Stáhnout prezentaci
1
Imunitní odpověď založená na protilátkách
J. Ochotná

2
Protilátková reakce vyvolaná:
Antigeny nezávislými na T lymfocytech Vyvolávají převážně tvorbu IgM protilátek Jedná se zvláště o bakteriální polysacharidy, lipopolysacharidy a polymerní formy proteinů Antigeny závislými na T lymfocytech Reakce na tyto Ag probíhá ve dvou fázích – primární a sekundární Tvorba paměťových buněk a vznik vysokoafinních protilátek

3
T-nezávislá a T-závislá imunitní odpověď

4
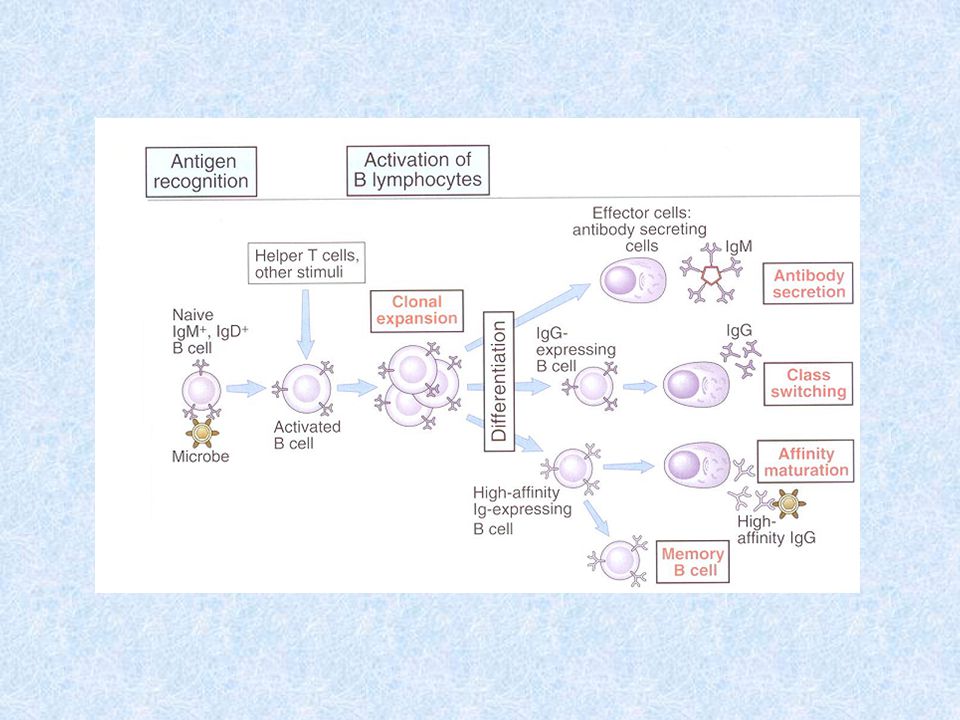
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Primární fáze protilátkové reakce Probíhá v sekundárních lymfatických orgánech Stimulace B lymfocytu vazbou Ag na BCR Pohlcení Ag APC a jeho prezentace prostřednictvím MHC gp II prekurzorům TH buněk → vznik klonu antigenně specifických TH2 buněk, které poskytují pomoc příslušným B lymfocytům, což vede k jejich proliferaci, diferenciaci na plazmatické bb (produkují Ab) a na paměťové bb
 a na paměťové bb.")
5
Primární fáze protilátkové reakce
Plazmatické bb jsou rozneseny oběhovým systémem do organismu (zvláště kostní dřeně) Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům
 Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy. Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům.")
6
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Sekundární fáze protilátkové reakce – po rozpoznání Ag na FDC Vznikne-li dostatečné množství imunokomplexů na FDC a jsou-li rozpoznány B lymfocyty Germinální reakce - pod vlivem signálů od FDC (Ag) a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí
 a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek. Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí.")
7
V sekundární fázi imunitní reakce vznikají protilátky s vyšší afinitou k Ag a s jinými efektorovými vlastnostmi závislými na izotypu, vznikají také paměťové bb pro další setkání s Ag Protilátky v organismu po primární infekci přetrvávají po dlouhou dobu Kontakt mezi CD40 (B lymfocyt) a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.
 a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.")
9
Protilátková odpověď Primární odpověď
následuje po prvním kontaktu s antigenem, kdy ještě nejsou přítomny paměťové buňky tvoří se převážně nízkoafinní IgM, postupně se tvoří ostatní izotypy s vyšší afinitou

10
Sekundární odpověď následuje po opakovaných setkáních se stejným antigenem, jsou již přítomny paměťové buňky hladina IgM protilátek je nižší, rychleji se tvoří ostatní izotypy, protilátky mají vyšší afinitu k antigenu, protože jsou aktivovány paměťové buňky

11
Dynamika tvorby protilátek při primární a sekundární odpovědi

12
Imunoglobuliny

13
Struktura imunoglobulinů
2 těžké (H) řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l Typy H řetězců – m, d, g (g1-4), a (a1,a2), e
 řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec. H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l. Typy H řetězců – m, d, g (g1-4), a (a1,a2), e.")
14
Struktura imunoglobulinů

15
Variabilní domény L a H řetězce tvoří vazebné místo pro Ag
Pantová oblast = místo, kde jsou těžké řetězce spojeny cystinovými můstky Fc části imunoglobulinů jsou silně glykosylovány, váží se na Fc receptory J řetězec Sekreční komponenta

16
Funkce imunoglobulinů
Neutralizace Ag Aglutinace Ag Aktivace komplementu Opsonizace Aktivace mastocytů prostřednictvím IgE ADCC

17
Třídy imunoglobulínů a jejich funkce
Rozlišujeme podle konstantní části H řetězce IgM, IgD, IgG ( IgG1 – IgG4), IgA (IgA1, IgA2), IgE IgM - jako monomer tvoří BCR sekretovaný ve formě pentamerů (10 vazebných míst) první izotyp, který se tvoří po setkání s Ag neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR v séru se nachází ve velmi malé koncentraci - (koncentrace 0,1 g/l; biol. poločas 3 dny)
, IgA (IgA1, IgA2), IgE. IgM - jako monomer tvoří BCR - sekretovaný ve formě pentamerů (10 vazebných míst) - první izotyp, který se tvoří po setkání s Ag - neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů - (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR - v séru se nachází ve velmi malé koncentraci. - (koncentrace 0,1 g/l; biol. poločas 3 dny)")
18
IgG - jednotlivé izotypy IgG1-IgG4 se liší schopností aktivovat komplement a vázat se na Fc receptory fagocytů (opsonizace) - funkce: neutralizace, opsonizace, aktivace komplementu, ADCC - přestupuje placentou - tvořen při sekundární imunitní odpovědi (koncentrace 8 – 18 g/l; biol. poločas 21 dnů)
")
19
IgA slizniční IgA - ochrana sliznic, neutralizace, opsonizace, neaktivuje komplement dimér se sekreční komponentou sliny, slzy, mateřské mléko sérový IgA – monomer, dimer či trimer (koncentrace 0,9 – 3,5 g/l; biol. poločas 6 dnů) IgE - uplatňuje se při obraně proti mnohobuněčným parazitům je hlavní příčinou alergických reakcí - (koncentrace 3x10-4 g/l; biol. poločas 2 dny)
 IgE - uplatňuje se při obraně proti mnohobuněčným parazitům - je hlavní příčinou alergických reakcí. - (koncentrace 3x10-4 g/l; biol. poločas 2 dny)")
20
Imunoglobuliny

21
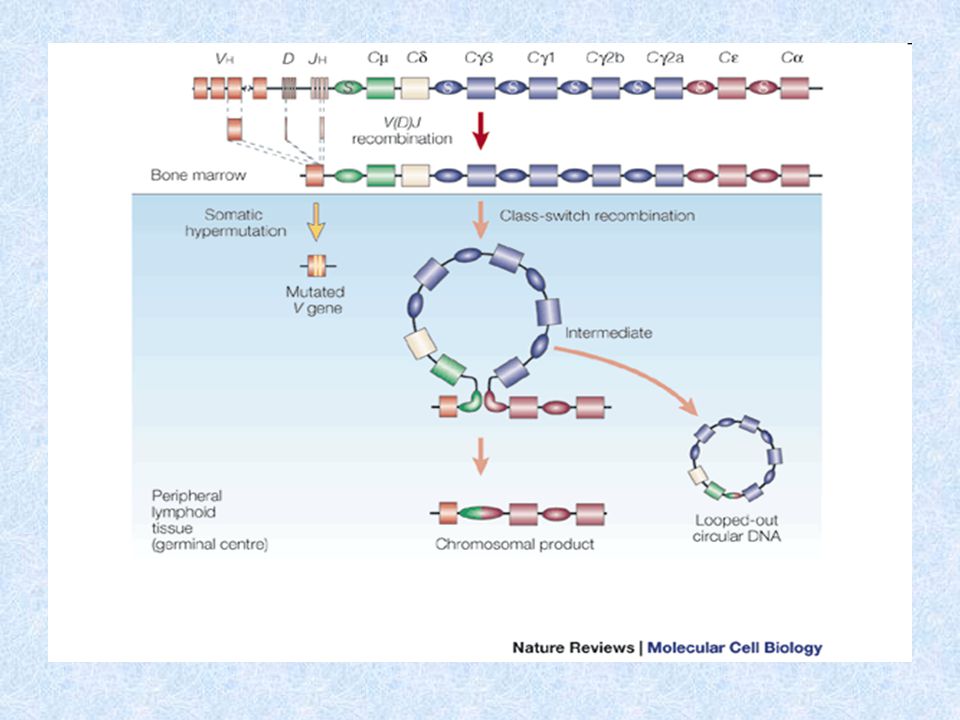
Genové segmenty pro H řetězce - chromozóm 14
V (variabilita) - několik set D (diversita) - asi 50 J (joining) – 9 C konstantní části H řetězce Genové segmenty pro L řetězce - k chromozóm 2 - l chromozóm 22 V (variabilita) J (joining) C konstantní části L řetězce Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk
 - několik set. D (diversita) - asi 50. J (joining) – 9. C konstantní části H řetězce. Genové segmenty pro L řetězce - k chromozóm 2. - l chromozóm 22. V (variabilita) J (joining) C konstantní části L řetězce. Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů. Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk.")
22
Genetický základ tvorby imunoglobulinů

23
Přeskupování genů kódujících H řetězce
D-J přeskupení - vyštěpení úseku IgH mezi některým D a J segmentem (probíhá na obou chromozómech) V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.
 V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.")
24
Přeskupování genů kódujících L řetězce
Nejprve se přeskupují geny kódující L řetězec k, dochází k vyštěpování úseků mezi některým V a J segmentem (souběžně na obou chromozómech), pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.
, pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.")
25
Izotypový přesmyk (class switch)
Dochází k němu během terminální diferenciace B lymfocytu po aktivaci Ag na povrchu FDC (nezbytný kostimulační signál přes CD40) Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů a vyštěpují úseky genů switch sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce
 Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů a vyštěpují úseky genů. switch sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň. Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce.")
27
Izotypový přesmyk (class switch) - pokračování
Cytokiny regulují k jakému izotypovému přesmyku dojde: IL-4 stimuluje přesmyk na IgE a IgG4, (u myší IgG1) TGFb stimuluje přesmyk na IgG2 a IgA
 TGFb stimuluje přesmyk na IgG2 a IgA.")
28
Antiidiotypové protilátky
IDIOTOP = každá determinanta variabilní části protilátky, může být součástí paratopu, či může ležet mimo paratop IDIOTYP = soubor idiotopů (souhrn identických vazebných struktur pro Ag na protilátkách stejné specifity) Idiotypické struktury protilátek 1. generace jsou rozpoznávány některými B lymfocyty jako antigeny a mohou se proti nim tvořit tzv. antiidiotypové protilátky (protilátky 2. generace; některá vazebná místa mohou připomínat Ag, který vyvolal tvorbu protilátek 1. generace) Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi
 Idiotypické struktury protilátek 1. generace jsou rozpoznávány některými B lymfocyty jako antigeny a mohou se proti nim tvořit tzv. antiidiotypové protilátky (protilátky 2. generace; některá vazebná místa mohou připomínat Ag, který vyvolal tvorbu protilátek 1. generace) Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi.")
29
Slizniční a kožní imunitní systém

30
Funkce a struktura slizničního a kožního imunitního systému sliznice a kůže jsou ve stálém kontaktu s vnějším prostředím, proto je zde soustředěno asi 80% imunokompetentních buněk kůže – bariéra proti mechanickému, fyzikálnímu a chemickému poškození a proti průniku mikroorganismů, u člověka představuje povrch asi 1,5 m2 slizniční imunitní systém – brání průniku patogenních mikroorganismů a rozvoji sebepoškozujících zánětlivých imunitních reakcí proti patogenům a neškodným antigenům z vnějšího prostředí, sliznice mají plochu asi 400 m2

31
Přirozené neimunitní obranné mechanismy: Mechanické obranné mechanismy – neporušený povrch sliznic a epitelií; řasinkový epitel pomáhá odstraňovat antigeny zachycené v hlenu, k jejich odstranění napomáhá i kašel, kýchání, zvracení a průjem; longitudinální tok vzduchu v dýchacích cestách a tekutiny v močových cestách Chemické obranné mechanismy – mastné kyseliny na povrchu kůže; lysozym obsažený ve slinách, slzách a na sliznicích narušuje bakteriální stěnu; kyselina chlorovodíková v žaludeční šťávě ničí choroboplodné zárodky; antimikrobiální peptidy produkované epitelovými buňkami narušují membránu patogenních organismů (jsou součástí přirozené imunity), kyselé pH moče brání růstu patogenů
, kyselé pH moče brání růstu patogenů")
32
Přirozené neimunitní obranné mechanismy:
Mikrobiální obranné mechanismy – přirozená mikroflóra soutěží s patogenními mikroorganismy o povrchové receptory, o nutriční zdroje a také napomáhá produkci bakteriostatických a baktericidních látek, které brání množení patogenních mikroorganismů Další fyziologické faktory, které ovlivňují obranyschopnost organismu – tělesná teplota 37oC není vhodná pro růst mnoha patogenů; tenze kyslíku ve tkáních brání růstu anaerobních patogenů; obranyschopnost organismu je také ovlivněna věkem, stresem a hormonálními změnami

33
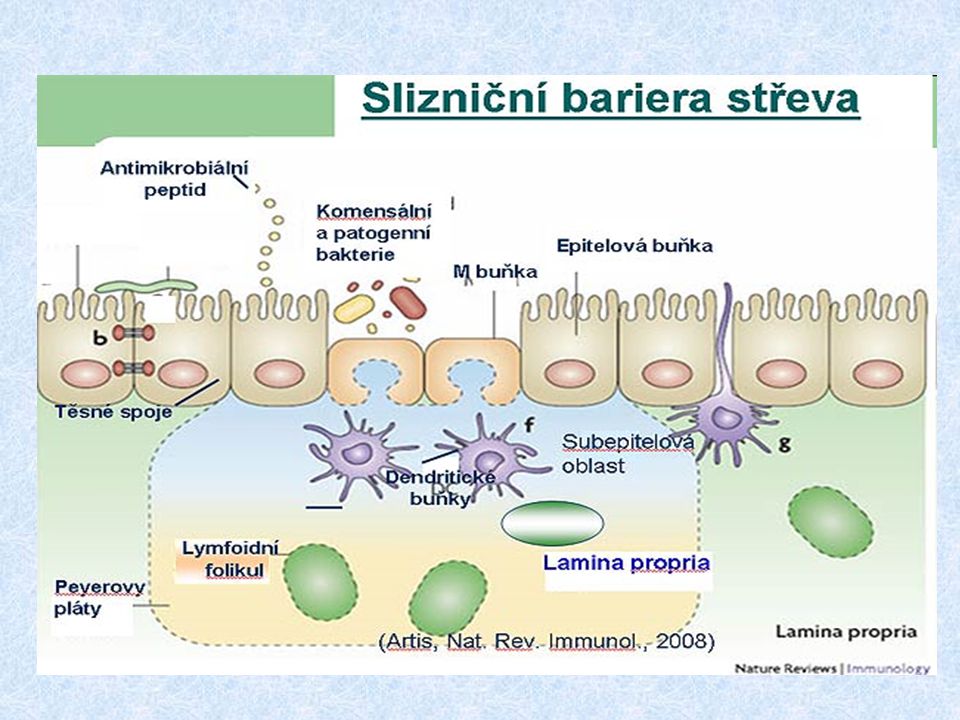
Struktura slizničního imunitního systému MALT (mucous associated lymphoid tissue) BALT (bronchus associated lymphoid tissue) GALT (gut associated lymphoid tissue) NALT (nasal-associated lymphoid tissue) o-MALT (organisovaný) – je tvořen lymfoidními folikuly pod sliznicí; patrové a nosní mandle, apendix, Peyerovy plaky d-MALT (difúzní) – je tvořen leukocyty difuzně rozprostřenými v lamina propria (T a B lymfocyty, makrofágy, neutrofily, eozinofily a žírné bb.)
 BALT (bronchus associated lymphoid tissue) GALT (gut associated lymphoid tissue) NALT (nasal-associated lymphoid tissue) o-MALT (organisovaný) – je tvořen lymfoidními folikuly pod sliznicí; patrové a nosní mandle, apendix, Peyerovy plaky d-MALT (difúzní) – je tvořen leukocyty difuzně rozprostřenými v lamina propria (T a B lymfocyty, makrofágy, neutrofily, eozinofily a žírné bb.)")
34
Humorální mechanismy slizničního imunitního systému s IgA
Humorální mechanismy slizničního imunitního systému s IgA * sekreční imunoglobulin A * nejvýznamější slizniční imunoglobulin; přítomný i v mateřském mléce * transcytoza – IgA je přes epitel transportován pomocí transportního Fc receptoru (poly-Ig-receptor), na luminální straně je IgA odštěpen i s částí receptoru tzv. sekreční komponentou, která chrání Ig před střevními proteázami * neutralizace antigenů na sliznicích, neaktivuje komplement, váže se na Fc receptory fagocytů * imunokomplexy s IgA mohou být zachyceny v Peyerových placích a mohou indukovat imunitní odpověď
, na luminální straně je IgA odštěpen i s částí receptoru tzv. sekreční komponentou, která chrání Ig před střevními proteázami * neutralizace antigenů na sliznicích, neaktivuje komplement, váže se na Fc receptory fagocytů * imunokomplexy s IgA mohou být zachyceny v Peyerových placích a mohou indukovat imunitní odpověď.")
35
s IgM. sekreční imunoglobulin M
s IgM * sekreční imunoglobulin M * uplatňuje se u novorozenců a selektivního deficitu IgA * více náchylný k degradaci střevními proteázami * neutralizace antigenů na slizničních površích IgG * dostává se na sliznice difúzí * uplatňuje se zvláště v dolních dýchacích cestách

36
Indukce slizniční imunitní reakce Orální tolerance
Indukce slizniční imunitní reakce Orální tolerance * většina antigenů podaných perorálně vyvolá supresi specifické imunity (rozhodující je velikost antigenní částice) * Tr lymfocyty (regulační) – produkce IL-10 Indukce slizniční imunitní reakce M-buňky - specializované enterocyty, které zajišťují transport Ag (endocytují Ag z okolí) jsou v těsném kontaktu s lymfocyty a APC Slizniční imunizace vede ke stimulaci TH2 a TH3 lymfocytů a produkci IgA
 * Tr lymfocyty (regulační) – produkce IL-10 Indukce slizniční imunitní reakce M-buňky - specializované enterocyty, které zajišťují transport Ag (endocytují Ag z okolí) - jsou v těsném kontaktu s lymfocyty a APC Slizniční imunizace vede ke stimulaci TH2 a TH3 lymfocytů a produkci IgA.")
38
Imunologický význam kojení mateřské mléko obsahuje:
Imunologický význam kojení mateřské mléko obsahuje: * sIgA, IgG (neutralizace infekčních mikroorganismů, jejich produktů a potencionálních alergenů – než se plně rozvine slizniční imunitní systém novorozence) * CD 59 (protektin) – ochrana bb. před působením komplementu * lysozym, laktoferin, složky komplementu, cytokiny včetně interferonů * imunokompetentní bb.
 * CD 59 (protektin) – ochrana bb. před působením komplementu * lysozym, laktoferin, složky komplementu, cytokiny včetně interferonů * imunokompetentní bb.")
39
Kožní imunitní systém epidermis
Kožní imunitní systém epidermis * keratinocyty - sekrece cytokinů (IL-1, 6, TNF, IL-10, TGFb) exprese MHCgpII → mohou sloužit jako APC * Langerhansovy bb. – kožní dendritické bb. (APC) * rozptýlené intraepiteliální lymfocyty * melanocyty dermis * fibroblasty - produkce kolagenu odstraňování apoptotických bb. * mastocyty * T lymfocyty (malé množství) * cévy, vlasové folikuly, potní a mazové žlázy
 - exprese MHCgpII → mohou sloužit jako APC * Langerhansovy bb. – kožní dendritické bb. (APC) * rozptýlené intraepiteliální lymfocyty * melanocyty dermis * fibroblasty - produkce kolagenu - odstraňování apoptotických bb. * mastocyty * T lymfocyty (malé množství) * cévy, vlasové folikuly, potní a mazové žlázy.")
40
Imunitní mechanismy zánětu (lokální a systémová reakce)
")
41
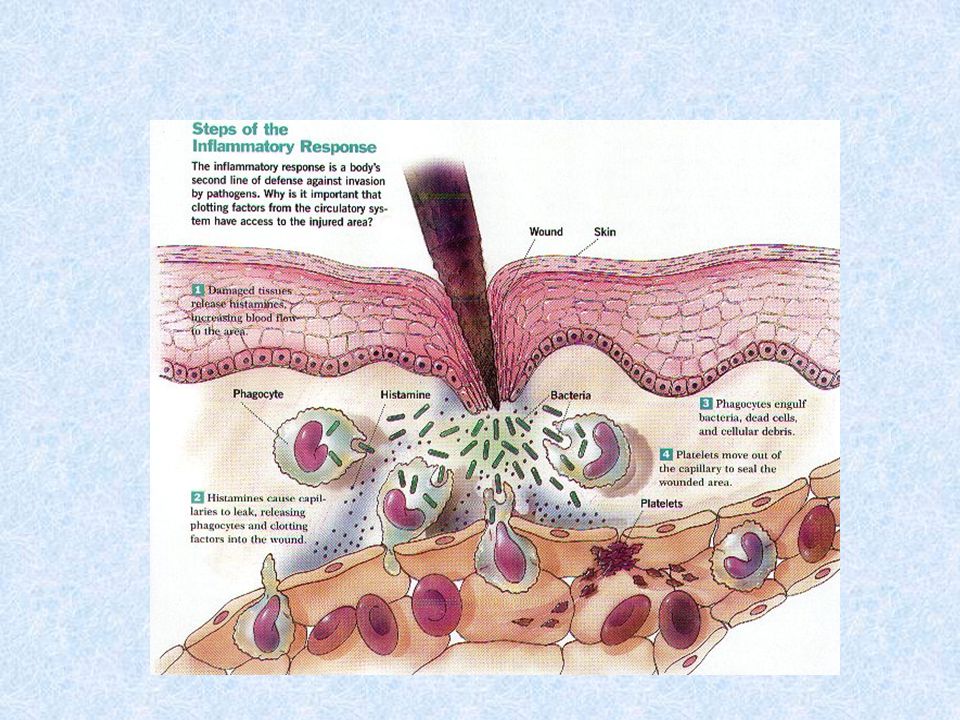
Zánět Je souhrn fyziologických reakcí na porušení integrity organismu, které vedou k ochraně proti infikování poškozeného místa, k lokalizaci poškození a jeho zhojení. První signály k rozvoji zánětlivých reakcí pocházejí od mastocytů, fagocytů a od látek uvolněných z poškozených buněk a součástí mezibuněčné hmoty.

42
Zánět - fyziologická obranná reakce - obvykle odezní bez následků, poškozená tkáň se zcela zhojí) patologický zánět – chronický, alergický, autoimunitní Odpověď organismu - lokální systémová
 - patologický zánět – chronický, alergický, autoimunitní Odpověď organismu - lokální - systémová")
43
Lokální odpověď organismu na zánět Projevy - bolest (dolor), teplo (calor), zčervenání (rubor), otok (tumor) a ztrátou funkce (funkcio laesa)
, teplo (calor), zčervenání (rubor), otok (tumor) a ztrátou funkce (funkcio laesa)")
44
Lokální zánět * zvýšení permeability cév ( vazoaktivní aminy, složky komplementu C3a, C5a, leukotrieny; otok v místě zánětu ) * zvýšení exprese adhezivních molekul na endoteliích * aktivace koagulačního, fibrinolytického, komplementového a kininového systému * ovlivnění místních nervových zakončení ( prostaglandiny, bolest ) * změny regulace teploty ( IL- 1, IL-6, TNF, prostaglandiny )
 * zvýšení exprese adhezivních molekul na endoteliích * aktivace koagulačního, fibrinolytického, komplementového a kininového systému * ovlivnění místních nervových zakončení ( prostaglandiny, bolest ) * změny regulace teploty ( IL- 1, IL-6, TNF, prostaglandiny )")
46
Systémová odpověď na zánět
Systémová odpověď na zánět * je závislá na rozsahu poškození a délce trvání lokálního zánětu * horečka (prozánětlivé cytokiny , IL-1, IL-6, TNF, IFN… stimulují hypotalamové centrum termoregulace) * mobilizace tkáňového metabolismu * indukce exprese Hsp (heat-shock-proteins; fungují jako chaperony) * produkce proteinů akutní fáze (CRP, SAP, C4, C5; opsonizace a aktivace komplementu)
 * mobilizace tkáňového metabolismu * indukce exprese Hsp (heat-shock-proteins; fungují jako chaperony) * produkce proteinů akutní fáze (CRP, SAP, C4, C5; opsonizace a aktivace komplementu)")
47
* zvýšená jaterní syntéza některých sérových transportních proteinů (ceruloplasmin, transferin) * zvýšená syntéza proteázových inhibitorů (makroglobulín) * leukocytóza Septický šok – při masivním průniku mikroorganismů do krevního oběhu Anafylaktický šok – při degranulaci bazofilů po kontaktu s alergenem
 * zvýšená syntéza proteázových inhibitorů (makroglobulín) * leukocytóza Septický šok – při masivním průniku mikroorganismů do krevního oběhu Anafylaktický šok – při degranulaci bazofilů po kontaktu s alergenem")
48
Reparace poškozené tkáně eliminace poškozených buněk fagocyty aktivace fibroplastických mechanismů aktivace angiogeneze regenerace a remodelace tkání

49
Fyziologické imunitní regulační mechanismy

50
Regulace antigenem Vyvolání a vyhasnutí imunitní odpovědi
Afinitní maturace B lymfocytů Udržení imunologické paměti Antigenní kompetice Prahová hustota komplexu MHC gp II-Ag na APC

51
Regulace protilátkami
Protilátky kompetují s BCR o antigen (negativní regulátor stimulace B lymfocytů) Imunokomplexy s IgG se váží na B lymfocyt na BCR a FcgR, důsledkem je blokování aktivace B lymfocytů Zatím je nejasný význam regulace pomocí idiotypové sítě
 Imunokomplexy s IgG se váží na B lymfocyt na BCR a FcgR, důsledkem je blokování aktivace B lymfocytů. Zatím je nejasný význam regulace pomocí idiotypové sítě.")
52
Regulace cytokiny a mezibuněčným kontaktem
Interakce APC - T lymfocyt Interakce TH1 – makrofág Interakce TH2 – B lymfocyt Vzájemná regulace aktivit TH1 versus TH2 Vývoj subpopulací leukocytů Negativní regulace efektorových lymfocytů: CTLA-4 inhibiční receptor T lymfocytů, váže ligandy CD80 a CD86 Inhibiční receptory NK buněk Sebedestrukční interakce apoptotického receptoru Fas s ligandem FasL na povrchu aktivovaných T lymfocytů

53
Suprese zprostředkovaná T lymfocyty
Vzájemná negativní interakce TH1 a TH2 zprostředkovaná cytokiny (TH2 lymfocyty produkují IL-4 a IL-10, které potlačují imunitní reakce založené na TH1 buňkách) Klonální eliminace či anergizace T lymfocytů po styku s antigenem na povrchu jiných buněk, než APC (chybí kostimulační signály) Regulační T lymfocyty pomáhají udržet toleranci k autoantigenům
 Klonální eliminace či anergizace T lymfocytů po styku s antigenem na povrchu jiných buněk, než APC (chybí kostimulační signály) Regulační T lymfocyty pomáhají udržet toleranci k autoantigenům.")
54
Faktory ovlivňující výsledek imunitní odpovědi
Tentýž antigen může navodit aktivní imunitní odpověď nebo stav aktivní tolerance, výsledek odpovědi závisí na mnoha faktorech: Stavu imunitního systému Vlastnostech antigenu Dávce antigenu Způsobu podání

55
Cytokiny ( tkáňové hormony )
")
56
Cytokiny Regulační proteiny a glykoproteiny produkované leukocyty i jinými buňkami Základní regulátory imunitního systému Uplatňují se i mimo imunitní systém (angiogeneze, regenerace tkání, kancerogeneze, ovlivnění řady mozkových funkcí,embryoníální vývoj…) Cytokiny - sekretované - membránové (zajištěno lokální působení; CD 80, CD86, CD40L, FasL..)
 Cytokiny - sekretované. - membránové (zajištěno lokální působení; CD 80, CD86, CD40L, FasL..)")
57
Pleiotropní účinek Působí v kaskádě Cytokinová síť Cytokinový systém je redundantní Působení cytokinů- autokrinní - parakrinní - endokrinní Jsou označovány jako interleukiny (vyjímka: TNF, lymfotoxin, TGF, interferony, CSF a růstové faktory)
")
58
B lymfocyt komunikuje s makrofágem a T lymfocytem pomocí cytokinů

59
Rozdělení cytokinů podle funkce
Prozánětlivé cytokiny (IL-1 a , IL-6, 8, 12, 18, TNF) Protizánětlivé cytokiny (IL-1Ra, IL-4, IL-10, TGF ) Cytokiny s aktivitou růstových faktorů hemopoetických bb. ( IL-2, 3, 4, 5, 6, CSF, SCF, LIF, EPO ) Cytokiny uplatňující se v humorální imunitě TH2 ( IL-4, 5, 9, 13 ) Cytokiny uplatňující se v buněčně zprostředkované imunitě TH1 ( IL- 2, 12, IFN, GM-CSF, lymfotoxin ) Cytokiny s antivirovým účinkem ( IFN-, IFN-, IFN- )
 Protizánětlivé cytokiny (IL-1Ra, IL-4, IL-10, TGF ) Cytokiny s aktivitou růstových faktorů hemopoetických bb. ( IL-2, 3, 4, 5, 6, CSF, SCF, LIF, EPO ) Cytokiny uplatňující se v humorální imunitě TH2 ( IL-4, 5, 9, 13 ) Cytokiny uplatňující se v buněčně zprostředkované imunitě TH1 ( IL- 2, 12, IFN, GM-CSF, lymfotoxin ) Cytokiny s antivirovým účinkem ( IFN-, IFN-, IFN- )")
60
Přehled nejvýznamnějších cytokinů
Zdroj Funkce IL-1 MF, N Kostimulace T lymf., indukce TNF a IL-8, pyrogen IL-2 Th1 Růstový faktor pro T lymf. IL-4 Th2, bazofily Diferenciace Th2, stimulace B lymf., indukce izotypového přesmyku na IgE a IgG4, inhibice Th1 IL-5 Th2, eozinofily Stimulace B lymf., růstový faktor pro eozinofily IL-6 Th2, MF, N Stimulace T a B lymf., stimulace produkce Ig, indukce syntézy proteinů akutní fáze, pyrogen IL-8 MF a ostatní bb. Chemotaxe a aktivace granulocytů (především neutrofilů) IL-10 Th2,M, Treg Inhibice Th1 a MF, indukce diferenciace B lymf. na plazmocyty IL-12 MF, DC, B Diferenciace Th1, stimulace NK TNF M, MF, NK Indukce lokálního zánětu, aktivace endotelu, , indukce apoptózy TGFb T, MF, trombocyty Protizánětlivý účinek (tlumení proliferace lymfocytů, tlumení produkce Ig, tlumení cidní aktivity MF), stimulace fibroblastů a osteoblastů, zesílení produkce mezibuněčné hmoty IFNa L, M, MF Inhibice virové replikace IFNb Fibroblasty, epitelie IFNg Th1, NK Aktivace MF, stimulace exprese MHC gp., inhibice Th2 MF – makrofágy; M – monocyty; N – neutrofily; DC – dendritické bb.; NK – přirození zabíječi; L – lymfocyty; B – B lymf.; T – T lymf.
 IL-10. Th2,M, Treg. Inhibice Th1 a MF, indukce diferenciace B lymf. na plazmocyty. IL-12. MF, DC, B. Diferenciace Th1, stimulace NK. TNF. M, MF, NK. Indukce lokálního zánětu, aktivace endotelu, , indukce apoptózy. TGFb. T, MF, trombocyty. Protizánětlivý účinek (tlumení proliferace lymfocytů, tlumení produkce Ig, tlumení cidní aktivity MF), stimulace fibroblastů a osteoblastů, zesílení produkce mezibuněčné hmoty. IFNa. L, M, MF. Inhibice virové replikace. IFNb. Fibroblasty, epitelie. IFNg. Th1, NK. Aktivace MF, stimulace exprese MHC gp., inhibice Th2. MF – makrofágy; M – monocyty; N – neutrofily; DC – dendritické bb.; NK – přirození zabíječi; L – lymfocyty; B – B lymf.; T – T lymf.")
61
Receptory cytokinů Jsou složeny ze 2 či 3 podjednotek
Jedna váže cytokin, další asociovány s cytoplazmatickými signalizačními molekulami (protein-kinázami) Signalizační podjednotka bývá sdílena několika různými cytokinovými receptory – tzv receptorové rodiny Signalizace přes tyto receptory může vést k proliferaci, diferenciaci, aktivaci efektorových mechanismů či zablokování buněčného cyklu a indukci apoptózy
 Signalizační podjednotka bývá sdílena několika různými cytokinovými receptory – tzv receptorové rodiny. Signalizace přes tyto receptory může vést k proliferaci, diferenciaci, aktivaci efektorových mechanismů či zablokování buněčného cyklu a indukci apoptózy.")













 Mgr. Martin Šmíd.>")



 Nespecifická : Fagocytóza granulocytů,monocytů.>")



>")