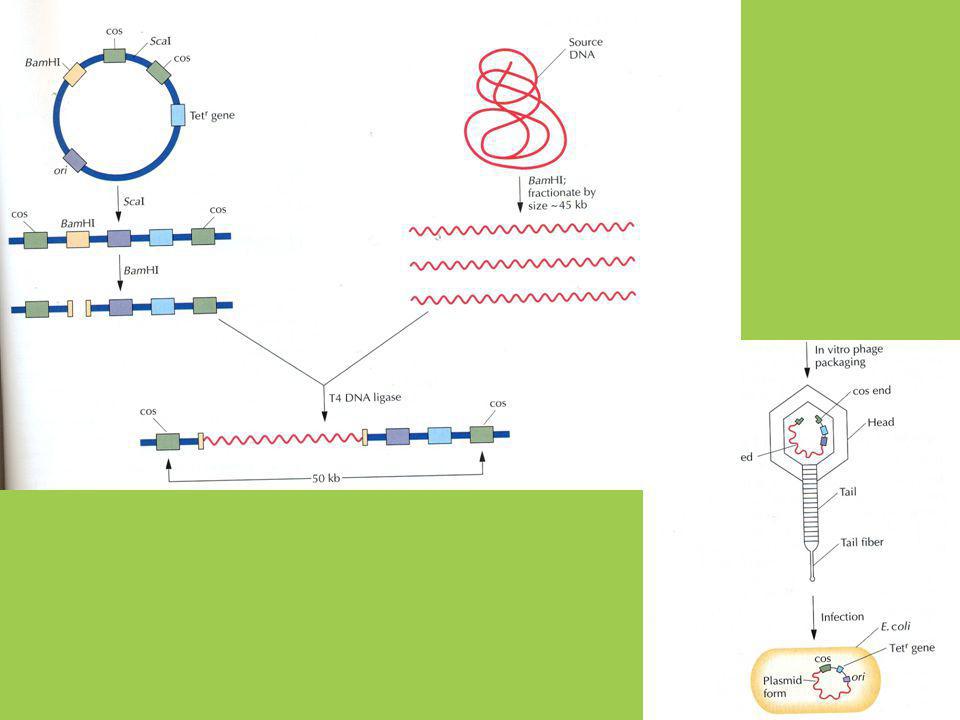
KLONOVACÍ VEKTORY Virové - BAKTERIOFÁGY
bakteriální viry s dvouřetězcovou lineární DNA klonovací kapacita 2-25kb inzertu mnohonásobné množení v závislosti na hostitelské bakterii cos BAKTERIOFÁG lambda kb Střední část genomu není důležitá pro lytický růst a lze ji nahradit inzertovou DNA snadná manipulace díky cos místům (lepivé konce) na koncích λ DNA, které umožňují její cirkulaci Zacirkuluje se pouze bakteriofág, který má vzdálenost mezi cos místy 37-52kb tvorba cDNA a genomových knihoven cos
 na koncích λ DNA, které umožňují její cirkulaci. Zacirkuluje se pouze bakteriofág, který má vzdálenost mezi cos místy 37-52kb. tvorba cDNA a genomových knihoven. cos.")
29
KLONOVACÍ VEKTORY C) bakteriálně-virové - KOSMIDY
odvozené z plasmidu do kterého byly naklonovány cos místa bakteriofága λ Inzertová DNA je naklonována do lineárního kosmidu a sbalena in vitro virovým mechanismem a infikována do bakterie v bakterii je pak cirkulována a replikuje se dál jako plasmid malá velikost cca 5kb – není potřeba infekčních lytických λ proteinů, pouze selekční marker k antibiotiku, velikost inzertové DNA se může pohybovat až k 47kb D) bakteriálně-virové - FASMIDY (phagemid) odvozené od bakteriofága M13 s malou ssDNA (6.4kb), který se po infekci do bakterie replikuje jako kruhovitá dsDNA (plasmid, snadná manipulace, restrikce) infekční fágové částice však opět obsahují pouze ssDNA (vhodný zdroj pro izolaci ssDNA např. pro hybridizaci) do infekčních částic mohou být sbaleny částice až dvakrát větší než je fágová DNA
 bakteriálně-virové - FASMIDY (phagemid) odvozené od bakteriofága M13 s malou ssDNA (6.4kb), který se po infekci do bakterie replikuje jako kruhovitá dsDNA (plasmid, snadná manipulace, restrikce) infekční fágové částice však opět obsahují pouze ssDNA (vhodný zdroj pro izolaci ssDNA např. pro hybridizaci) do infekčních částic mohou být sbaleny částice až dvakrát větší než je fágová DNA.")
31
KLONOVACÍ VEKTORY E) kvasinkové - YAC umělé kvasinkové chromosomy (yeast artificial chromosomes) KYVADLOVÉ VEKTORY umožňující existenci a replikaci ve dvou rozdílných organismech: pYES bakterie – kvasinka - počátek replikace z přirozeného kvasinkového vektoru 2μ Ti-plazmid bakterie – rostlina viz přednáška transgenoze rostlin patří mezi kyvadlové vektory (shuttle vector) v bakterii jako kruhová dsDNA (plasmid) vedle sekvencí bakteriálního plasmidu obsahují části kvasinkového chromozomu (centromera, počátek replikace,dvě telomery) linearizací se chromozom aktivuje (oddělení telomer od sebe) obrovská klonovací kapacita až 2000kb nestabilita a složitá manipulace (protoplasty)
 v bakterii jako kruhová dsDNA (plasmid) vedle sekvencí bakteriálního plasmidu obsahují části kvasinkového chromozomu (centromera, počátek replikace,dvě telomery) linearizací se chromozom aktivuje (oddělení telomer od sebe) obrovská klonovací kapacita až 2000kb. nestabilita a složitá manipulace (protoplasty)")
32
KLONOVACÍ VEKTORY F) bakteriální - BAC umělé bakteriální chromosomy (bacterial artificial chromosomes) Odvozené od E.coli F-plasmidu (fertility plasmid), „low copy“ 1-2 kopie na buńku VÝHODY oproti YAC: stabilita (cirkulární) snadná manipulace potlačení rekombinace Vhodné fragmenty genomické DNA pro ligaci jsou získány pulsní gelovou elektroforesou VELKÝ VÝZNAM PŔI MANIPULACI S CELÝMI GENOMY INVITROGEN
, „low copy 1-2 kopie na buńku. VÝHODY oproti YAC: stabilita (cirkulární) snadná manipulace. potlačení rekombinace. Vhodné fragmenty genomické DNA pro ligaci. jsou získány pulsní gelovou elektroforesou. VELKÝ VÝZNAM PŔI MANIPULACI S CELÝMI GENOMY. INVITROGEN.")
33
Obecný princip molekulárního klonování
klonovací vector + DNA obsahující požadovaný gen (izolace) restrikce ligace transformace selekce pomocí selekčních markerů reprodukce a syntéza cílového proteinu izolace
 restrikce. ligace. transformace. selekce pomocí selekčních markerů. reprodukce a syntéza cílového proteinu. izolace.")
34
Využití molekulárního klonování
uchovávání, zmnožení a manipulace s genetickou informací (cDNA a genomové knihovny) produkce proteinů pro různorodé využití vnášení nových či pozměněných genů do organismů (GMO, genová terapie) farmaceutika zemědělství základní výzkum průmysl medicína
 produkce proteinů pro různorodé využití. vnášení nových či pozměněných genů do organismů (GMO, genová terapie) farmaceutika. zemědělství. základní výzkum. průmysl. medicína.")
35
SELEKČNÍ MARKERY geny kódující degradaci ANTIBIOTIKA
geny nesené na klonovacím vektoru, exprimované současně s transgenem, na jejichž fenotypovém projevu lze transformované buňky snadno selektovat geny kódující degradaci ANTIBIOTIKA INHIBITORY SYNTÉZY BAKTERIÁLNÍ BUNĚČNÉ STĚNY AMPICILIN gen Amp koduje -laktamasu která štěpí -laktámový kruh penicilinových antibiotik CEFOTAXIM INHIBITORY SYNTÉZY PROTEINů CHLORAMFENIKOL, KANAMYCIN (vážou se na ribozomální podjednotky) gen Cml kóduje chloramfenikol acyltransferasu gen Kan kóduje aminoglykosid fosfotransferasu AMPICILIN
 gen Cml kóduje chloramfenikol acyltransferasu. gen Kan kóduje aminoglykosid fosfotransferasu. AMPICILIN.")
36
SELEKČNÍ MARKERY geny kódující degradaci ANTIBIOTIKA
INHIBITORY SYNTÉZY PROTEINů TETRACYKLIN (zabraňuje vazbě aminoacyl tRNA k ribozomu) gen Tet kóduje membránový protein, který aktivně transportuje TC ven z buňky HYGROMYCIN B (interferuje s velkou ribozomální podjednotkou a zabraňuje elongaci syntetizovaného peptidu) gen způsobující rezistenci hgh kóduje fosforylasu inaktivující toto antibiotikum. PůSOBÍ I NA EUKARYOTICKÉ BUŇKY!!! (selekce transgenních rostlin) INHIBITORY TRANSKRIPCE A REPLIKACE ACTINOMYCIN D (inhibuje transkripci tím, že se váže na specifické úseky DNA) RIFAMPICIN (inhibuje prokaryotní RNA polymerasu, ale i chloroplastovou a mitochondriální, proto je toxicky i pro člověka) ZEOCIN – glykopeptid izolovaný ze Streptomyces, který štěpí po vstupu do buňky jakoukoliv DNA. Rezistentní gen Sh ble kóduje inhibiční protein (13.6 kDa) tohoto glykopeptidu. koncentrace účinné pro selekci: Baktérie g/mL Kvasinky g/mL Savčí buňky g/mL ZEOCIN
 gen Tet kóduje membránový protein, který aktivně transportuje TC ven z buňky. HYGROMYCIN B (interferuje s velkou ribozomální podjednotkou a zabraňuje elongaci syntetizovaného peptidu) gen způsobující rezistenci hgh kóduje fosforylasu inaktivující toto antibiotikum. PůSOBÍ I NA EUKARYOTICKÉ BUŇKY!!! (selekce transgenních rostlin) INHIBITORY TRANSKRIPCE A REPLIKACE. ACTINOMYCIN D (inhibuje transkripci tím, že se váže na specifické úseky DNA) RIFAMPICIN (inhibuje prokaryotní RNA polymerasu, ale i chloroplastovou a mitochondriální, proto je toxicky i pro člověka) ZEOCIN – glykopeptid izolovaný ze Streptomyces, který štěpí po vstupu do buňky jakoukoliv DNA. Rezistentní gen Sh ble kóduje inhibiční protein. (13.6 kDa) tohoto glykopeptidu. koncentrace účinné pro selekci: Baktérie g/mL. Kvasinky g/mL. Savčí buňky g/mL. ZEOCIN.")
37
kyvadlový vektor pro transformaci kvasinek
SELEKČNÍ MARKERY 2) geny kódující enzymy narušující NUTRIČNÍ NEDOSTATEČNOST (ztráta autotrofie) - používají se především pro selekci transgenních kvasinek Gen URA3 kóduje enzym orotidin 5´-fosfát dekarboxylasu, enzym metabolické dráhy uracilu LYS2 kóduje aminoadipát reduktasu (bisyntéza lyzinu) ADE2 (C1-tetrahydrofolát syntasa), LEU2 (- izopropylmalát dehydrogenasa), TRP1 (fosforibozylanthranilát izomerasa) Princip selekce: pro transformaci daného plasmidu se použijí specifické kmeny kvasinek deficientní pro daný gen, selekce se pak provádí na minimálním médiu (bez živin). Příklad: kmen Saccharomyces cerevisiae INVSc1 genotyp: MATa his31 leu2 trp1-289 ura3-52 /MAT his31 leu2 trp1-289 ura3-52 fenotyp: His-, Leu-, Trp-, Ura- tzn. že tento kmen je autotrofní na histidin, leucin, tryptofan a uracil a na médiu bez těchto živin neporoste kyvadlový vektor pro transformaci kvasinek
 geny kódující enzymy narušující NUTRIČNÍ NEDOSTATEČNOST (ztráta autotrofie) - používají se především pro selekci transgenních kvasinek. Gen URA3 kóduje enzym orotidin 5´-fosfát dekarboxylasu, enzym metabolické dráhy uracilu. LYS2 kóduje aminoadipát reduktasu (bisyntéza lyzinu) ADE2 (C1-tetrahydrofolát syntasa), LEU2 (- izopropylmalát dehydrogenasa), TRP1 (fosforibozylanthranilát izomerasa) Princip selekce: pro transformaci daného plasmidu se použijí. specifické kmeny kvasinek deficientní pro daný gen, selekce. se pak provádí na minimálním médiu (bez živin). Příklad: kmen Saccharomyces cerevisiae INVSc1. genotyp: MATa his31 leu2 trp1-289 ura3-52. /MAT his31 leu2 trp1-289 ura3-52. fenotyp: His-, Leu-, Trp-, Ura- tzn. že tento kmen je autotrofní na histidin, leucin, tryptofan a uracil a na médiu bez těchto živin neporoste. kyvadlový vektor pro transformaci kvasinek ")
38
SELEKČNÍ MARKERY 3) blue/white screening test O OH HOCH2 HO N H Br
pomocný test pro selekci transformovaných bakterií či bakteriofágu klonovací vektor nese gen lacZ+ který kóduje enzym -galaktosidasu (štěpí laktosu na galaktosu a glukosu) MCS je umístěn uvnitř tohoto genu, a po vložení inzertu se tento gen stává neaktivní X-GAL O OH HOCH2 HO N H Br Cl galaktosa modré barvivo Ampicilin (zabije netransformované bakterie) X-GAL (5-Bromo-4-chloro-3-indolyl b-D-galactopyranoside) umělý substrát IPTG (isopropyl-1-thio--D-galaktopyranoside) induktor exprese lacZ genu
 MCS je umístěn uvnitř tohoto genu, a po vložení inzertu se tento gen stává neaktivní. X-GAL. O. OH. HOCH2. HO. N. H. Br. Cl. galaktosa. modré barvivo. Ampicilin (zabije netransformované bakterie) X-GAL (5-Bromo-4-chloro-3-indolyl b-D-galactopyranoside) umělý substrát. IPTG (isopropyl-1-thio--D-galaktopyranoside) induktor exprese lacZ genu.")
39
lytický a lysogenní cyklus virů

40
struktura lambda bakteriofága

41
inzert (genomová knihovna)
SELEKČNÍ MARKERY 4) lysogenní selekce bakteriofágových vektoru LAMBDA FIX II VEKTOR ( bakteriofágový vektor) - selekce se provádí na speciálním kmeni E.coli (lysogenní pro P2 bakteriofága, tzn. že hostitelská buňka je již napadená jiným bakteriofágem) Pokud má vektor genotyp red/gam+ na daném kmeni neroste je tzv. Sensitivní k P2 interferenci (Spi+ fenotyp) red a gam geny se nacházejí ve střední části (stuffer) a jsou nahrazeny inzertem. Bakteriofág s inzertem je tudiž Spi- a může být vyselektován na P2 lysogenním kmeni red/gam+ kb red/gam- příliš malý nesbalí se do kapsidy inzert (genomová knihovna)
 lysogenní selekce bakteriofágových vektoru. LAMBDA FIX II VEKTOR ( bakteriofágový vektor) - selekce se provádí na speciálním kmeni E.coli (lysogenní pro P2 bakteriofága, tzn. že hostitelská buňka je již napadená jiným bakteriofágem) Pokud má vektor genotyp red/gam+ na daném kmeni neroste je tzv. Sensitivní k P2 interferenci (Spi+ fenotyp) red a gam geny se nacházejí ve střední části (stuffer) a jsou nahrazeny inzertem. Bakteriofág s inzertem je tudiž Spi- a může být vyselektován na P2 lysogenním kmeni. red/gam+ 12 kb red/gam- příliš malý. nesbalí se. do kapsidy. inzert (genomová knihovna)")
Podobné prezentace

>")





>")
>")










