Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
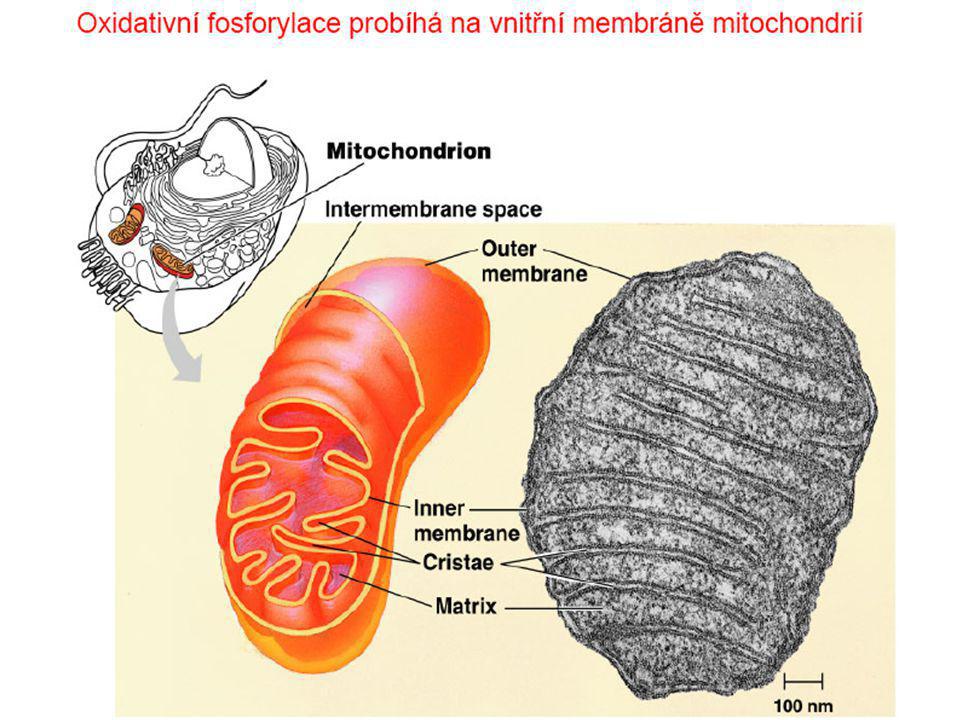
Dýchací řetězec Viz též přednášky prof. Kodíčka (další materiály na webu)
")
2
Odbourávání glukosy (včetně substrátových fosforylací)
C6H12O6 + 6O2 -->6 CO2 + 6H2O + 38 ATP

20
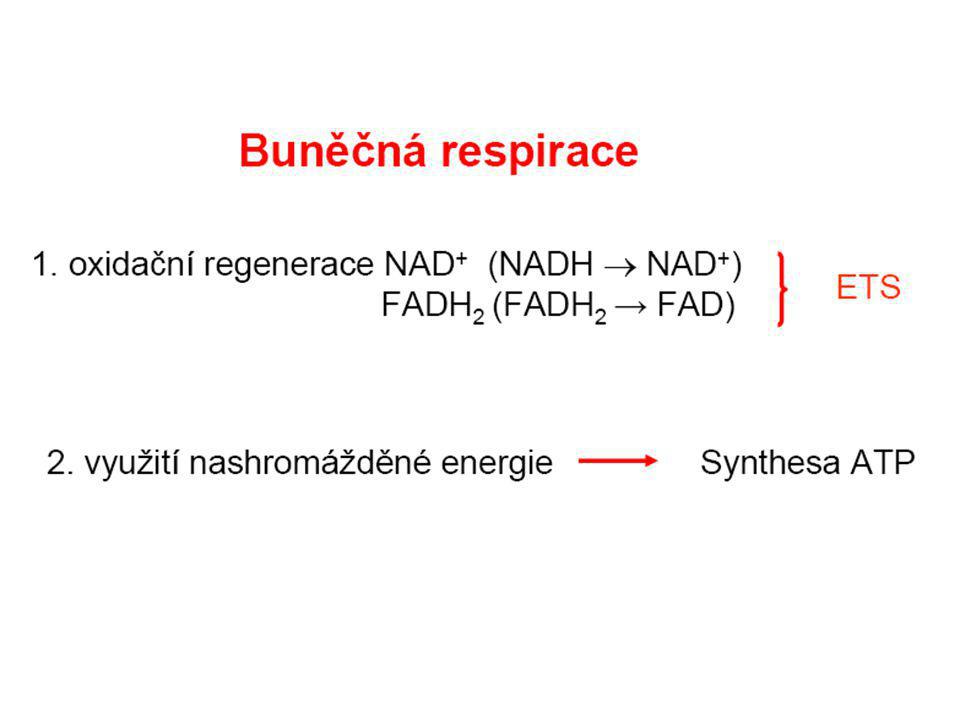
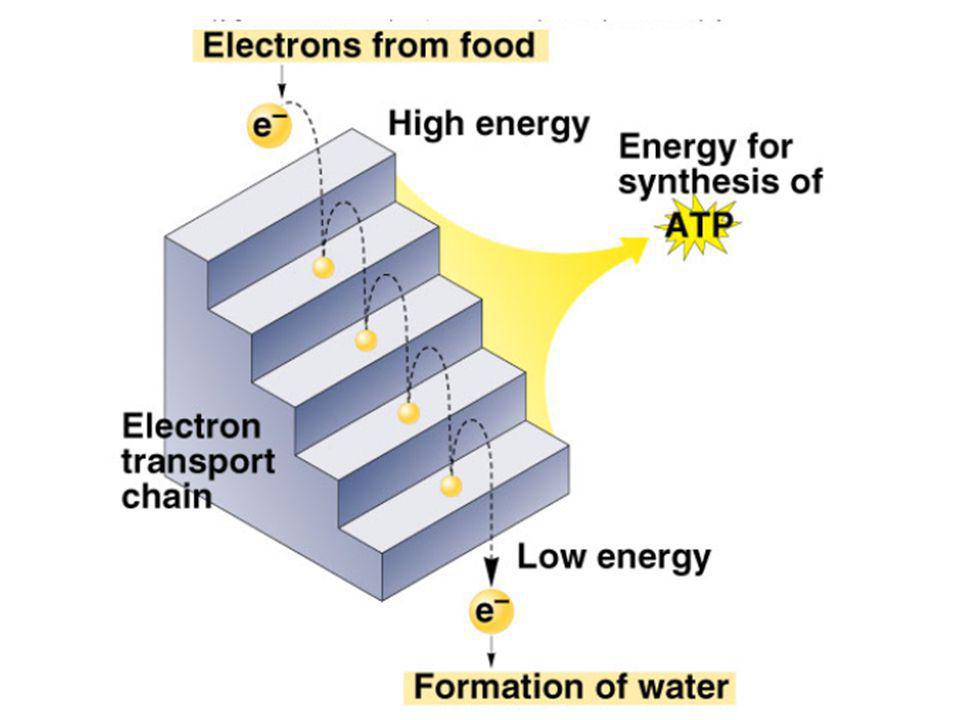
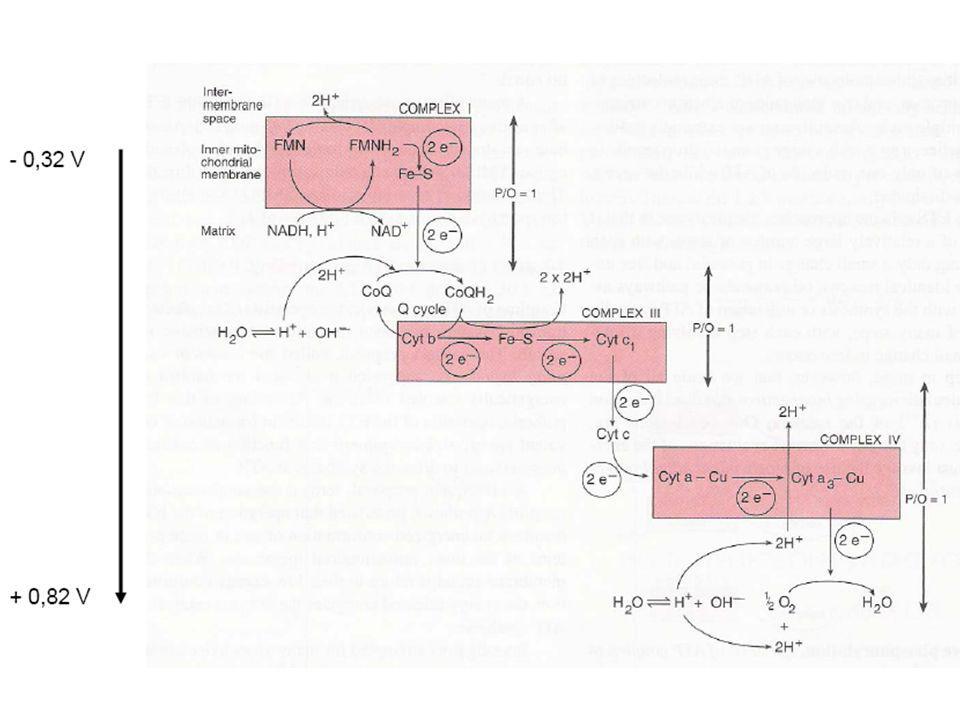
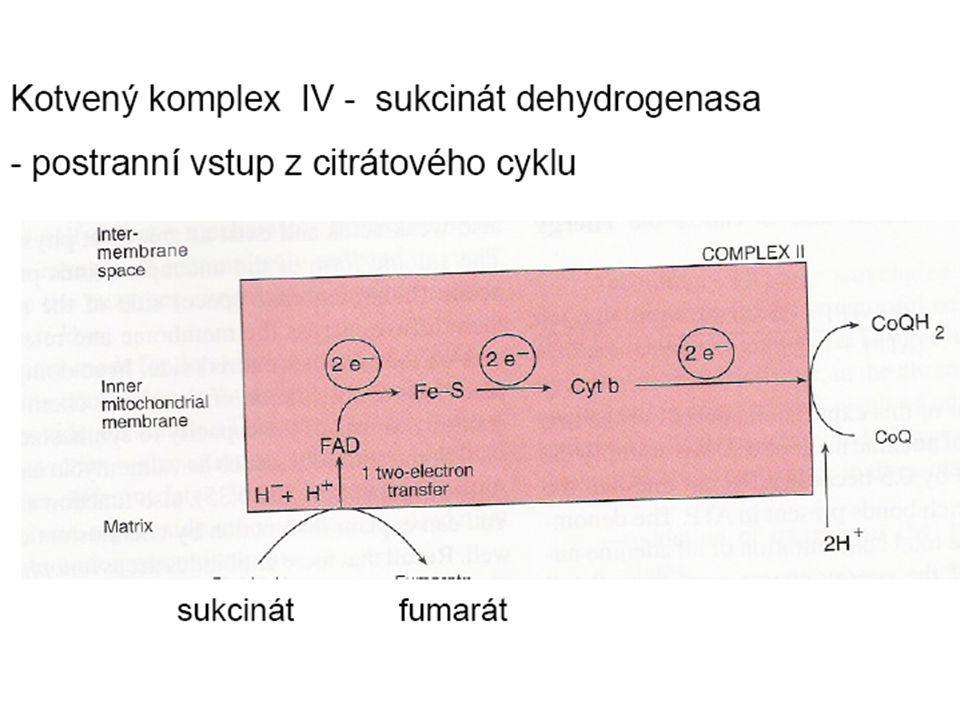
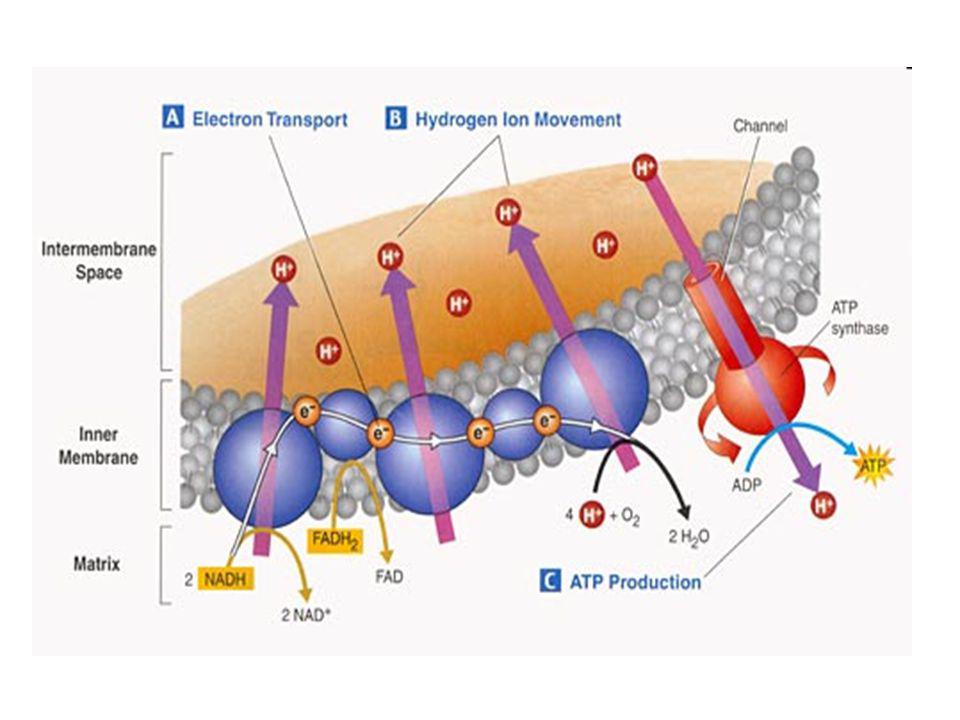
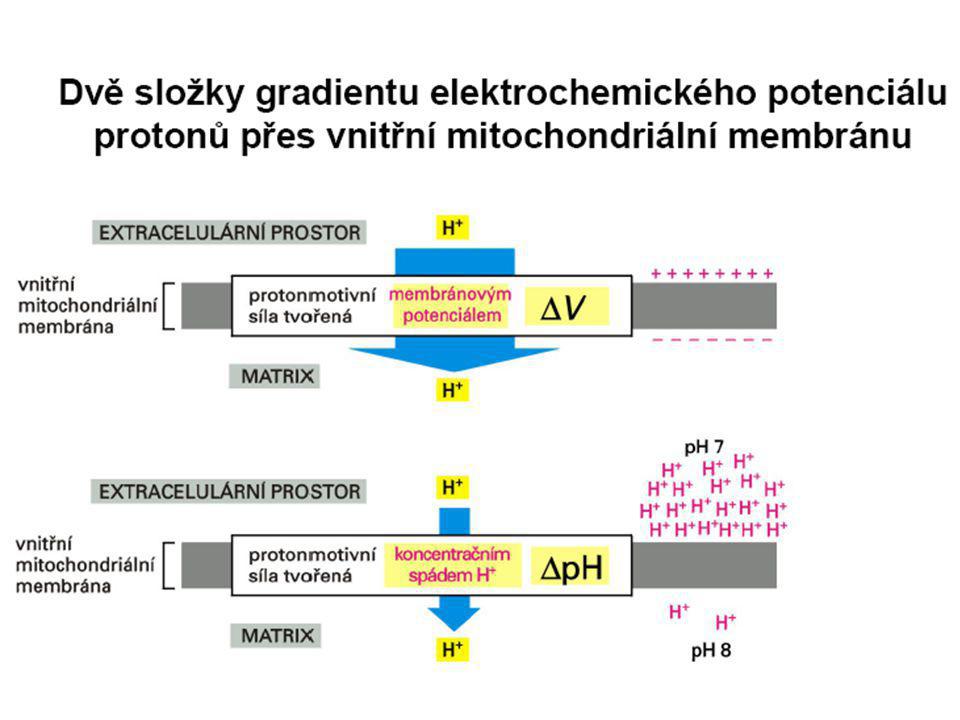
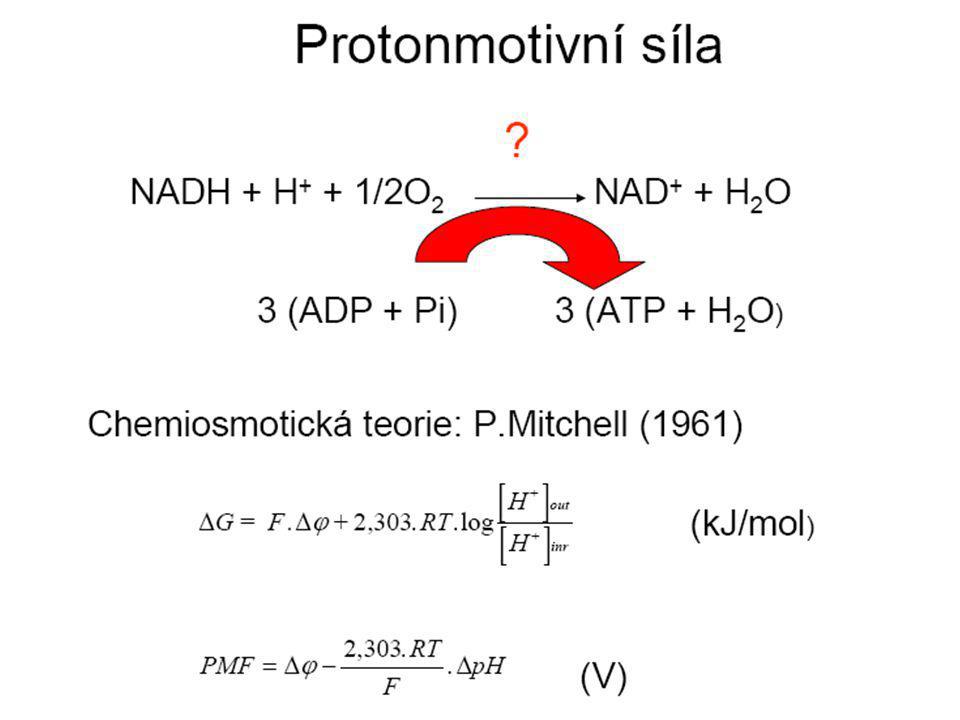
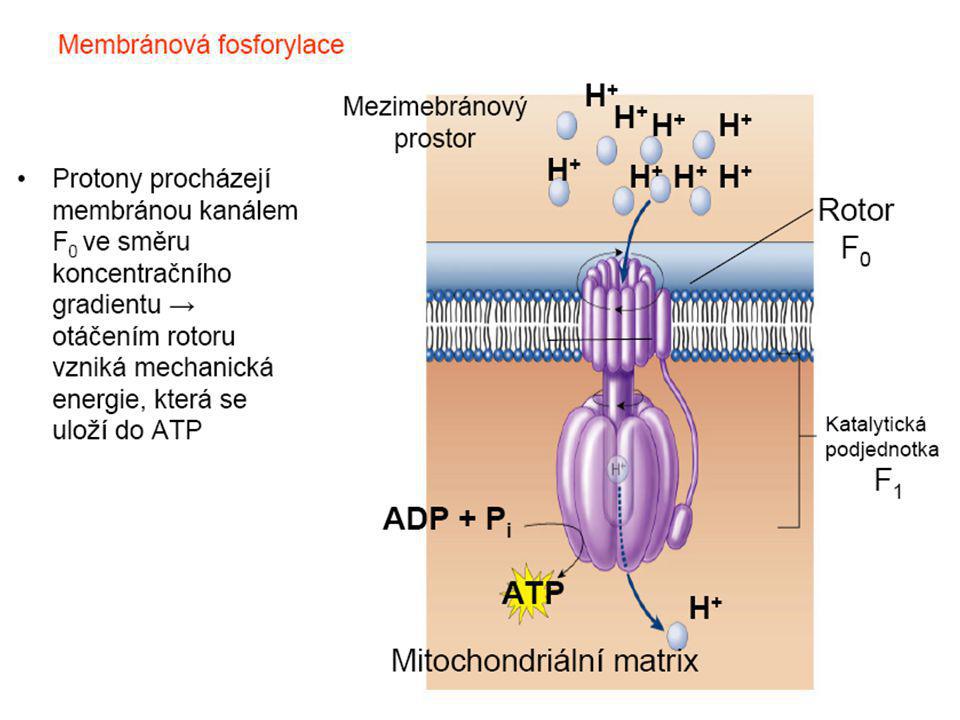
Dýchací řetězec - souhrn
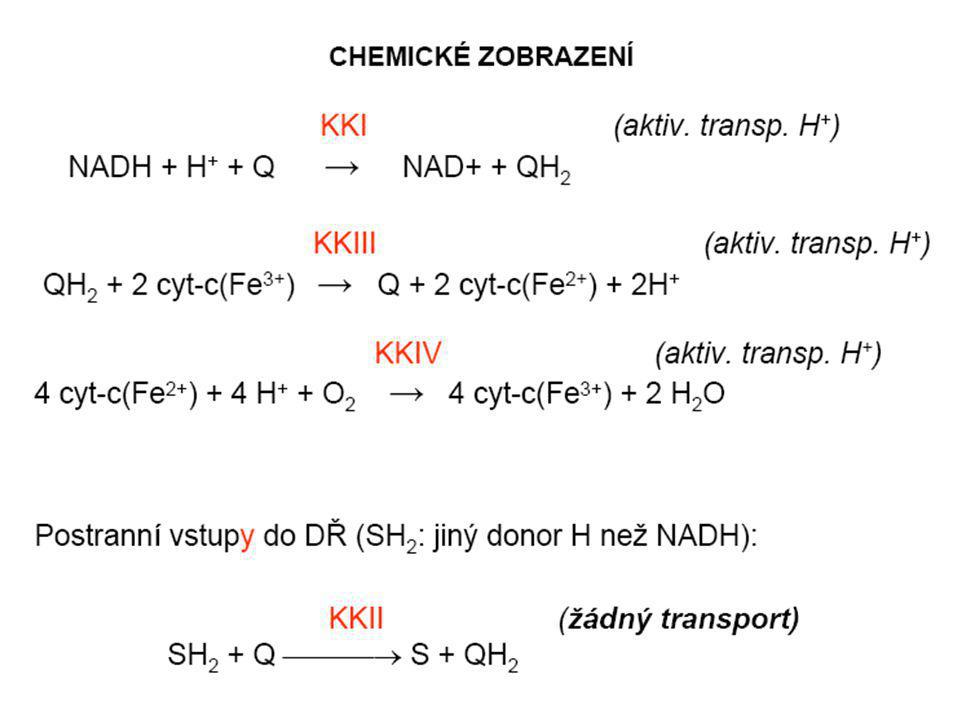
systém oxidoreduktas a mobilních přenašečů atomů vodíku nebo elektronů, který v biologické membráně zajišťuje reoxidaci redukovaných kofaktorů (viz aktivní vodík) a na úkor této exergonické reakce aktivně transportuje vodíkové protony, čímž generuje proton-motivní sílu. U eukaryot je tento systém lokalisován ve vnitřní membráně mitochondrie, u prokaryot v cytoplazmatické membráně. U savců je dýchací řetězec tvořen enzymy ubichinon-reduktasa (kotvený komplex I), který katalysuje přenos vodíku z NADH na ubichinon (viz koenzym Q) (NADH + H+ + Q → NAD+ + QH2), cytochrom-c-reduktasa (kotvený komplex III) (QH2 + 2 cytochrom-c(Fe3+) → Q + 2 cytochrom-c(Fe2+) + 2 H+) a cytochrom-c-oxidasa (kotvený komplex IV) (4 cytochrom-c(Fe2+) + O2 + 4 H+ → 4 cytochrom-c(Fe3+) + 2 H2O). Tyto tři kotvené komplexy (integrální membránové enzymy) zajišťují endergonický transport protonů z matrix mitochondrie přes vnitřní membránu (úhrnem přibližně 12 H+ na oxidaci jednoho NADH); kotvený komplex II je tzv. postranní vstup do dýchacího řetězce. Konečným akceptorem elektronů u aerobních organismů je kyslík (viz respiarace aerobní), u respirujících anaerobních chemoorganotrofů je to jiné oxidační činidlo přijímané z vnějšího prostředí (viz respirace anaerobní).
 a na úkor této exergonické reakce aktivně transportuje vodíkové protony, čímž generuje proton-motivní sílu. U eukaryot je tento systém lokalisován ve vnitřní membráně mitochondrie, u prokaryot v cytoplazmatické membráně. U savců je dýchací řetězec tvořen enzymy ubichinon-reduktasa (kotvený komplex I), který katalysuje přenos vodíku z NADH na ubichinon (viz koenzym Q) (NADH + H+ + Q → NAD+ + QH2), cytochrom-c-reduktasa (kotvený komplex III) (QH2 + 2 cytochrom-c(Fe3+) → Q + 2 cytochrom-c(Fe2+) + 2 H+) a cytochrom-c-oxidasa (kotvený komplex IV) (4 cytochrom-c(Fe2+) + O2 + 4 H+ → 4 cytochrom-c(Fe3+) + 2 H2O). Tyto tři kotvené komplexy (integrální membránové enzymy) zajišťují endergonický transport protonů z matrix mitochondrie přes vnitřní membránu (úhrnem přibližně 12 H+ na oxidaci jednoho NADH); kotvený komplex II je tzv. postranní vstup do dýchacího řetězce. Konečným akceptorem elektronů u aerobních organismů je kyslík (viz respiarace aerobní), u respirujících anaerobních chemoorganotrofů je to jiné oxidační činidlo přijímané z vnějšího prostředí (viz respirace anaerobní).")
21
Dýchací řetězec - souhrn

22
Fotosynthesa

23
Celkově zjednodušeně: 6 CO2 + 6 H2O → C6H12O6 + 6 O2

24
Fotosynthesa

26
Fotosynthesa CO2 + 4 [H] -CH(OH)- + H2O Potřebujeme:
- energii: ADP + Pi + h ATP + H2O redukční činidlo: NADP+ + H2D + h NADPH + H+ + D (D = donor)
![Fotosynthesa CO2 + 4 [H] -CH(OH)- + H2O Potřebujeme:](http://slideplayer.cz/slide/1987566/7/images/26/Fotosynthesa+CO2+%2B+4+%5BH%5D+%EF%82%AE+-CH%28OH%29-+%2B+H2O+Pot%C5%99ebujeme%3A.jpg "- energii: ADP + Pi + h ATP + H2O. redukční činidlo: NADP+ + H2D + h NADPH + H+ + D. (D = donor)")
28
Dva základní typy fotosynthesy
anoxygenní oxygenní ANOXYGENNÍ fotosynthesu uskutečňují gramnegativní bakterie (purpurové sirné a nesirné, zelené sirné bakterie) Donorem vodíku jsou jiné anorganické látky než voda (např. H2S) a žádným z vedlejších produktů není kyslík. Všechny fotosynthetické procesy probíhají za anaerobních podmínek. OXYGENNÍ fotosynthesu uskutečňují sinice a rostliny. K redukci CO2 je vždy využíván vodík uvolněný pri fotolyse vody a vedlejším produktem je kyslík. Oxygenní fotosynthesa je hlavním zdrojem kyslíku v atmosfére. V podstatě veškerý atmosférický kyslík pochází z činnosti fotosynthetisujících organismu.
 Donorem vodíku jsou jiné anorganické látky než voda (např. H2S) a žádným z vedlejších produktů není kyslík. Všechny fotosynthetické procesy probíhají za anaerobních podmínek. OXYGENNÍ fotosynthesu uskutečňují sinice a rostliny. K redukci CO2 je vždy využíván vodík uvolněný pri fotolyse vody a vedlejším produktem je kyslík. Oxygenní fotosynthesa je hlavním zdrojem kyslíku v atmosfére. V podstatě veškerý atmosférický kyslík pochází z činnosti fotosynthetisujících organismu.")
29
Fotosynthesa 6 CO2 + 6 H2O + 2,7 kJ C6H12O6 + 6O2
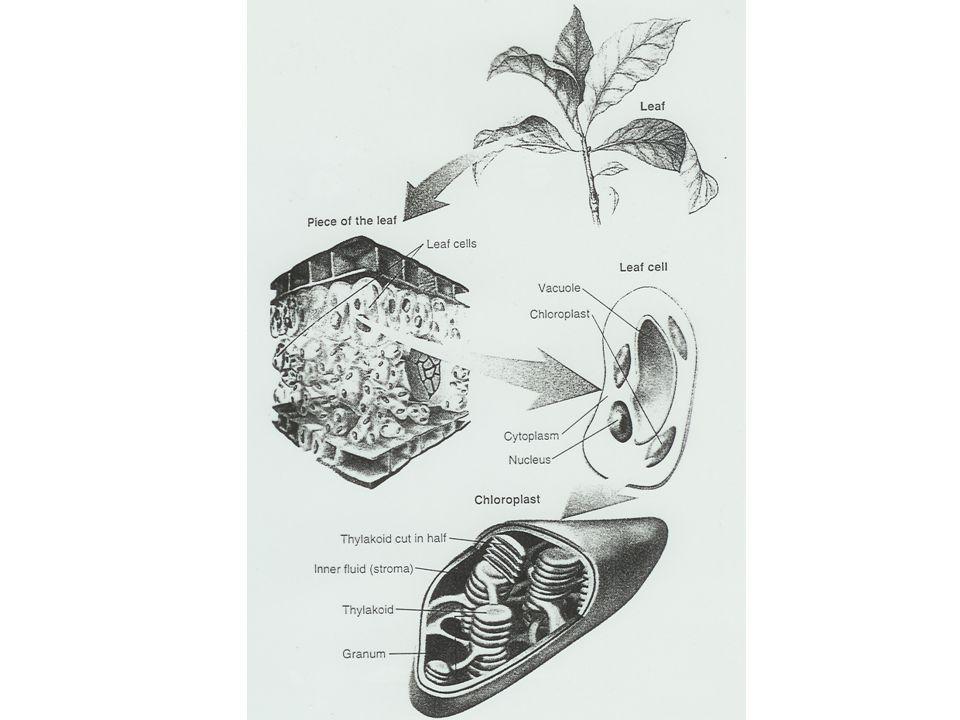
proces, kterým se v zelených rostlinách světelná energie mění na energii chemických vazeb; dvě fáze, tj. světelnou a temnostní CO2 je pomocí tohoto procesu asimilováno, a zabudováno do organických sloučenin, produktem jsou sacharidy (škrob, cukry) sumárně lze fotosyntézu vyjádřit rovnicí: 6 CO2 + 6 H2O + 2,7 kJ C6H12O6 + 6O2 vázána u vyšších rostlin na chloroplasty - cca v 1 buňce, 2 biomembrány; semiautochtonní; stroma a síť tylakoidů = trubičky a polštářky, orientované v grana, tj. granální x agranální thylakoidy tylakoidy - vnější stěna: bílkovinná struktura, vnitřní stěna: lipidická struktura; obsahují fotosyntecká barviva (vázané na tzv. fotosystémy), přenašeče elektronů a enzymy (1. světelná fáze fotosyntézy)
 sumárně lze fotosyntézu vyjádřit rovnicí: 6 CO2 + 6 H2O + 2,7 kJ C6H12O6 + 6O2. vázána u vyšších rostlin na chloroplasty - cca v 1 buňce, 2 biomembrány; semiautochtonní; stroma a síť tylakoidů = trubičky a polštářky, orientované v grana, tj. granální x agranální thylakoidy. tylakoidy - vnější stěna: bílkovinná struktura, vnitřní stěna: lipidická struktura; obsahují fotosyntecká barviva (vázané na tzv. fotosystémy), přenašeče elektronů a enzymy (1. světelná fáze fotosyntézy)")
30
Fotosynthesa Fotosynthesa probíhá ve dvou fázích: primární (světelné) a sekundární (temnostní), přičemž každá v jiné části chloroplastu. U eukaryot se reakce světelné fáze uskutečňují na membránách tylakoidu, fáze temnostní ve stromatu chloroplastu.
 a sekundární (temnostní), přičemž každá v jiné části chloroplastu. U eukaryot se reakce světelné fáze uskutečňují na membránách tylakoidu, fáze temnostní ve stromatu chloroplastu.")
33
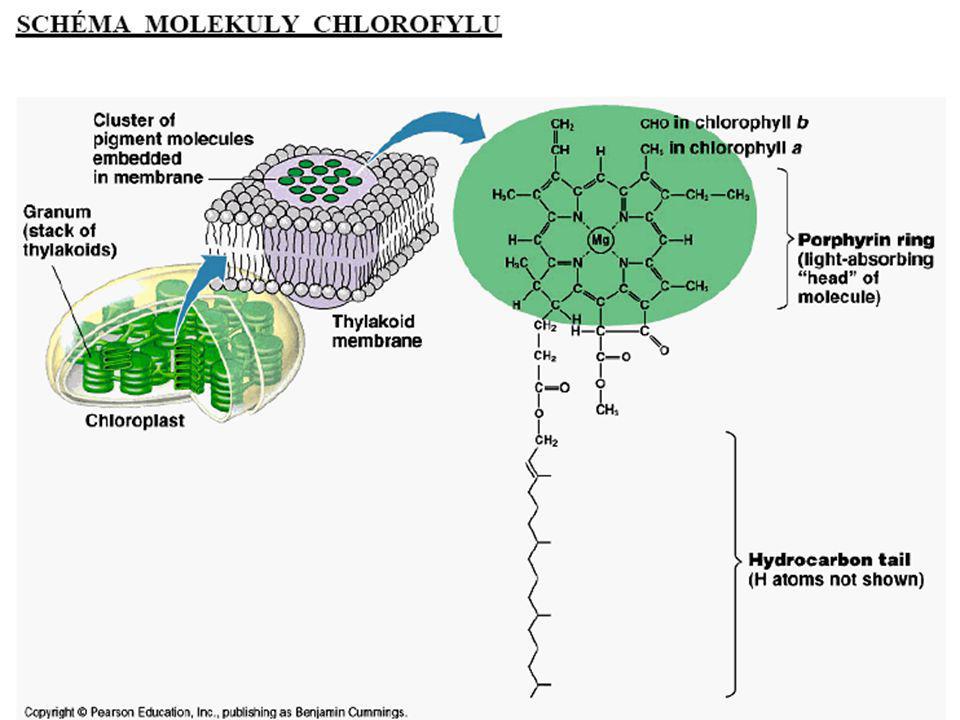
Chlorofyly

34
stroma - obsahuje enzymy Calvinova cyklu (2
stroma - obsahuje enzymy Calvinova cyklu (2. temnostní fáze), DNA, RNA, škrob.
, DNA, RNA, škrob.")
35
předpokladem fotosyntézy, tj
předpokladem fotosyntézy, tj. transformace světelné energie v energii chemických vazeb, je schopnost zelených rostlin absorbovat energii fotonů světla svými elektrony ve fotosyntetických pigmentech - využívají pouze tzv. fotosynteticky účinnou část světla ležící v oblasti viditelného záření o vlnové délce () nm Princip: molekula chlorofylu zachytí kvantum zářivé energie excitace e- na vyšší orbital; excitovaný e- má za normálních podmínek krátkou životnost ( 10-9s) a vrací se zpět na původní energetickou hladinu a část absorbované E se vyzáří do okolí (termální depopulace, fluorescence) na membránách tylakoidů dochází k zvláštnímu fyzikálně-chemickému jevu, tj. k otočení spinu (spinového čísla) excitovaného e-, vznik tzv. tripletového stavu, prodloužení doby excitace 10-9s možnost metabolického využití absorbované světelné E
 nm. Princip: molekula chlorofylu zachytí kvantum zářivé energie excitace e- na vyšší orbital; excitovaný e- má za normálních podmínek krátkou životnost ( 10-9s) a vrací se zpět na původní energetickou hladinu a část absorbované E se vyzáří do okolí (termální depopulace, fluorescence) na membránách tylakoidů dochází k zvláštnímu fyzikálně-chemickému jevu, tj. k otočení spinu (spinového. čísla) excitovaného e-, vznik tzv. tripletového stavu, prodloužení doby excitace 10-9s možnost metabolického využití absorbované světelné E.")
36
B) excitovaný elektron je na základě vysoké elektronegativity vypuzen z molekuly donoru a zachycen akceptorem, donor přechází v KATIONT po doplnění elektronu se vrací do energeticky neutrálního stavu. Výsledek: v obou případech je stejný; dochází k zřetězení jednotlivých elektronových přenašečů s postupně klesajícím elektrochemickým potenciálem, tzv. štafetové předávání elektronů

37
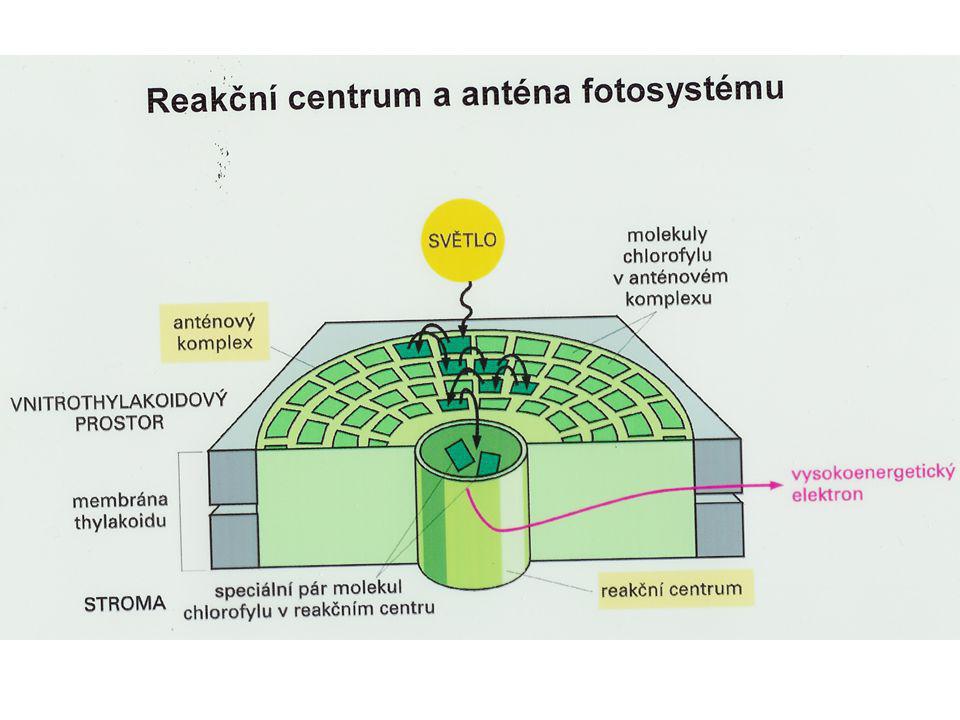
FOTOSYSTÉMY (PS I a PS II)
pigment-proteinové komplexy situované na granálních membránách tylakoidů v chloroplastech; fce: vazba fotosyntetických pigmentů, tj. hlavní a doplňková barviva:

38
PS II – vázán na vnitřní (lipidickou) část membrány, reakční centrum absorbuje světlo o vlnové délce 680 nm, místo fotolýzy H2O PS I – vázán na vnější (proteinovou) část membrány, reakční centrum absorbuje světlo o vlnové délce 700 nm, místo vzniku ATP, NADPH+ H+ chlorofyl „a“ je vázán na reakční centrum, dále antenální molekuly doplňkových pigmentů, účinnost je až 95%
 část membrány, reakční centrum absorbuje světlo o vlnové délce 700 nm, místo vzniku ATP, NADPH+ H+ chlorofyl „a je vázán na reakční centrum, dále antenální molekuly doplňkových pigmentů, účinnost je až 95%")
40
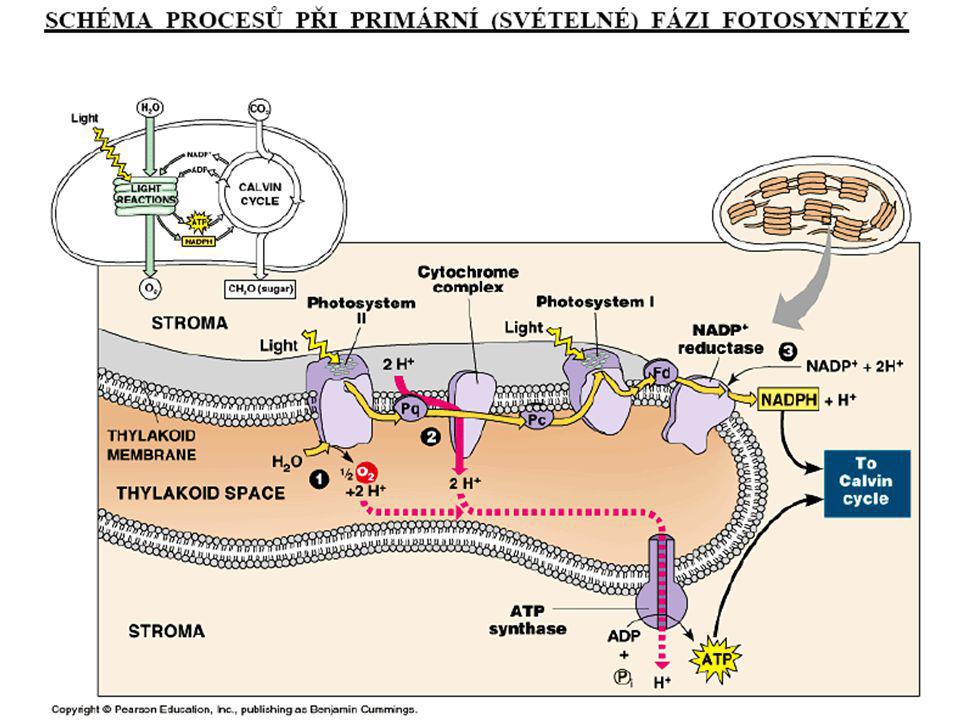
Schéma světlé fáze fotosyntesy

41
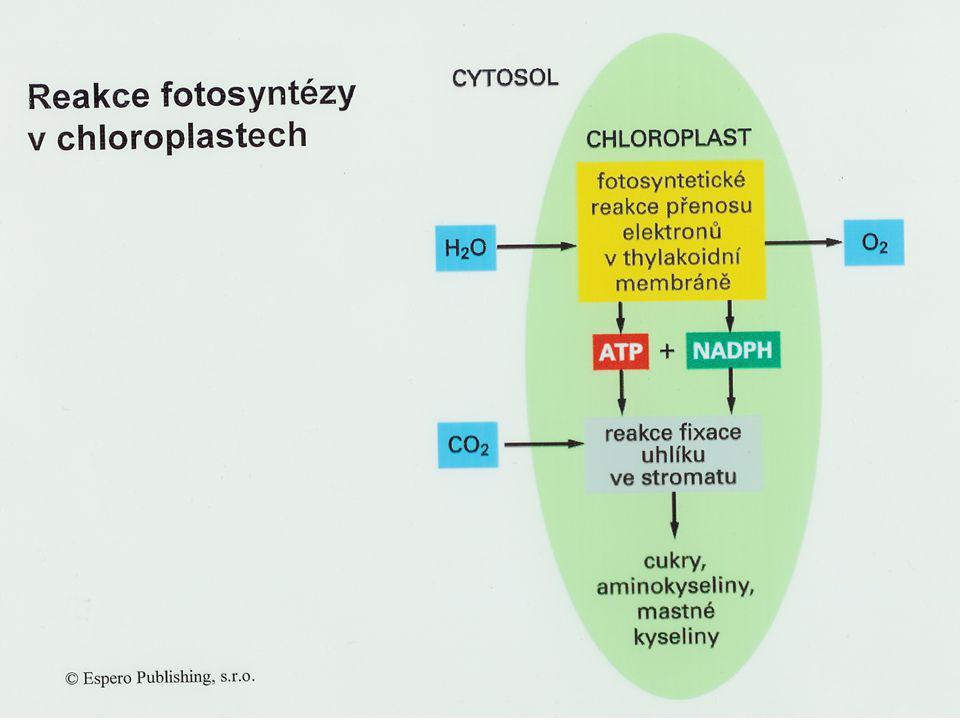
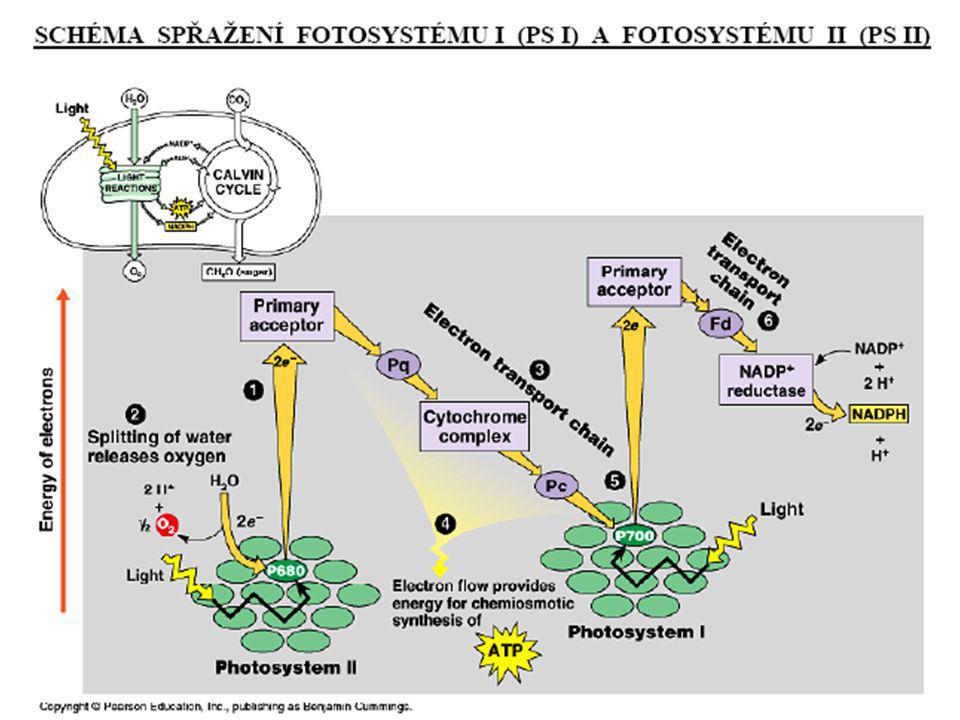
Primární (světelná) fáze fotosynézy
Reakce: 1. fotolýza vody; 2. tvorba ATP, redukovaných koenzymů NADPH+H+; 3. předávání elektronů v rámci kaskády redoxních přenašečů, odstupňovaných hodnotami klesajícího elektrochemického potenciálů. fotofosforylace cyklická fotofosforylace acyklická

42
Primární (světelná) fáze fotosynézy
Základní procesy: a) produkce H+ a jejich hromadění v tylakoidním prostoru, vznik H+ gradientu a ATP; b) transport elektronů přes tylakoidní membránu, pomocí tzv. kaskády redoxních přenašečů na NADP Výsledek: ATP, NADPH+H+, O2
 produkce H+ a jejich hromadění v tylakoidním prostoru, vznik H+ gradientu a ATP; b) transport elektronů přes tylakoidní membránu, pomocí tzv. kaskády redoxních přenašečů na NADP. Výsledek: ATP, NADPH+H+, O2.")
43
Primární (světelná) fáze fotosynézy
(1) absorpce slunečního záření fotoreceptorem a excitace elektronu fotoreceptoru (2) fotolýza vody: H2O H+ + 2e- + 1/2O2 (3) fotoredukce NADP+: NADP+ + 2H+ + 2e NADPH + H+ Protony (2H+) v této reakci pocházejí z fotolýzy vody, z níž byly uvolněny po proběhlých procesech uvedených v bode (1). (4) fotofosforylace: ADP + Pi ATP + H2O protonový gradient
 absorpce slunečního záření fotoreceptorem a excitace elektronu fotoreceptoru. (2) fotolýza vody: H2O 2H+ + 2e- + 1/2O2. (3) fotoredukce NADP+: NADP+ + 2H+ + 2e- NADPH + H+ Protony (2H+) v této reakci pocházejí z fotolýzy vody, z níž byly uvolněny po proběhlých procesech uvedených v bode (1). (4) fotofosforylace: ADP + Pi ATP + H2O. protonový. gradient.")
45
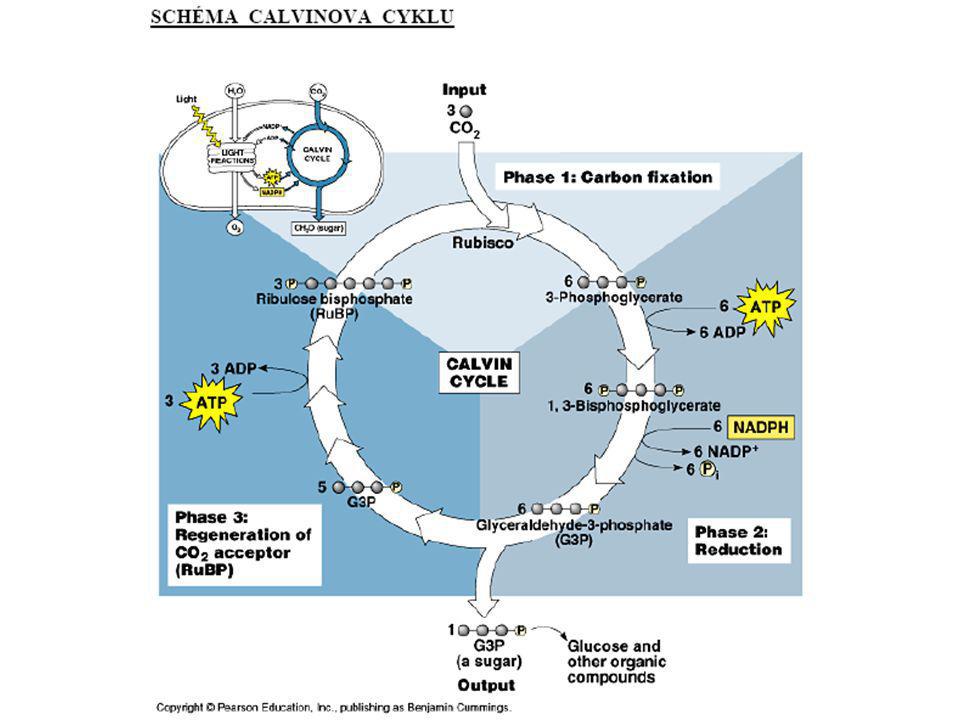
sekundární (temnostní) fáze fotosyntézy
Calvinův cyklus sekundární (temnostní) fáze fotosyntézy - není vázána na světlo, probíhá ve stromatu chloroplastů
 fáze fotosyntézy. - není vázána na světlo, probíhá ve stromatu chloroplastů.")
46
1) 2)
 2)")
48
Syntesa látek z produktů Calvinova cyklu
Sacharosa: vznik mimo chloroplasty; 2GAP F-1,6DP F-6P G-1P (opačná glykolysa); G-1P + UTP UDPG + F-6P sacharosafosfát Biosyntéza sacharózy Škrob: vznik ve stromatu; F-6P G-1P + ATP ADPG (1,4 vazba), podobně vzniká i amylopektin (1,6 vazba) polymerace. Fruktany: vznik ve stromatu; ve vodě rozpustné polymery fruktosy, rychlý zdroj energie (vakuoly).
; G-1P + UTP UDPG + F-6P sacharosafosfát. Biosyntéza sacharózy. Škrob: vznik ve stromatu; F-6P G-1P + ATP ADPG (1,4 vazba), podobně vzniká i amylopektin (1,6 vazba) polymerace. Fruktany: vznik ve stromatu; ve vodě rozpustné polymery fruktosy, rychlý zdroj energie (vakuoly).")
49
Problémy fixace CO2 RUBISCO může fungovat jako karboxylasa (váže CO2, Calvinův cyklus) nebo jako oxidoreduktasa (vazba s O2), v poměru 3:1 =fotorespirace O2 + RUBP PGA + 2P glykolát, problematické a energeticky náročné zpracování produktů fotorespirace, ve 3 různých organelách (chloroplast, peroxisom, mitochondrie) při teplotě a intenzitě ozáření se snižuje koncentrace CO2 v chloroplastech a zvyšuje se poměr fotorespirace
 nebo jako oxidoreduktasa (vazba s O2), v poměru 3:1 =fotorespirace. O2 + RUBP PGA + 2P glykolát, problematické a energeticky náročné zpracování produktů fotorespirace, ve 3 různých organelách (chloroplast, peroxisom, mitochondrie) při teplotě a intenzitě ozáření se snižuje koncentrace CO2 v chloroplastech a zvyšuje se poměr fotorespirace.")
50
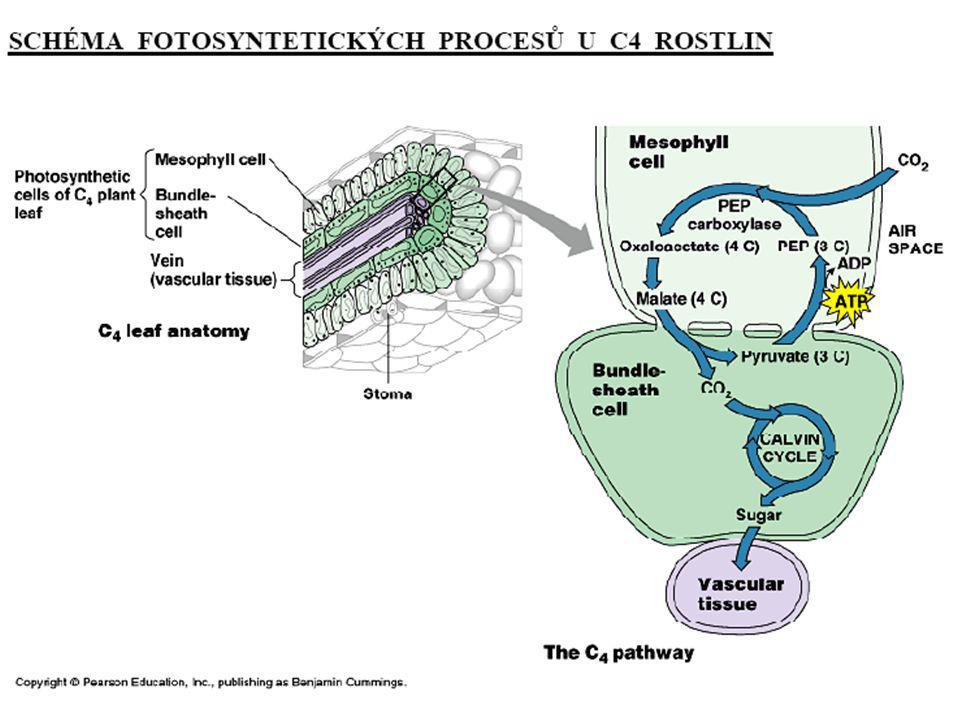
C4-rostliny V C4 rostlinách jsou prostorově separovány děje Calvinova cyklu od dějů Hatchova-Slackova cyklu. Buňky mezofylu, lokalizované blízko povrchu listů, neobsahují RubisCO a proto nemohou zapojit přijatý CO2 do Calvinova cyklu. Přijatý CO2 je v těchto buňkách konvertován v Hatchově-Slackově cyklu na malát (případně jinou čtyřuhlíkatou sloučeninu), který je transportován přes plazmodezmy do hlouběji uložených buněk pochev cévních svazků, kde je dekarboxylován. Tyto buňky obsahují RubisCO a proto za jeho katalytického účinku může být uvolněný CO2 zapojen do Calvinova cyklu. Regenerovaný pyruvát může být transportován zpět do buněk mezofylu a po fosforylaci na fosfoenolpyruvát využit v dalším Hatchově-Slackově cyklu.
, který je transportován přes plazmodezmy do hlouběji uložených buněk pochev cévních svazků, kde je dekarboxylován. Tyto buňky obsahují RubisCO a proto za jeho katalytického účinku může být uvolněný CO2 zapojen do Calvinova cyklu. Regenerovaný pyruvát může být transportován zpět do buněk mezofylu a po fosforylaci na fosfoenolpyruvát využit v dalším Hatchově-Slackově cyklu.")
52
CAM rostliny zástupci čeledí Crasulaceae, Liliaceae, Cactaceae, Orchideaceae Princip - časově izolovaná dvojí karboxylace: 1. noc = otevřené průduchy, fixace CO2 na PEP (fosfoenolpyruvát) vznik malátu, aktivní transport do vakuol (spotřeba 1 ATP) + obnova RUBP (ribulosabisfosfát) (spotřeba 1 ATP). 2. den = zavřené průduchy, malát do Calvinova cyklu.
 vznik malátu, aktivní transport do vakuol (spotřeba 1 ATP) + obnova RUBP (ribulosabisfosfát) (spotřeba 1 ATP). 2. den = zavřené průduchy, malát do Calvinova cyklu.")
54
FAKTORY OVLIVŇUJÍCÍ FOTOSYNTÉZU
koncentrace CO2, teplota, H2O, minerální výživa, imise (rozklad chlorofylu)
")
55
Srovnání fotosynthesy a respirace

Podobné prezentace