Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Miroslav Strnad / Dušan Lazár oddělení biofyziky CRH a Katedra biofyziky, Př.f., UP http://biofyzika.upol.cz Fotosyntéza - temnostní (uhlíkové) reakce
 reakce")
2
Obsah C 3 cyklus (C alvin-Benson-Bassham cyklus) a C3 rostliny Oxidativní C2 cyklus (fotorespirace) C4 cyklus (Hatch-Slackův cyklus) a C4 rostliny CAM cyklus a CAM rostliny Syntéza škrobu a sacharózy Měření světelných a uhlíkových reakcí fotosyntézy
 a C3 rostliny Oxidativní C2 cyklus (fotorespirace) C4 cyklus (Hatch-Slackův cyklus) a C4 rostliny CAM cyklus a CAM rostliny Syntéza škrobu a sacharózy Měření světelných a uhlíkových reakcí fotosyntézy")
3
Temnostní fáze (reakce) fotosyntézy Prvotní představa byla, že temnostní fáze fotosyntézy není vůbec závislá na světle. Tato fáze sice není přímo závislá na světle, ale využívá produktů světelné fáze fotosyntézy - NADPH a ATP) => vhodnější název je „uhlíkové“ reakce (fáze) fotosyntézy.
 => vhodnější název je „uhlíkové reakce (fáze) fotosyntézy..")
4
Calvin-Benson-Bassham cyklus (pšenice, ječmen, hrach, …) Objevili ho Melvin Calvin a významně přispěli i Andrew Benson a James Bassham (=> CBB cyklus). M. Calvin za tento cyklus dostal v roce 1961 Nobelovu cenu za chemii. Popisuje fixaci CO 2 odehrávající se ve stromatu chloroplastů (mezofylových buněk u C3 rostlin). Protože se přímo po fixaci CO 2 tvoří 3-uhlíkové sloučeniny, také se nazývá C3-cyklus. C3 cyklus představuje standardní formu fixace CO 2 u většiny rostlin (tzv. C3 rostliny). Sumárně lze zapsat jako: 3 CO 2 + 6 NADPH + 5 H 2 O + 9 ATP → glyceraldehyd-3-fosfát + 2 H + + 6 NADP + + 9 ADP + 8 P i Často se chybně objevují rovnice, kde produkt je hexóza, což je špatně (psáno na základě oxidativní fosforylace v mitochondriích).
. Protože se přímo po fixaci CO 2 tvoří 3-uhlíkové sloučeniny, také se nazývá C3-cyklus. C3 cyklus představuje standardní formu fixace CO 2 u většiny rostlin (tzv. C3 rostliny). Sumárně lze zapsat jako: 3 CO NADPH + 5 H 2 O + 9 ATP → glyceraldehyd-3-fosfát + 2 H NADP ADP + 8 P i Často se chybně objevují rovnice, kde produkt je hexóza, což je špatně (psáno na základě oxidativní fosforylace v mitochondriích)..")
5
Cyklus má 3 fáze: kyrboxylaci redukci regeneraci

6
Jedná se skutečně o cyklus - karboxylace 3 molekul ribulózy-1,5- bisfosfátu (RuBP) vede ve výsledku (následným užitím i ATP a NADPH) k syntéze 1 molekuly glyceraldehydu-3-fosfát (G3P, GAP) a poté k regeneraci opět 3 molekul ribulózy-1,5- bisfosfátu. Každá reakce je enzymatická = reakce je umožněna daným enzymem.

7
Klíčový enzym cyklu je enzym umožňující karboxylaci 1 molekuly RuBP na 2 molekuly 3-fosfoglycerátu (PGA), nazývaný ribulózy- 1,5-bisfosfát karboxyláza/oxygenáza (Rubisco). Celková reakce je vlastní karboxylace za tvorby nestabilního meziproduktu a následné hydrolýzy. Vstupní CO 2 je zabudováno pouze do 1 z molekul PGA.

8
I když Rubisco je klíčový enzym v asimilaci CO 2, bylo zjištěno, že rychlost asimilace CO 2 je nejvíce kontrolována enzymem sedoheptulóza-1,7-bisfosfatázou a až poté Rubiscem. Vysoké energetické požadavky cyklu naznačují nutnost jeho regulace => v případě kdy není dostatek (ve tmě) všech reaktantů (NADPH, a ATP) je cyklus „vypnutý“ => některé enzymy cyklu jsou ve tmě samovolně deaktivovány a v případě potřeby následně aktivovány. Aktivace některých enzymů C3 cyklu se děje pomocí thioredoxinů (viz dále). Aktivace Rubisca se děje ve 2 krocích: 1) odstranění nežádoucích látek (carboxyarabinitol-1-fosfát, ale i RuBP) navázaných ve tmě na Rubisco před jeho vlastní aktivací 2) tvorba vlastního aktivního stavu Rubisca
 všech reaktantů (NADPH, a ATP) je cyklus „vypnutý => některé enzymy cyklu jsou ve tmě samovolně deaktivovány a v případě potřeby následně aktivovány. Aktivace některých enzymů C3 cyklu se děje pomocí thioredoxinů (viz dále). Aktivace Rubisca se děje ve 2 krocích: 1) odstranění nežádoucích látek (carboxyarabinitol-1-fosfát, ale i RuBP) navázaných ve tmě na Rubisco před jeho vlastní aktivací 2) tvorba vlastního aktivního stavu Rubisca.")
9
ad 1) Děje se pomocí dalšího enzymu – Rubisco aktivázy, která se aktivací Rubisca inaktivuje a poté pomocí ATP a thioredoxinů opět aktivuje. Na obrázku níže vlevo RuBP inhibuje Rubisco, protože je na něho navázané před vlastní aktivací Rubisca (viz dále).
..")
10
ad 2) Rubisco bez navázaných nežádoucích látek musí být nejdříve ztratit 2 protony, pak musí být karbamylováno (navázání CO 2 ) a až navázání Mg 2+ iontů vede k tvorbě aktivního Rubisca. Při navázání CO 2 na Rubisco při jeho karbamalyci, tato molekula CO 2 není ta molekula CO 2, která se fixuje při karboxylaci RuBP!

11
Redukovaný thioredoxin redukuje disulfidické skupiny (–S-S-) enzymů C3 cyklu a tím je aktivuje. Nejdříve se však thioredoxin sám musí redukovat a to se děje pomocí elektronů (pocházejících ze světelné fáze fotosyntézy) z feredoxinu a za přítomnosti enz y mu feredoxin/thioredoxin reduktáza.
 z feredoxinu a za přítomnosti enz y mu feredoxin/thioredoxin reduktáza..")
12
Pomocí thioredoxinů se v C3 cyklu aktivuje: fruktoza-1,6-bisfosfat fosfatáza ribuloza-5-fosfat kináza sedoheptuloza-1,7-bisfosfat fosfatáza NADP/glycerladehyd-3-fosfát dehydrogenáza ale i Rubisco aktiváza

13
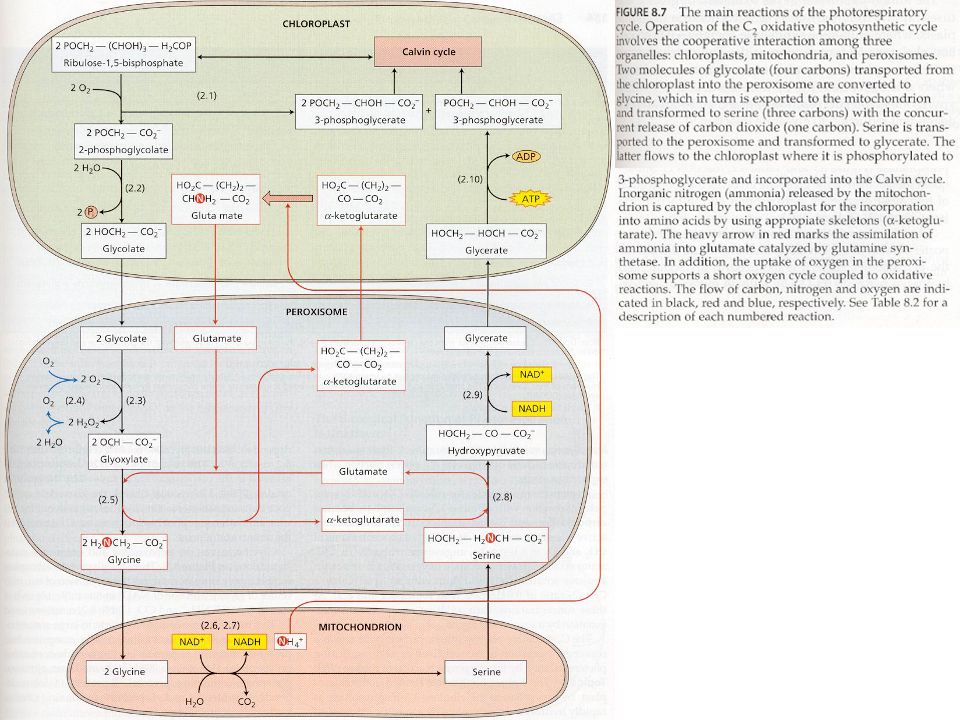
Fotorespirace Rubisco, jak už sám název napovídá (karboxylázaoxygenáza), nekatalyzuje pouze fixaci CO 2 (karboxylace), ale i opačný proces, to je oxygenaci (= navázání O 2, čili dýchaní = respirace). Protože při této reakci (oxygenaci) se opět zpotřebovávají produkty světlené fáze fotosyntézy, tato reakce se nazývá fotorespirace. Protože v cyklu dochází k oxygenaci a 1. meziproduktem fotorespirace je molekula se 2 uhlíky (2-fosfoglykolát), nazývá se sled těchto reakcí jako oxidativní C2 cyklus. Zda-li proběhne asimilace CO 2 nebo fotorespirace závisí na koncentraci CO 2 a O 2. Fotorespirace je tedy jev konkurující asimilaci CO 2 a důvod existence fotorespirace není známý.
 se opět zpotřebovávají produkty světlené fáze fotosyntézy, tato reakce se nazývá fotorespirace. Protože v cyklu dochází k oxygenaci a 1. meziproduktem fotorespirace je molekula se 2 uhlíky (2-fosfoglykolát), nazývá se sled těchto reakcí jako oxidativní C2 cyklus. Zda-li proběhne asimilace CO 2 nebo fotorespirace závisí na koncentraci CO 2 a O 2. Fotorespirace je tedy jev konkurující asimilaci CO 2 a důvod existence fotorespirace není známý..")
14
Celý oxidativní C2 cyklus je prostorově rozdělen a probíhá ve 3 organelách – chloroplastech, peroxizomech a mitochondriích. 2-fosfoglykolát je ještě v chloroplastech přeměněn na glykolát, který putuje do peroxizomu, kde je přeměněn na glycin, který dále putuje do mitochondrií, kde je přeměněn na serin, za uvolnění CO 2 (proto „dýchání“). Serin je zpět transportován do peroxizomu, kde je přeměněn na glycerát, který putuje zpět do chloroplastu, kde je přeměněn na 3-fosfoglycerát, který se pak dále účastní C3 cyklu v chloroplastu. Celý cyklus je také propojen přes využití anorganického dusíku (amoniaku, NH 4 + ) uvolněného v mitochondriích při přeměně glycinu na serin. Toto NH 4 + putuje z mitochondrií do chloroplastu, kde je zabudováván do glutaminu a následně do kyseliny glutaminové (= glutamát) (oba jsou aminokyseliny), která se dále účastní reakcí v peroxizomu – viz obr ázek na další straně. Část C 2 cyklu je také spojena se spotřebou a obnovou O 2 v peroxizomu – viz obrázek na další straně.
. Serin je zpět transportován do peroxizomu, kde je přeměněn na glycerát, který putuje zpět do chloroplastu, kde je přeměněn na 3-fosfoglycerát, který se pak dále účastní C3 cyklu v chloroplastu. Celý cyklus je také propojen přes využití anorganického dusíku (amoniaku, NH 4 + ) uvolněného v mitochondriích při přeměně glycinu na serin. Toto NH 4 + putuje z mitochondrií do chloroplastu, kde je zabudováván do glutaminu a následně do kyseliny glutaminové (= glutamát) (oba jsou aminokyseliny), která se dále účastní reakcí v peroxizomu – viz obr ázek na další straně. Část C 2 cyklu je také spojena se spotřebou a obnovou O 2 v peroxizomu – viz obrázek na další straně..")
16
Některé rostliny si vyvinuly mechanismy, jak zvyšovat množství CO 2 v zásobě a tím eliminovat fotorespiraci. Jedná se o tzv.: 1) C4 rostliny a C4 cyklus 2) CAM rostliny a CAM cyklus C4 rostliny a C4 cyklus (kukuřice, proso, cukrová třtina, …) Protože cyklus u C4 rostlin začíná syntézou 4-uhlíkatých kyselin, cyklus se nazývá C4 cyklus. Cyklus se také nazývá Hatch-Slackův cyklus (objeven v roce 1966 Hatchem a Slackem) C4 cyklus je spojen s rozdílnou anatomií C4 rostlin v porovnání s C3 rostlinami: - U C3 rostlin, pouze mezofylové buňky mají chloroplasty, ve kterých také probíhá i C3 cyklus. - C4 rostliny mají chloroplasty jak v mezofylových buňkách, tak i v buňkách pochev svazků cévních (bundle sheath cells). V jednotlivých typech buněk probíhají jednotlivé fáze C4 cyklu => jednotlivé typy buněk musí být blízko u sebe. Transport metabolitů mezi jednotlivými typy buněk je umožněn pomocí plasmodesmat = kanály v buněčných stěnách rostlinných buňek.
 C4 rostliny a C4 cyklus 2) CAM rostliny a CAM cyklus C4 rostliny a C4 cyklus (kukuřice, proso, cukrová třtina, …) Protože cyklus u C4 rostlin začíná syntézou 4-uhlíkatých kyselin, cyklus se nazývá C4 cyklus. Cyklus se také nazývá Hatch-Slackův cyklus (objeven v roce 1966 Hatchem a Slackem) C4 cyklus je spojen s rozdílnou anatomií C4 rostlin v porovnání s C3 rostlinami: - U C3 rostlin, pouze mezofylové buňky mají chloroplasty, ve kterých také probíhá i C3 cyklus. - C4 rostliny mají chloroplasty jak v mezofylových buňkách, tak i v buňkách pochev svazků cévních (bundle sheath cells). V jednotlivých typech buněk probíhají jednotlivé fáze C4 cyklu => jednotlivé typy buněk musí být blízko u sebe. Transport metabolitů mezi jednotlivými typy buněk je umožněn pomocí plasmodesmat = kanály v buněčných stěnách rostlinných buňek..")
17
Struktura listu C4 rostlin a plasmodesmata

18
C4 cyklus má 4 kroky: 1) Fixace CO 2 (ten nejdříve pomocí karbonic anhydrázy změněn na HCO 3 - ) do fosfoelnolpyruvatu (PEP) za vzniku 4-uhlíkatých kyselin v mezofylových buňkách. 2) Transport těchto kyselin do buněk pochev svazku cévních. 3) Dekarboxylace (= oddělení CO 2 ) těchto kyselin v buňkách pochev svazku cévních za vzniku 3- uhlíkatých kyselin a generování velké koncentrace CO 2. Zde probíhá standardní Calvinův cyklus, ale díky velké koncentraci CO 2 je minimalizována fotorespirace. 4) Transport 3-uhlíkatých kyselin zpět do mezofylových buněk, kde se z nich regeneruje PEP.
 Transport těchto kyselin do buněk pochev svazku cévních. 3) Dekarboxylace (= oddělení CO 2 ) těchto kyselin v buňkách pochev svazku cévních za vzniku 3- uhlíkatých kyselin a generování velké koncentrace CO 2. Zde probíhá standardní Calvinův cyklus, ale díky velké koncentraci CO 2 je minimalizována fotorespirace. 4) Transport 3-uhlíkatých kyselin zpět do mezofylových buněk, kde se z nich regeneruje PEP..")
19
Podle toho, jaká 4- a 3-uhlíkatá kyselina vzniká na konci kroků 1 (malát x aspartát) a 3 (pyruvát x alanin) a podle toho, jaký enzym katalyzuje dekarboxylaci na začátku kroku 3, se C4 cyklus ještě rozděluje na 3 další typy (jeden z nich na obrázku nahoře).
 a 3 (pyruvát x alanin) a podle toho, jaký enzym katalyzuje dekarboxylaci na začátku kroku 3, se C4 cyklus ještě rozděluje na 3 další typy (jeden z nich na obrázku nahoře).")
20
CAM rostliny a CAM cyklus (kaktusy, agáve, ananas, …) CAM = Crassulecean Acid Metabolism, pojmenováno podle čeledi rostlin Crassulaceae = tlusticovité, u kterých byl tento cyklus poprvé objeven. Na rozdíl od C4 cyklu, kde fixace CO 2 a karboxylace jsou prostorově odděleny, u CAM cyklu je fixace CO 2 a karboxylace oddělena časově: - příjem CO 2 se děje v noci, kdy jsou otevřená stomata (= průduchy) rostlin. V noci také dochází (v cytoplazmě) k fixaci CO 2 do PEP za vzniku malátu a následně koncového produktu kyseliny jablečné (malic acid), která se uchovává ve vakuolách - karboxylace se děje ve dne, kdy jsou stomata uzavřena: ve dne kyselina jablečná vystupuje z vakuol, mění se na malát (v cytoplazmě) a z toho se dekarboxylací uvolní CO 2, který je následně v chloroplastu použitý ke karboxylaci ve standardním C3 cyklu Protože ve dne mají CAM rostliny uzavřená stomata, hromadí se ve dne u CAM rostlin CO 2, čímž se opět eliminuje fotorespirace (jako u C4 cyklu). Navíc, protože při zavřených průduších nedochází k transpiraci (= vypařování vody), CAM cyklus výrazně šetří vodou v rostlině => je převážně u rostlin žijících v suchém prostředí.
 rostlin. V noci také dochází (v cytoplazmě) k fixaci CO 2 do PEP za vzniku malátu a následně koncového produktu kyseliny jablečné (malic acid), která se uchovává ve vakuolách - karboxylace se děje ve dne, kdy jsou stomata uzavřena: ve dne kyselina jablečná vystupuje z vakuol, mění se na malát (v cytoplazmě) a z toho se dekarboxylací uvolní CO 2, který je následně v chloroplastu použitý ke karboxylaci ve standardním C3 cyklu Protože ve dne mají CAM rostliny uzavřená stomata, hromadí se ve dne u CAM rostlin CO 2, čímž se opět eliminuje fotorespirace (jako u C4 cyklu). Navíc, protože při zavřených průduších nedochází k transpiraci (= vypařování vody), CAM cyklus výrazně šetří vodou v rostlině => je převážně u rostlin žijících v suchém prostředí..")
21
Noční fixace CO 2 do PEP a vznik malátu se děje v cytoplazmě dané buňky. Malát opětovně vzniklý ve dne se dekarboxyluje opět v cytoplazmě a produkty dekarboxylace (CO 2 a pyruvát) putují do chloroplastu, kde jsou použity (ve dne) pro karboxylaci ve standradním C3 cyklu a syntéze škrobu (starch), který je v noci zpětně použit na tvorbu PEP.
 putují do chloroplastu, kde jsou použity (ve dne) pro karboxylaci ve standradním C3 cyklu a syntéze škrobu (starch), který je v noci zpětně použit na tvorbu PEP..")
22
Protože 2 protichůdné reakce, karboxylace PEP v noci a dekarboxylace malátu ve dne, probíhají v cytoplazmě, ale v jinou dobu, je potřeba existence nějaké časové regulace => PEP- karboxyláza (Pepka) je v noci pomocí kinázy fosforylována a tím je převedena do aktivního stavu, který není citlivý na malát a naopak ve dne je Pepka fosfatázou defosforylována do stavu, jehož aktivita je inhibována malátem. Fosforylace Pepky pomocí kinázy je ř ízena expresi genů kinázy v odezvě na denní rytmus, kde zpětnovazebná regulace exprese kinázy je řízena pravděpodobně malátem z cytoplasmy, jehož množství je řízeno denním rytmem.

23
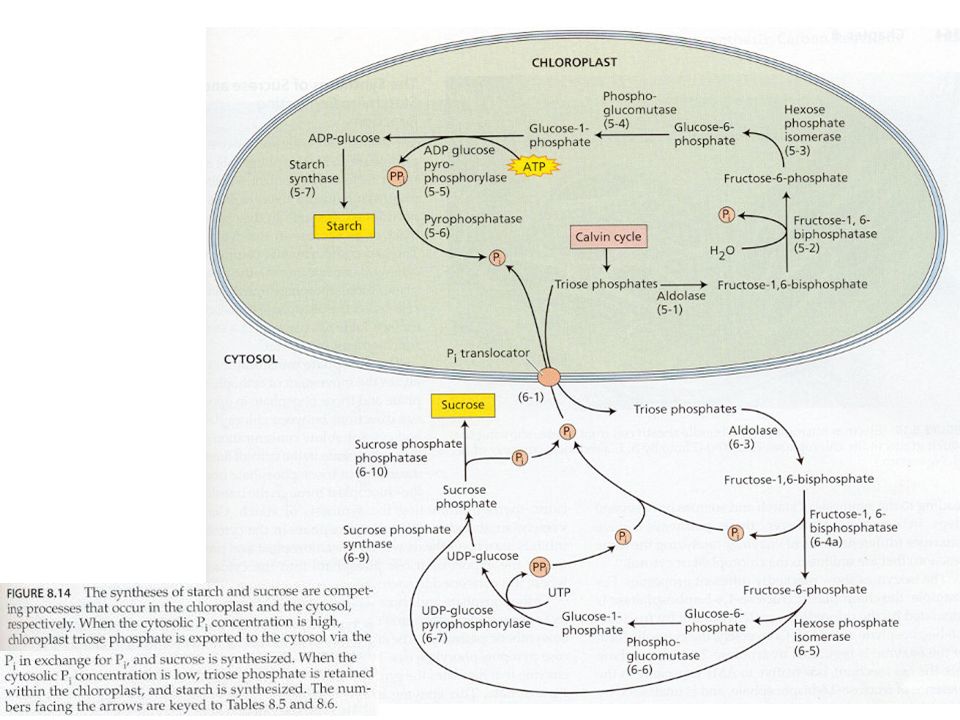
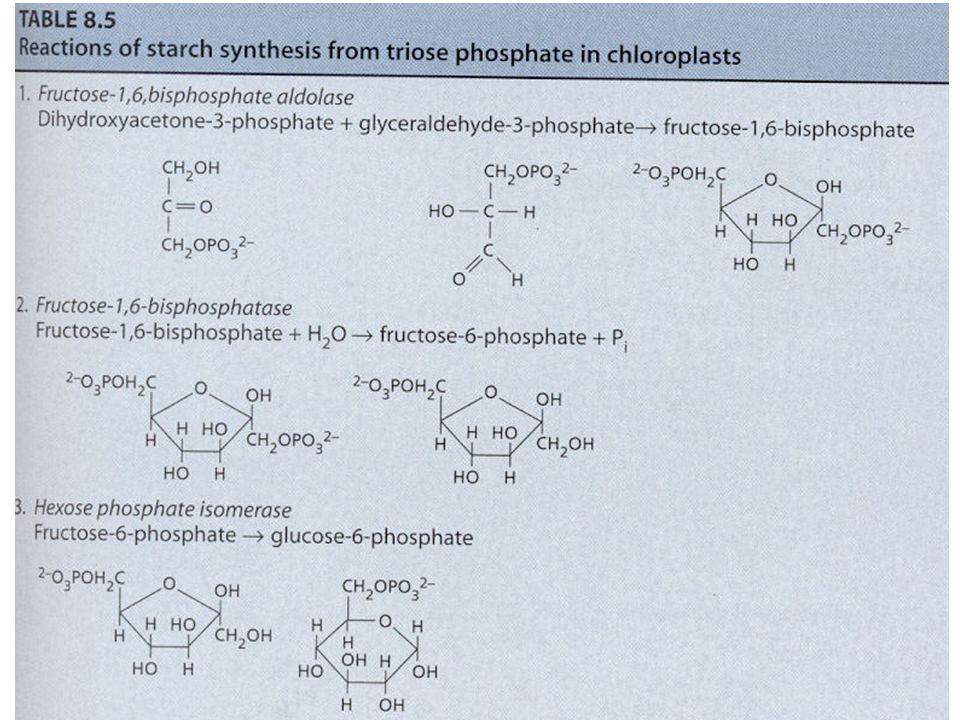
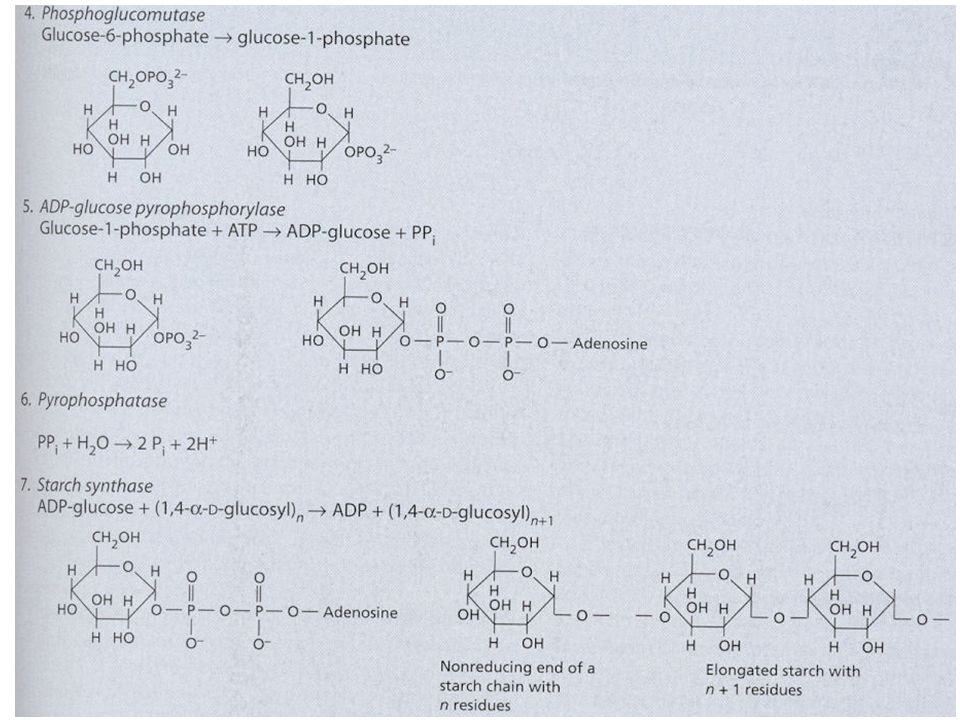
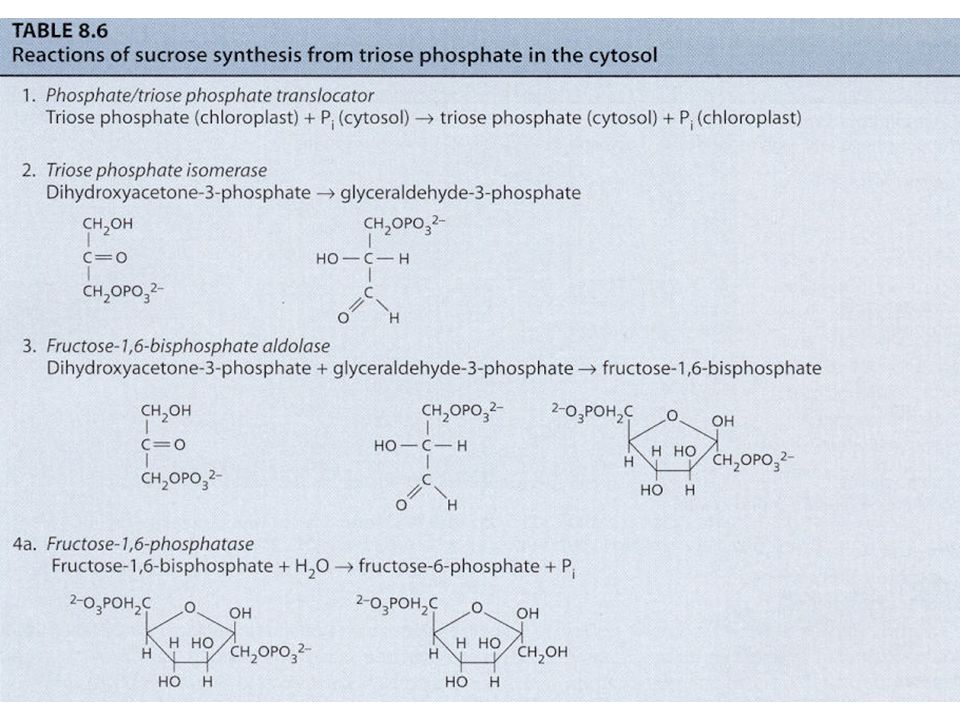
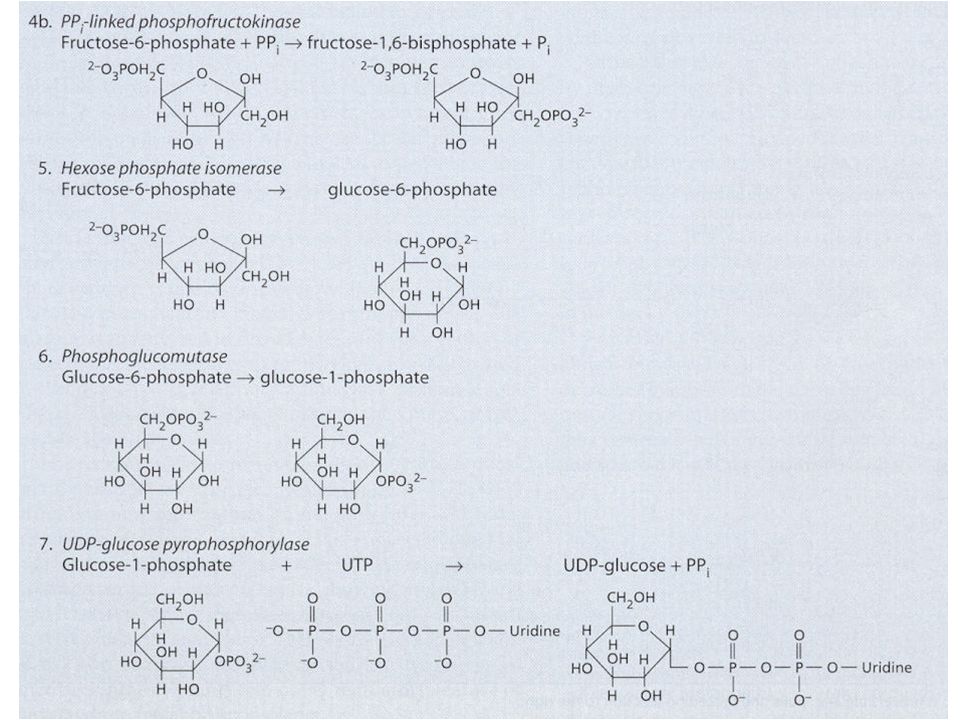
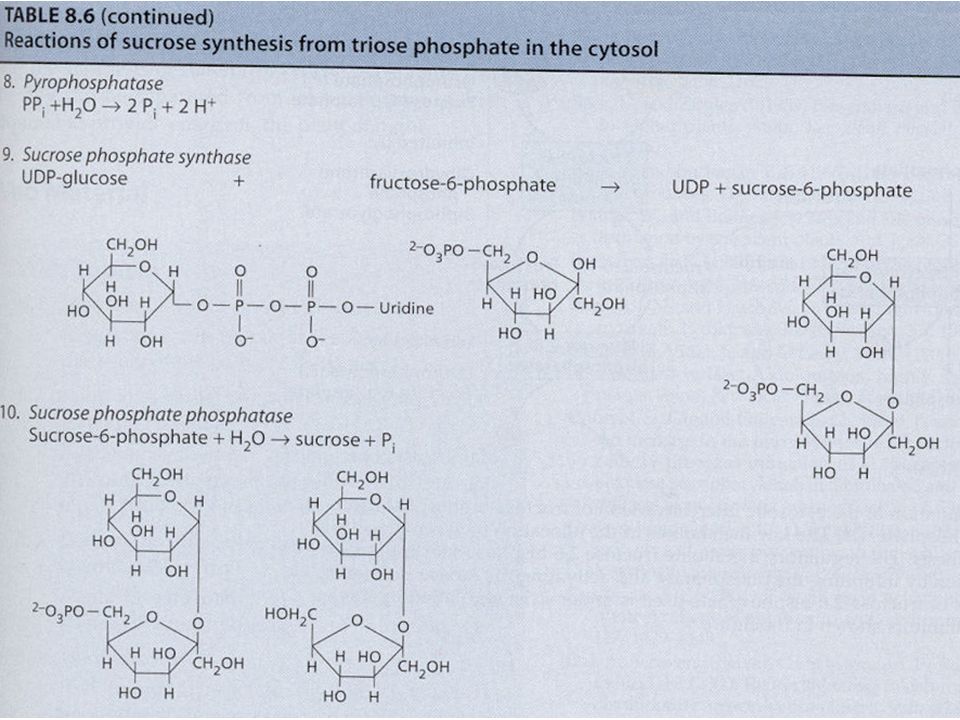
Syntéza škrobu a sacharózy Jak již bylo uvedeno u C3 cyklu, karboxylace 3 molekul ribulózy-1,5- bisfosfátu (RuBP) vede ve výsledku k syntéze 1 molekuly glyceraldehydu-3-fosfát (G3P, GAP; = triozofosfát, TP). Tyto molekuly TP jsou použity pro syntézu škrobu a sacharózy. Syntéza škrobu a sacharózy jsou konkurující si reakce. Škrob se tvoří v chloroplastech a sacharóza v cytoplazmě buňky. Zda-li se bude tvořit škrob nebo sacharóza je převážně řízeno koncentrací anorganického fosforu (ortofosfát, Pi) v cytoplazmě. Pi vstupuje/vystupuje do/z chloroplastu záměnou za TP pomocí Pi translokátorů. Je-li hodně Pi v cytoplazmě, je tento Pi zaměněn za TP z chloroplastu a tento TP v cytoplazmě stimuluje enzym UDP- glukozapyrofosforyláza a dojde ke tvorbě sacharózy. Opačně, je-li Pi v cytoplazmě málo, TP zůstává v chloroplastu a Pi v cytoplazmě – Pi v cytoplazmě působí inhibičně na předchozí enzym => nedochází ke tvorbě sacharózy v cytoplazmě, ale ke tvorbě škrobu v chloroplastech.
 v cytoplazmě. Pi vstupuje/vystupuje do/z chloroplastu záměnou za TP pomocí Pi translokátorů. Je-li hodně Pi v cytoplazmě, je tento Pi zaměněn za TP z chloroplastu a tento TP v cytoplazmě stimuluje enzym UDP- glukozapyrofosforyláza a dojde ke tvorbě sacharózy. Opačně, je-li Pi v cytoplazmě málo, TP zůstává v chloroplastu a Pi v cytoplazmě – Pi v cytoplazmě působí inhibičně na předchozí enzym => nedochází ke tvorbě sacharózy v cytoplazmě, ale ke tvorbě škrobu v chloroplastech..")
25
Pro regulaci syntézy sacharózy a štěpení cukrů (glykolýza) v cytoplazmě je důležitá vzájemná přeměna fruktóza-1,6-bisfosfátu na fruktóza-6-fosfát enzymem fruktóza-1,6-bisfosfatáza a opačně enzymem fruktóza-6-fosfát kináza. Tyto enzymy jsou buď inhibované, nebo aktivované fruktóza-2,6-bisfosfátem, jehož tvorba je také regulována metabolity C3 cyklu.

31
Měření světelných a uhlíkových reakcí fotosyntézy Absorpční spektra a kinetika: excitovaný nebo základní stav, ale i redukovaný nebo oxidovaný stav molekuly absorbuje energii při charakteristických vlnových délkách. Sledováním poměrů absorpcí (excitovaný x základní stav a redukovaný x oxidovaný stav) lze zjistit počáteční stavy molekul. Sledováním kinetiky změn absorpce lze zjistit rychlostní konstanty „přeměny“ molekul, to je přenosu energie a elektronů. Možno měřit v časech od stovek femtosekund (přenos energie) po sekundy (přenos elektronů). Metoda je vhodná pro studium světelné fáze fotosyntézy, hlavně okamžitě po osvětlení, kdy ještě nefungují uhlíkové reakce fotosyntézy a měřené absorpční změny odrážejí pouze světelné reakce fotosyntézy. Příklad – měření kinetiky absorpce při 820 nm odráží redoxní stav převážně P700 +, ale i oxidovaného PC
 lze zjistit počáteční stavy molekul. Sledováním kinetiky změn absorpce lze zjistit rychlostní konstanty „přeměny molekul, to je přenosu energie a elektronů. Možno měřit v časech od stovek femtosekund (přenos energie) po sekundy (přenos elektronů). Metoda je vhodná pro studium světelné fáze fotosyntézy, hlavně okamžitě po osvětlení, kdy ještě nefungují uhlíkové reakce fotosyntézy a měřené absorpční změny odrážejí pouze světelné reakce fotosyntézy. Příklad – měření kinetiky absorpce při 820 nm odráží redoxní stav převážně P700 +, ale i oxidovaného PC.")
32
Fluorescenční spektra a kinetika: fluorescence představuje konkurenční způsob využití excitační energie k fotosyntetickému využití => funguje-li fotosyntéza je emise fluorescence malá a naopak. Opět lze měřit spektra nebo kinetiku a metoda je opět vhodná pro studium světelné fáze fotosyntézy. Příklad – měření kinetiky fluorescence – tzv. OJIP křivky fotosyntetický aparát absorpce světla emise fluorescence fotochemie disipace na teplo ↑ využití světla na fotochemii pak ↓ emise fluorescence a naopak

33
Rychlost vývoje O 2 : možno měřit polarograficky nebo absorpčně (tzv. Hillovy reakce). Časně po osvětlení (do 1 sekundy) odráží převážně světelnou fázi fotosyntézy, v delších časech pak i uhlíkovou fázi fotosyntézy. Rychlost asimilace CO 2 : měří se při dané intenzitě světla a dané koncentraci CO 2. Z měření lze vyhodnotit závislost rychlosti asimilace CO 2 (P) na intenzitě světla (I) – tzv. PI křivka nebo na koncentraci CO 2 – tzv. CO 2 křivka (PI křivku lze měřit i pro rychlost vývoje O 2, kde P znamená fotosyntézu). Metoda je vhodná převážně pro studium uhlíkové fáze fotosyntézy, ale odráží v sobě i světelnou fázi fotosyntézy. Příklad – PI křivky (pro rychlost vývoje O 2 )
. Časně po osvětlení (do 1 sekundy) odráží převážně světelnou fázi fotosyntézy, v delších časech pak i uhlíkovou fázi fotosyntézy. Rychlost asimilace CO 2 : měří se při dané intenzitě světla a dané koncentraci CO 2. Z měření lze vyhodnotit závislost rychlosti asimilace CO 2 (P) na intenzitě světla (I) – tzv. PI křivka nebo na koncentraci CO 2 – tzv. CO 2 křivka (PI křivku lze měřit i pro rychlost vývoje O 2, kde P znamená fotosyntézu). Metoda je vhodná převážně pro studium uhlíkové fáze fotosyntézy, ale odráží v sobě i světelnou fázi fotosyntézy. Příklad – PI křivky (pro rychlost vývoje O 2 ).")
Podobné prezentace

>")
















>")
