Markery asistovaná selekce
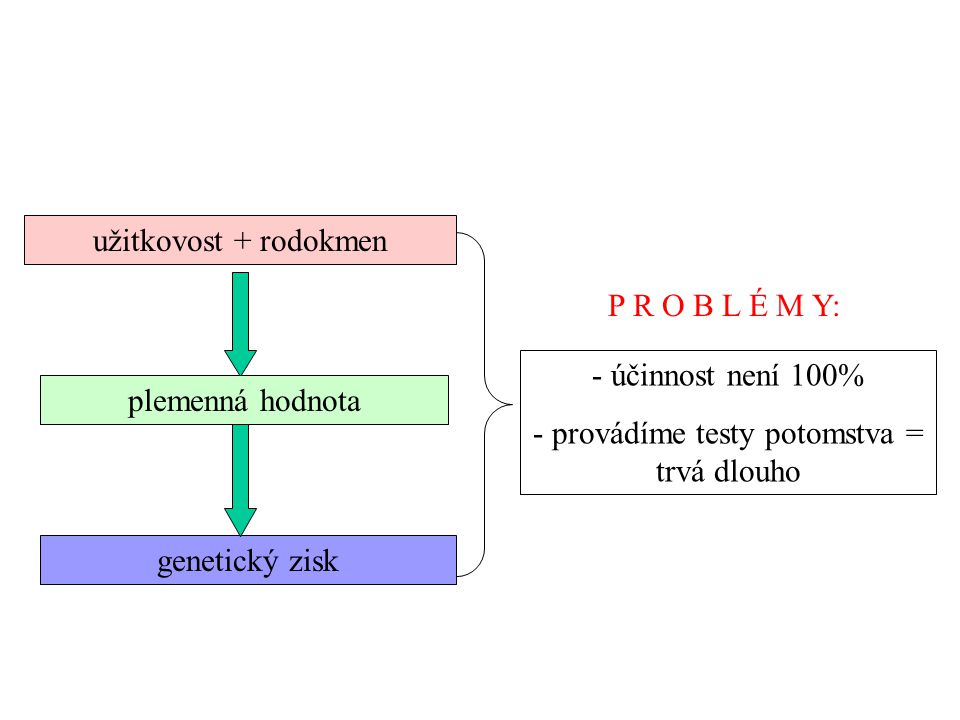
- provádíme testy potomstva = trvá dlouho užitkovost + rodokmen P R O B L É M Y: - účinnost není 100% - provádíme testy potomstva = trvá dlouho plemenná hodnota genetický zisk
P O Ž A D U J E M E E F E K T I V I T U: možnost provést výběr u mladých zvířat; zkrácení generačního intervalu vyšší selekční efekt (zisk za rok) H L E D Á M E geny spojené s vysokou užitkovostí resp. lokusy, kde se tyto geny nacházejí tj. lokusy kvantitativních vlastností = QTL
víme, že vlastnosti ovlivňuje mnoho genů s malým účinkem předpokládáme, že účinek některých polygenů může být výraznější = majorgeny
hledáme genetické markery neznáme genotyp v QTL umístění QTL hledáme genetické markery M A R K E RY místa, kde se sekvence DNA jedinců liší snadno detekovatelný současnými metodami ve vazbě s QTL
přímé markery snadnější využití marker je součástí QTL genotyp markeru vždy odpovídá genotypu QTL halotanový gen myostatinový gen
riziko využití na genetickém efektu se podílí více než jedna mutace (např. myostatin, několik mutací uvnitř tohoto genu dává stejný výsledný efekt) nesprávná identifikace kandidátního genu jako majorgenu
nepřímé markery jsou lokalizovány poblíž QTL nejsou příčinou mutace nevíme, zda je s QTL vázán jeden nebo několik markerů
u nepřímých markerů vazba QTL – marker je specifická podle rodin; sledujeme nejméně 2 generace (plemeník x potomci) využíváme informace o fenotypu potomků vazbu nezjistíme, pokud je plemeník homozygotní v markeru nebo QTL
postup detekce markeru stanovení vazbové mapy určení podílu proměnlivosti vysvětlené markerem σv2 identifikace jedinců, kteří nesou žádoucí alelu a jejich využití ve šlechtění
vazba marker – QTL Q = varianta QTL s účinkem +α q = varianta QTL s účinkem 0 M = dominantní varianta markeru m = recesivní varianta markeru
genetické mapy udávají: pořadí genů vzdálenost genů vzdálenost = pravděpodobnost, s jakou dochází k rekombinaci 1cM = 1 rekombinace / 100 meióz 10cM = 10 rekombinací / 100 meióz
segregace QTL v rámci rodin sledujeme skupiny polosourozenců potomci heterozygotních otců gamety rekombinace pravděpodobnost M Q není q je m
marker QTL frekvence průměr M Q 1–r / 2 μ + α q r / 2 μ m Průměr pro stanovení účinku QTL srovnáme očekávané průměry skupin potomků podle předané alely marker QTL frekvence průměr M Q 1–r / 2 μ + α q r / 2 μ m Průměr [(1-r)(μ + α) / 2 + (r . μ) /2] / 0,5 = μ + (1-r) α [r(μ + α) / 2 + (1 - r) μ /2] / 0,5 = μ + r.α Rozdíl (1-2r) α
rekombinace je blízká hodnotě 0,5 pokud není rozdíl mezi skupinami s alelou M a m pak nemáme QTL efekt QTL je nízký rekombinace je blízká hodnotě 0,5 z diference (1-2r) α nerozlišíme, zda je velký účinek QTL a volná vazba je nízký účinek QTL a pevná vazba
detekce QTL – narušení vazby vztah marker - alelaa využití 2 markerů intervalové mapování (REML) regresní metody detekce QTL – narušení vazby křížení inbredních linií zpětné křížení křížení outbredních populací
Grandsire design (Weller et al. 1990) metodický postup pro stanovení vztahu markeru a vlastnosti vypracovaný pro dojený skot; v plánu pokusu vycházíme od dědů – heterozygotů (Qq) v daném markeru; u jejich synů zařazených v inseminaci zjistíme alelu přenesenou od děda (Qxq); syny rozdělíme do podskupin podle přenesené alely; syny vyhodnotíme podle užitkovosti dcer (kontrola dědičnosti); rozdíly mezi průměrnými plemennými hodnotami mezi skupinami uvnitř dědů jsou způsobeny markerem
význam využití informace o QTL nízká dědivost vlastnosti; vlastnost se projevuje jen u jednoho pohlaví; znak není měřitelný před dospělostí jedince měření znaku je obtížné nebo vyžaduje usmrcení zvířete
význam je odvozen od velikosti účinku QTL frekvence Q alely v populaci: pokud je v populaci fixovaná, MAS nemůže přinést zlepšení pravděpodobnosti, že Mm jedinec předá Q alelu
zpřesnění odhadu plemenné hodnoty
modelování efektu genotypu QTL y = Xb + Wq + Zu + e
může přinést krátkodobé zvýšení efektu selekce o 2-30%; nelze počítat, že nahradí klasickou selekci založenou na principech populační genetiky; může negativně ovlivnit odezvu na selekci u ostatních polygenů.