Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Strategie regulace (proteinové) enzymové aktivity.
Allosterická kontrola. Allosterické proteiny obsahují regulační místa odlišná od substrátových. Allosterický z řečtiny „allos“ = další, „steros“ = uspořádání. Skládají se z podjednotek (protomerů). Mají schopnost kooperativity. Jako příklady enzym aspartáttranskarbamoylasa (ATCasa) a neenzymový kyslíkový přenašeč hemoglobin. Mnohočetné formy enzymů. Isozymy – jsou homologní enzymy katalyzující stejnou reakci, ale lišící se jemně v struktuře a více v Km a Vlim a regulačních vlastnostech.
. Mají schopnost kooperativity. Jako příklady enzym aspartáttranskarbamoylasa (ATCasa) a neenzymový kyslíkový přenašeč hemoglobin. Mnohočetné formy enzymů. Isozymy – jsou homologní enzymy katalyzující stejnou reakci, ale lišící se jemně v struktuře a více v Km a Vlim a regulačních vlastnostech.")
2
3. Reversibilní kovalentní modifikace
3. Reversibilní kovalentní modifikace. Katalytické vlastnosti řady enzymů se mění po kovalentní vazbě nějaké skupiny na jejich molekulu – nejčastěji fosforylace. Modifikující enzymy jsou proteinkinasy a proteinfosfatasy. 4. Proteolytická aktivace. Mnoho proteinů je syntetizováno v neaktivní formě. U enzymů jsou to zymogeny (proenzymy). Po odštěpení části řetězce přechází zymogen na aktivní enzym. Jako aktivující složky působí proteolytické enzymy jako chymotrypsin, pepsin a trypsin. Aktivace je ireversibilní. Kaspasy, proteolytické enzymy, účastnící se programové buněčné smrti, jsou aktivovány z formy prokaspas. Dalším příkladem je kaskáda enzymů při srážení krve. 5. Kontrola množstvím přítomného enzymu-kontrola na úrovni transkripce.
. Po odštěpení části řetězce přechází zymogen. na aktivní enzym. Jako aktivující složky působí proteolytické enzymy jako chymotrypsin, pepsin a trypsin. Aktivace je ireversibilní. Kaspasy, proteolytické enzymy, účastnící se programové buněčné smrti, jsou aktivovány. z formy prokaspas. Dalším příkladem je kaskáda enzymů při srážení krve. 5. Kontrola množstvím přítomného enzymu-kontrola na úrovni transkripce.")
3
ALLOSTERIE Aspartáttranskarbamoylasa jako příklad allosterického enzymu. Allosterické enzymy se neřídí kinetikou Michaelise a Mentenové. Modely allosterie: Symetrický model ( Jacques Monod, Jeffries Wyman a Jean-Pierre Changeux – podle autorů zkracovaný jako MWC model). Angl. concerted (přel. současný) Sekvenční model (Daniel Koshland). Conceptual insights – Stryer, Chapter 10. Hemoglobin je příkladem neenzymového allosterického proteinu
. Angl. concerted (přel. současný) Sekvenční model (Daniel Koshland). Conceptual insights – Stryer, Chapter 10. Hemoglobin je příkladem neenzymového allosterického proteinu.")
4
Enzym Aspartáttranskarbamoylasa (EC 2. 1. 3
Enzym Aspartáttranskarbamoylasa (EC ) je allostericky inhibován konečným produktem metabolické cesty cytidintrifosfátem (CTP). Inhibice zpětnou vazbou. ATCasa katalyzuje kritický stupeň kondenzace Asp s karbamoylfosfátem při syntéze pyrimidinových nukleotidů.
 je allostericky inhibován konečným produktem metabolické cesty cytidintrifosfátem (CTP). Inhibice zpětnou vazbou. ATCasa katalyzuje kritický stupeň kondenzace Asp s karbamoylfosfátem při syntéze pyrimidinových nukleotidů.")
5
Cytidintrifosfát (CTP) inhibuje ATCasu
Cytidintrifosfát (CTP) inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.
 inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.")
6
Od slide 6 po slide 13 je prezentován způsob objasnění struktury ATCasy. Není podstané pro pochopení principu allosterie !! Modifikace cysteinových vedlejších řetězců (ATCasy). Oddělení podjednotek chromatografií na iontoměniči nebo centrifugací.
. Oddělení podjednotek chromatografií na iontoměniči nebo centrifugací..")
7
Centrifugační studie ATCasy.
Centrifugace v gradientu sacharosy. Sedimentační koeficient nativního enzymu je 11,6 S. Dvě podjednotky 2,8 S a 5,8 S. Svedberg (zkratka S či Sv), též Svedbergova jednotka, je odvozená jednotka fyzikální veličiny známé jako sedimentační koeficient. Tato veličina udává čas, za který proběhne sedimentace dané makromolekuly při její ultracentrifugaci. Platí: 1 Svedberg = 1 S = sekundy Jak od sebe oddělíme obě podjednotky a jak lze odstranit p-hydroxymerkuribenzoát ? Větší podjednotka (c) je katalytická, ale není inhibována CTP. Složena ze tří řetězců po 34 kD se označuje c3. Regulační podjednotka (r) váže CTP. Složena ze dvou řetězců po 17 kD je označena r2. Nativní enzym je složen ze dvou katalytických trimerů a tří regulačních dimerů (2 c3 + 3 r2 = c6r6).
, též Svedbergova jednotka, je odvozená jednotka fyzikální veličiny známé jako sedimentační koeficient. Tato veličina udává čas, za který proběhne sedimentace dané makromolekuly při její ultracentrifugaci. Platí: 1 Svedberg = 1 S = sekundy. Jak od sebe oddělíme obě podjednotky a jak lze odstranit. p-hydroxymerkuribenzoát Větší podjednotka (c) je katalytická, ale není inhibována CTP. Složena ze tří řetězců po 34 kD se označuje c3. Regulační podjednotka (r) váže CTP. Složena ze dvou řetězců po 17 kD je označena r2. Nativní enzym je složen ze dvou katalytických trimerů a tří regulačních dimerů (2 c3 + 3 r2 = c6r6).")
8
Ultracentrifugační studie ATCasy
Ultracentrifugační studie ATCasy. A) Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym disocioval na regulační (r) a katalytické podjednotky (c).
 Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym disocioval na regulační (r) a katalytické podjednotky (c).")
9
Kvarterní struktura ATCasy. Vpravo je zjednodušený pohled
Kvarterní struktura ATCasy. Vpravo je zjednodušený pohled. Katalytické trimery jsou dva (druhý je zakrytý). Regulační dimery jsou tři. 2 c3 + 3 r2 = c6r6
. Regulační dimery jsou tři. 2 c3 + 3 r2 = c6r6.")
10
Strukturní podobnost mezi reakčním produktem ATCasy N-fosfonokarbamoylaspartátem a PALA (bisubstrátový analog reakčního meziproduktu). Krystalizace ACTasy za přítomnosti PALA - lokalizace aktivního místa.

11
Aktivní místo ATCasy-trimer
Aktivní místo ATCasy-trimer. V aktivním místě, které obsahuje hlavně vedlejší skupiny z jedné podjednotky, ale také část druhé podjednotky (zeleně), je vázán PALA.
, je vázán PALA.")
12
ATCasa existuje ve dvou konformacích: kompaktní , relativně inaktivní označovaná jako T (tense) a uvolněná (relaxed) označovaná R. PALA stabilizuje R stav. ATCasa splňuje kritéria symetrického modelu.

13
Cytidintrifosfát (CTP) stabilizuje T stav
Cytidintrifosfát (CTP) stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.
 stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.")
14
T a R stavy v rovnováze. I v situaci bez přítomnosti substrátu existuje ATCasa v rovnováze obou stavů. Bez substrátu převažuje T nad R stavem faktorem 200.

15
ATCase vykazuje sigmoidní kinetiku
ATCase vykazuje sigmoidní kinetiku. Základní podmínka allosterie proteinu: a) Kvartérní struktura (složen z podjednotek). b) Závislost rychlosti enzymové reakce na koncentraci substrátu je esovitá křivka – sigmoidní tvar.
 Kvartérní struktura (složen z podjednotek). b) Závislost rychlosti enzymové reakce na koncentraci substrátu je esovitá křivka – sigmoidní tvar.")
16
Tvorbu sigmoidní křivky lze interpretovat jako směs dvou Michaelis-Mentenovských enzymů, jeden s vysokou hodnotou KM odpovídající T stavu a druhý s nízkou hodnotou KM odpovídající R stavu. S růstem koncentrací substrátu se rovnováha posunuje od T k R stavu.

17
Vliv CTP na kinetiku ATCasy
Vliv CTP na kinetiku ATCasy. CTP je allosterický inhibitor, stabilizuje T stav. Znesnadňuje vstup substrátu do aktivního místa.

18
Vliv ATP na kinetiku ATCasy. ATP je alosterický aktivátor
Vliv ATP na kinetiku ATCasy. ATP je alosterický aktivátor. Stabilizuje stav R – usnadněný vstup substrátu do aktivního místa.

19
Působení substrátů na allosterické enzymy se označuje jako homotropní efekt
(z řečtiny „stejný“). Působení nesubstrátových molekul na allosterické enzymy jako např. CTP a ATP na ATCasu se označuje jako heterotropní efekt. Zkuste uvažovat proč působí ATP jako aktivátor ATCasy (fyziologická úvaha).
. Působení nesubstrátových molekul na allosterické enzymy jako např. CTP a ATP na ATCasu se označuje jako heterotropní efekt. Zkuste uvažovat proč působí ATP jako aktivátor ATCasy (fyziologická úvaha).")
20
Glukokinasa Glukokinasa (EC ), systematický název: ATP:D-glukosa 6-fosfotransferasa je enzym katalyzující fosforylaci glukosy na glukosa-6-fosfát. Glukokinasa je přítomna v buňkách jater, pankreatu, mozku, střevě lidí a mnoha obratlovců. V každém z těchto orgánů hraje významnou roli při regulaci metabolismu sacharidů. Působí jako glukosový sensor. Reaguje na změny hladin glukosy při hladovění nebo nasycení. Glukokinasa (GK) je hexokinasový izozym homologní s dalšími hexokinasami. GK je kódovaná odlišným genem, na rozdíl od hexokinas, má také odlišné kinetické vlastnosti a s tím související různé odlišné funkce. GK má, na rozdíl od ostatních hexokinas, nižší afinitu ke glukose a její aktivita je lokalizována jen v několika typech buněk.
, systematický název: ATP:D-glukosa 6-fosfotransferasa je enzym katalyzující fosforylaci glukosy na glukosa-6-fosfát. Glukokinasa je přítomna v buňkách jater, pankreatu, mozku, střevě lidí a mnoha obratlovců. V každém z těchto orgánů hraje významnou roli při regulaci metabolismu sacharidů. Působí jako glukosový sensor. Reaguje na změny hladin glukosy při hladovění nebo nasycení. Glukokinasa (GK) je hexokinasový izozym homologní s dalšími hexokinasami. GK je kódovaná odlišným genem, na rozdíl od hexokinas, má také odlišné kinetické vlastnosti. a s tím související různé odlišné funkce. GK má, na rozdíl od ostatních hexokinas, nižší afinitu ke glukose a její aktivita je lokalizována jen v několika typech buněk.")
21
Glukokinasa GK mění svoji strukturu v závislosti na koncentraci glukosy. Je saturována při koncentraci glukosy 8 mM/L. GK není inhibována produktem – glukosa-6-fosfátem !!!! GK vykazuje, ač monomer, kooperativitu – allosterické chování !!! GK je monomer 465 aminokyselin o molekulové hmotnosti 50kD. Na molekule enzymu jsou dvě vazebná místa. Jedno pro substráty (aktivní místo) vážící glukosu a MgATP a druhé, pravděpodobně, pro allosterický aktivátor-inhibitor, které dosud nebyly jednoznačně identifikovány. Ostatní hexokinasy jsou dimery. GK – aktivita může být zvyšována nebo redukována v minutových intervalech glukokinasovým regulačním proteinem(GKRP).
 vážící glukosu a MgATP a druhé, pravděpodobně, pro allosterický aktivátor-inhibitor, které dosud nebyly jednoznačně identifikovány. Ostatní hexokinasy jsou dimery. GK – aktivita může být zvyšována nebo redukována. v minutových intervalech glukokinasovým regulačním proteinem(GKRP).")
22
Glukokinasa s v aktivním místě vázanou glukosou

23
Dva modely allosterie. Symetrický model ( Jacques Monod, Jeffries Wyman a Jean-Pierre Changeux – podle autorů zkracovaný jako MWC model). Sekvenční model (Daniel Koshland).
.")
24
Conceptual insights – Chapter 10.

25
Kvantitativní popis modelu MWC
Kvantitativní popis modelu MWC. Frakční aktivita Yf je část aktivních míst s navázaným substrátem úměrná reakční rychlosti. a je poměr [ S ] / disociační konstantě S s enzymem ve stavu R; L je poměr koncentrací enzymu ve stavu T k enzymu ve stavu R. Vazba regulátorů ATP a CTP mění hodnotu L.

26
Symetrický (MWC) model – kvantitativní formulace.
Mějme enzym s n aktivními místy. Dva stavy enzymu v rovnováze: R – vysoká afinita k substrátu, T - nízká afinita k substrátu. Definujeme c jako poměr afinit obou forem enzymu k substrátu S, měřeno jako disociační konstanty.

27
Frakční aktivita Y, je frakce aktivních míst s navázanými substráty
Frakční aktivita Y, je frakce aktivních míst s navázanými substráty. Je přímo úměrná reakční rychlosti. a je poměr [ S ] ku disociační konstantě S v komplexu s enzymem v R stavu. L je poměr koncentrace enzymu v T stavu ku koncentraci enzymu v R stavu (KR). Vazba regulátorů jako je ATP nebo CTP na ATCasu mění hodnotu L a tím odezvu na koncentraci substrátu.
. Vazba regulátorů jako je ATP nebo CTP na ATCasu mění hodnotu L a tím odezvu na koncentraci substrátu.")
29
Příklad využití kvantitativního modelu s daty ATCasy.
U ATCasy se n = 6. Vynikajícího souhlasu s experimentálními daty se dosahuje při L = 200 a c = 0, 1. Z toho plyne, že za absence vázaného substrátu je posunuta rovnováha na stranu T faktorem 200 ( pouze 1 molekula z 200 je ve stavu R) a afinita R stavu k substrátu je 10x vyšší než ke stavu T. Když se substrát váže do každého aktivního místa, rovnováha se posouvá k R. Např.:Pokud je obsazena polovina aktivních míst substrátem(tři ze šesti) je poměr T/R 1 : 5. Znamená to, že téměř všechny molekuly jsou ve stavu R.
 a afinita R stavu k substrátu je 10x vyšší než ke stavu T. Když se substrát váže do každého aktivního místa, rovnováha se posouvá k R. Např.:Pokud je obsazena polovina aktivních míst substrátem(tři ze šesti) je poměr T/R 1 : 5. Znamená to, že téměř všechny molekuly jsou ve stavu R.")
30
Vliv CTP a ATP se řeší jednoduše změnou hodnoty L.
Při saturaci CTP se hodnota L zvyšuje na 1 250. To znamená, že je třeba daleko více substrátu k posunu rovnováhy do stavu R. Při saturaci ATP klesá hodnota L na 70.

31
SEKVENČNÍ MODEL Daniel KOSHLAND – postupná vazba ligandů (substrátů) na podjednotky proteinu vede k postupnému vzájemnému ovlivňování jednotlivých podjednotek. Možno vysvětlit i negativní kooperaci. U symetrického modelu (MWC) přechází celý enzym z jednoho stavu do druhého (T a R).
 na podjednotky proteinu vede k postupnému vzájemnému ovlivňování jednotlivých podjednotek. Možno vysvětlit i negativní kooperaci. U symetrického modelu (MWC) přechází celý enzym z jednoho stavu do druhého. (T a R).")
32
Jednoduchý sekvenční model tetramerního allosterického enzymu
Jednoduchý sekvenční model tetramerního allosterického enzymu. Postupnou vazbou substrátu na podjednotky se mění konformace okolních ze stavu T na R.

33
Max Perutz (1914 – 2002) a John Kendrew – Nobelova cena za chemii v roce 1962.
Max Perutz pracoval na rentgenové struktuře Hb více než 30 let na Univerzitě v Cambridge (UK). Struktura o rozlišení 2, 8 Å byla získána až v roce 1968. John Kendrew navazoval na M. Perutze, Objasnil strukturu velrybího myoglobinu – rentgenostrukturní analýzou, metodou isomorfní záměny. Těžký atom jako Hg2+ (bohatý na elektrony) se musí vázat na protein, tak, aby nedošlo ke změně struktury. Dochází k podstatné změně v intenzitě odrazu, ale ne v pozici elektronové hustoty.
. Struktura o rozlišení 2, 8 Å byla získána až v roce John Kendrew navazoval na M. Perutze, Objasnil strukturu velrybího myoglobinu – rentgenostrukturní analýzou, metodou isomorfní záměny. Těžký atom jako Hg2+ (bohatý na elektrony) se musí vázat na protein, tak, aby nedošlo. ke změně struktury. Dochází k podstatné změně v intenzitě odrazu, ale ne v pozici elektronové hustoty.")
34
Kvartérní struktura hemoglobinu – dva řetězce a a dva b tvoří pár ab dimerů – heterotetramer a2b2.

35
Kooperativní zvýšení transportu kyslíku u hemoglobinu
Kooperativní zvýšení transportu kyslíku u hemoglobinu. Kooperativita mezi vazebnými místy pro kyslík vede k zásobení tkání větším množstvím kyslíku (1, 7x) než by mohl nekooperativní protein. (pO2 je parciální tlak kyslíku).
 než by mohl nekooperativní protein. (pO2 je parciální tlak kyslíku).")
36
Jednotky tlaku 1 torr = 1 mm Hg ≈ 133,322 Pa
Pascal (značka Pa) je základní jednotka tlaku. Udává, jak velká síla (v Newtonech) působí na jednotkovou plochu (1 m2), tzn. je ekvivalentní N/m². 1 hPa = 100 Pa = 1 mb (milibar) 1 Pa = 1 N/m2 1 hPa = 100Pa 1 kPa = 1000 Pa 760 torrů ≈ 101,325 hPa 1 atm = Pa = 101,325 kPa = 0, MPa = 760 torr Parciální tlak plynu ve směsi je tlak, který by tento plyn vykazoval, pokud by byl v celém objemu sám.
 je základní jednotka tlaku. Udává, jak velká síla (v Newtonech) působí na jednotkovou plochu (1 m2), tzn. je ekvivalentní N/m². 1 hPa = 100 Pa = 1 mb (milibar) 1 Pa = 1 N/m2 1 hPa = 100Pa 1 kPa = 1000 Pa. 760 torrů ≈ 101,325 hPa. 1 atm = Pa = 101,325 kPa = 0, MPa = 760 torr. Parciální tlak plynu ve směsi je tlak, který by tento plyn vykazoval, pokud by byl v celém objemu sám.")
37
Krevní oběh – srdce a plíce
Krevní oběh – srdce a plíce. Formy Hb: HbO2 = oxyhemoglobin, deoxyhemoglobin, Mb = svalový myoglobin.

38
Karbonátanhydratasa EC 4.2.1.1 (metaloenzym – Zn2+) Zkratka CA.
 Zkratka CA.")
39
Aktivní místo živočišné a-CA (Zn2+ je vázán na tři imidazolové zbytky His)
")
40
Reakce katalyzované CA.
CO2 + H2O HCO H+ (za situace vysoké koncentrace CO2 v tkáních) Velmi rychlá reakce – 104 až 106 obratů za sekundu (číslo přeměny). Limitováno jen difůzí !!! Zpětná reakce: HCO H+ H2CO3 CO2 + H2O (v plících a nefronech ledvin za stavu nízké koncentrace CO2 , v rostlinách).
 Velmi rychlá reakce – 104 až 106 obratů za sekundu (číslo přeměny). Limitováno jen difůzí !!! Zpětná reakce: HCO3- + H+ H2CO3 CO2 + H2O (v plících a nefronech ledvin za stavu nízké koncentrace CO2 , v rostlinách).")
41
Funkce CA Primární funkcí CA u živočichů je udržování rovnováhy mezi CO2 a hydrgenuhličitanem za účelem udržování acidobazické rovnováhy v krvi a tkáních. Další funkcí je role CA při transportu CO2 z tkání. Existuje 14 různých izoforem CA u savců. Rostliny obsahují odlišnou formu nazvanou β-karbonátanhydratasa, která je z hlediska evoluce odlišný enzym, ale podílí se na stejné reakci a využívá atom zinku v aktivním místě. U rostlin je úkolem CA zvyšovat koncentraci CO2 v chloroplastech za účelem zvýšení rychlosti karboxylace katalyzované RuBisCem. Reakce fotosyntézy – RuBisCo využívá pouze CO2 a ne uhličitou kyselinu nebo hydrogenuhličitan.

42
Mechanismus působení CA
Prosthetická skupina Zn 2+ v aktivním místě je koordinována třemi His. Čtvrtá koordinační pozice je obsazena molekulou vody. Dochází k polarizaci vazby vodík – kyslík. Kyslík má negativní náboj a tím je vazba s vodíkem zeslabena. Čtyři His kolem vody jako substrátu váží proton, což vede k k uvolnění hydroxidové skupiny vázané na Zn. Příklad acidobazické katalýzy. Aktivní místo také obsahuje specifickou kapsu pro CO2 v těsné blízkosti hydroxidové skupiny. Na elektrony bohatý OH – se váže na oxid uhličity za tvorby hydrogenuhličitanu.

44
Pozice Fe2+ v deoxyhemoglobinu.

45
Vazba kyslíku vede ke strukturním změnám
Vazba kyslíku vede ke strukturním změnám. Atom Fe je vtažen po vazbě kyslíku do roviny hemu. S ním je vtažen i proximální His.

46
Konformační změny v hemoglobinu
Konformační změny v hemoglobinu. Pohyb Fe2+ po vazbě kyslíku vtáhne do kruhu proximální His, který jako součást a helixu ovlivní svým C koncem vztah mezi ab páry vedoucí k dalším strukturním změnám. Porovnáni se strukturou deoxy (šedá).
.")
47
Přechod ze stavu T do stavu R u hemoglobinu
Přechod ze stavu T do stavu R u hemoglobinu. Po vazbě kyslíku na pár a1 b1 se a1 b1 pár pootočí vůči druhému a2 b2 páru o 15o.

48
Posun mezi řetězci b2 a a1 hemoglobinu po vazbě kyslíku
Posun mezi řetězci b2 a a1 hemoglobinu po vazbě kyslíku. Stavy R vázán kyslík a T bez kyslíku.

49
Symetrický model allosterie
Symetrický model allosterie. V jedné kvarterní struktuře nemohou existovat vedle sebe formy T a R.

50
Sekvenční model allosterie
Sekvenční model allosterie. Ligand po vazbě na jednu podjednotku progresívně vyvolává konformační změny sousedních podjednotek.

51
Síť iontových párů v deoxyhemoglobinu
Síť iontových párů v deoxyhemoglobinu. C-koncové skupiny (Arg 141a a His 146b) v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.
 v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.")
52
Síť iontových párů v deoxyhemoglobinu
Síť iontových párů v deoxyhemoglobinu. C-koncové skupiny (Arg 141a a His 146b) v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.
 v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.")
53
Který z modelů je ten, který lze aplikovat na kooperativní vazbu kyslíku na hemoglobin (Hb) ?
Žádný z obou modelů nelze na Hb čistě aplikovat. Vyhovuje spíše kombinace obou modelů. Hb se třemi místy obsazenými kyslíkem splňuje kriteria sekvenčního modelu – nachází se v R stavu. Zbývající čtvrté vazebné místo má afinitu ke kyslíku 20x vyšší než plně deoxygenovaný Hb vážící svůj první kyslík. Kriteria sekvenčního modelu však nejsou splněna, protože Hb s kyslíkem navázaným pouze v jednom místě ze čtyř zůstává v T stavu. Tato struktura váže kyslík 3x pevněji než plně deoxygenovaný Hb – což odpovídá symetrickému (MWC) modelu. Závěr: Oba modely reprezentují idealizované limitní stavy, ke kterým se reálný systém blíží.
 modelu. Závěr: Oba modely reprezentují idealizované limitní stavy, ke kterým se reálný systém blíží.")
54
Describe and differentiate between the concerted model and the sequential model of allosteric regulation. How does hemoglobin do both? CONCERTED – MWC (allosteric) - connected enzyme subunits must exist in the same conformation - equilibrium (absense of substrate (or ligand)) favors the T-tense state (4 squares) vs. R-relaxed state (4 circles) - the binding of substrate to one subunits causes all remaining subunits to assume the R state (i.e.) thus enhancing their affinity for substrate SEQUENTIAL (allosteric) - subunits need not exist in same conformation - molecules of substrate bind via induced-fit protocol - conformational changes are not propagates to other subunits - substrate binding causes increased substrate affinity only Hemoglobin is a combination of both it is concerted: 3 sites occupied by oxygen associated with R state it is sequential: 1 site occupied by oxygen associated with T state
 - connected enzyme subunits must exist in the same conformation. - equilibrium (absense of substrate (or ligand)) favors the T-tense state (4 squares) vs. R-relaxed state (4 circles) - the binding of substrate to one subunits causes all remaining subunits to assume the R state (i.e.) thus enhancing their affinity for substrate. SEQUENTIAL (allosteric) - subunits need not exist in same conformation. - molecules of substrate bind via induced-fit protocol. - conformational changes are not propagates to other subunits. - substrate binding causes increased substrate affinity only. Hemoglobin is a combination of both. it is concerted: 3 sites occupied by oxygen associated with R state. it is sequential: 1 site occupied by oxygen associated with T state.")
57
Hillova rovnice, Hillův koeficient
In biochemistry, the binding of a ligand to a macromolecule is often enhanced if there are already other ligands present on the same macromolecule (this is known as Cooperative binding). The Hill coefficient, named for Archibald Vivian Hill, provides a way to quantify this effect. It describes the fraction of the macromolecule saturated by ligand as a function of the ligand concentration; it is used in determining the degree of cooperativeness of the ligand binding to the enzyme or receptor. It was originally formulated by Archibald Hill in 1910 to describe the sigmoidal O2 binding curve of hemoglobin. A coefficient of 1 indicates completely independent binding, regardless of how many additional ligands are already bound. Numbers greater than one indicate positive cooperativity, while numbers less than one indicate negative cooperativity. The Hill coefficient was originally devised to explain the cooperative binding of oxygen to hemoglobin (a system which has a Hill coefficient of 2.8-3). A.V. Hill (1886 – 1977) was an English physiologis, one of the founders of the diverse disciplines of biophysic and operations research. He shared the 1922 Nobel Prize in Physiolgy and Medicine “for his elucidation of the production of heat and mechanical work in muscles.
. The Hill coefficient, named for Archibald Vivian Hill, provides a way to quantify this effect. It describes the fraction of the macromolecule saturated by ligand as a function of the ligand concentration; it is used in determining the degree of cooperativeness of the ligand binding to the enzyme or receptor. It was originally formulated by Archibald Hill in 1910 to describe the sigmoidal O2 binding curve of hemoglobin. A coefficient of 1 indicates completely independent binding, regardless of how many additional ligands are already bound. Numbers greater than one indicate positive cooperativity, while numbers less than one indicate negative cooperativity. The Hill coefficient was originally devised to explain the cooperative binding of oxygen to hemoglobin (a system which has a Hill coefficient of 2.8-3). A.V. Hill (1886 – 1977) was an English physiologis, one of the founders of the diverse disciplines of biophysic and operations research. He shared the 1922 Nobel Prize in Physiolgy and Medicine for his elucidation of the production of heat and mechanical work in muscles.")
58
Taking the reciprocal of both sides, rearranging, inverting again, and then taking the logarithm on both sides of the equation leads to an alternative formulation of the Hill equation (θ = theta): θ - fraction of occupied sites where the ligand can bind to the active site of the receptor protein. [L] - ligand concentration Kd - Apparent dissociation constant derived from the law of mass action (equilibrium constant for dissociation) KA - ligand concentration producing half occupation (ligand concentration occupying half of the binding sites), that is also the microscopic dissociation constant. n - Hill coefficient, describing cooperativity (and many more, depending on the system, in the case of which the Hill equation is used)
 KA - ligand concentration producing half occupation (ligand concentration occupying half of the binding sites), that is also the microscopic dissociation constant. n - Hill coefficient, describing cooperativity (and many more, depending on the system, in the case of which the Hill equation is used)")
59
Hodnoty n a jejich interpretace
When appropriate, the value of the Hill constant describes the cooperativity of ligand binding in the following way: n > 1 - Positively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules increases. n < 1 - Negatively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules decreases. n = 1 - Noncooperative reaction: The affinity of the enzyme for a ligand molecule is not dependent on whether or not other ligand molecules are already bound. The Hill equation (as a relationship between the concentration of a compound adsorbing to binding sites and the fractional occupancy of the binding sites) is equivalent to the Langmuir equation.
 is equivalent to the Langmuir equation.")
60
Typický Hillův graf pro Hb. Y = frakce obsazených akt
Typický Hillův graf pro Hb. Y = frakce obsazených akt. míst; L – koncentrace ligandu (kyslík); kd = disociační konstanta kyslík – Hb; n = 2,8
; kd = disociační konstanta kyslík – Hb; n = 2,8.")
61
Leghemoglobin Leghemoglobin je hemoprotein analogický hemoglobinu u motýlokvětých rostlin fixujících dusík ve rhizobiích (dusík fixující baktérie) na kořenech. Leghemoglobin má také červenou barvu. Jedná se o symbiotický proces. Původně se mělo zato, že apoprotein leghemoglobinu syntetizuje rostlina a hem bakterie. Nejnovější poznatky ukazují, že hem je také syntetizován rostlinou. Leghemoglobin má vysokou afinitu ke kyslíku – asi 10 x vyšší než b-řetězec hemoglobinu (Km = 0, 01 mM).
 na kořenech. Leghemoglobin má také červenou barvu. Jedná se o symbiotický proces. Původně se mělo zato, že apoprotein leghemoglobinu syntetizuje rostlina a hem bakterie. Nejnovější poznatky ukazují, že hem je také syntetizován rostlinou. Leghemoglobin má vysokou afinitu ke kyslíku – asi 10 x vyšší než b-řetězec hemoglobinu. (Km = 0, 01 mM).")
62
Leghemoglobin U rostlin infikovaných Rhizobii by přítomnost kyslíku v kořenových nodulech silně redukovala funkci a aktivitu na kyslík citlivé nitrogenasy – enzymu, který fixuje atmosférický dusík. Leghemoglobin pufruje koncentarci volného kyslíku v cytoplasmě infikovaných buněk a brání vstupu kyslíku na nitrogenasu. . Poslední studie ukazují, že leghemoglobin skladuje jen tolik kyslíku kolik je třeba k podpoře respirace v nodulech na několik sekund. Jeho funkcí je tedy také transport kyslíku do respirujících symbiotických bakteriálních buněk, analogicky jak tak činí hemoglobin transportující kyslík do respirujících tkání.

63
Vliv 2,3-bisfosfoglycerátu (2,3-BPG) na afinitu kyslíku k hemoglobinu.
Bez přítomnosti 2,3-BPG je afinita hemoglobinu ke kyslíku podstatně vyšší a uvolňuje se jen 8 % přenášeného kyslíku do tkání. 2,3-DPG je přítomen v erythrocytech v koncentraci 2 mM (stejná jako konc. hemoglobinu).
.")
64
Způsob vazby 2,3-BPG (náboj-5) na lidský hemoglobin
Způsob vazby 2,3-BPG (náboj-5) na lidský hemoglobin. Váže se do centrální dutiny deoxyhemoglobinu na osm kladně nabitých skupin obou b řetězců.
 na lidský hemoglobin. Váže se do centrální dutiny deoxyhemoglobinu na osm kladně nabitých skupin obou b řetězců.")
65
Proč 2,3-BPG tak silně ovlivňuje afinitu hemoglobinu ke kyslíku ?
2,3-BPG se váže přednostně na deoxyhemoglobin, který stabilizuje. Deoxyhemoglobin je ve stavu T. Přechod do stavu R se děje jen po oddělení 2,3-BPG ! Fyziologický důsledek: Plodový (fetální) hemoglobin se liší od hemoglobinu dospělého člověka. Je to tetramer obsahující dva řetězce a a dva g !! Řetězce g jsou ze 72 % identické s řetězci b u hemoglobinu dospělých. Tato změna však znamená, že řetězce g mají o dva pozitivní náboje méně než b a tím je podstatně snížena afinita fetálního hemoglobinu k 2,3-BPG a tím zvýšena afinita ke kyslíku ve srovnání s hemoglobinem mateřským. To umožňuje bezproblémový přenos kyslíky z matky na plod.
 hemoglobin se liší od hemoglobinu dospělého člověka. Je to tetramer obsahující dva řetězce a a dva g !! Řetězce g jsou ze 72 % identické s řetězci b u hemoglobinu dospělých. Tato změna však znamená, že řetězce g mají o dva pozitivní náboje méně než. b a tím je podstatně snížena afinita fetálního hemoglobinu. k 2,3-BPG a tím zvýšena afinita ke kyslíku ve srovnání s hemoglobinem mateřským. To umožňuje bezproblémový přenos kyslíky z matky na plod.")
66
Inositol – inositolhexafosfát(IP6), fytinová kyselina.
, fytinová kyselina.")
67
Afinita ke kyslíku u fetálních erythrocytů.

68
Bohrův efekt. Rychle metabolizující tkáně, jako svalstvo, mají velké požadavky na kyslík a produkují velké množství H+ a CO2. Jak H+, tak CO2 jsou heterotropní efektory hemoglobinu zvyšující uvolňování kyslíku. Afinita hemoglobinu ke kyslíku klesá při vstupu hemoglobinu do oblastí s nižším pH. Transport z plic (pH 7, 4), parciální tlak kyslíku 100 torrů do aktivního svalu s pH 7, 2 a parciálního tlaku kyslíku 20 torrů vede k uvolnění 77 % celkové přenášené kapacity. Pokud by nedošlo k poklesu pH, tak by se uvolnilo pouze 66 %. Ve tkáních, kde je vyšší koncentrace CO2 klesá afinita hemoglobinu ke kyslíku. Za přítomnosti CO2 o parciálním tlaku 40 torrů se uvolní z hemoglobinu 90 % přenášeného kyslíku. Vliv H+ a CO2 na regulaci vazby kyslíku na hemoglobin se nazývá Bohrův efekt. Podle Christiana Bohra, který jev popsal v roce 1904.
, parciální tlak kyslíku 100 torrů do aktivního svalu s pH 7, 2 a parciálního tlaku kyslíku 20 torrů vede k uvolnění 77 % celkové přenášené kapacity. Pokud by nedošlo k poklesu pH, tak by se uvolnilo pouze 66 %. Ve tkáních, kde je vyšší koncentrace CO2 klesá afinita hemoglobinu ke kyslíku. Za přítomnosti CO2 o parciálním tlaku 40 torrů se uvolní z hemoglobinu 90 % přenášeného kyslíku. Vliv H+ a CO2 na regulaci vazby kyslíku na hemoglobin se nazývá Bohrův efekt. Podle Christiana Bohra, který jev popsal v roce")
69
Vliv pH a koncentrace CO2 na afinitu kyslíku k hemoglobinu.

70
Chemický základ Bohrova efektu.
V deoxyhemoglobinu tvoří tři aminokyseliny dvě iontové vazby stabilizující T stav. Tvorba jedné z vazeb závisí na přítomnosti dalšího protonu na His b 146. To umožňuje vazbu na Asp 94.

71
CO2 stabilizuje deoxyhemoglobin reakcí s terminální aminoskupinou za tvorby karbamátu. Změna náboje vede k pevnější vazbě mezi ab dimery a tím ke stabilizaci T stavu.

72
Hemoglobin s navázanými CO2 a H+ vstupuje do plic, kde uvolní CO2 a H+ a naváže kyslík.
Hemoglobin se však podílí pouze 14 % na celkovém transportu oxidu uhličitého a protonů. Jak CO2, tak H+ jsou transportovány krví jako hydrogenuhličitan (HCO3-), který se tvoří spontánně nebo enzymově karbonátanhydrasou.
, který se tvoří spontánně nebo enzymově karbonátanhydrasou.")
73
Glykovaný hemoglobin (HbA1)
Neenzymová glykace na terminální NH2 skupině (Val) β-řetězce Glykovaná frakce asi 5% celkového množství hemoglobinu (úměrná koncentraci glukózy v krvi) Měřením hladiny HbA1 lze získat informace o průběhu diabetes mellitus (odráží hladinu glukózy za posledních několik týdnů); Cukr CHO + NH2 CH2 Protein Cukr CH N CH2 Protein Cukr CH2 NH CH2 Protein Schiffova base Amadoriho přesmyk Glykovaný protein
 β-řetězce. Glykovaná frakce asi 5% celkového množství hemoglobinu (úměrná koncentraci glukózy v krvi) Měřením hladiny HbA1 lze získat informace o průběhu diabetes mellitus (odráží hladinu glukózy za posledních několik týdnů); Cukr CHO + NH2 CH2 Protein. Cukr CH N CH2 Protein. Cukr CH2 NH CH2 Protein. Schiffova base. Amadoriho přesmyk. Glykovaný protein.")
74
Přepínání hemoglobinu

75
MYOGLOBIN

77
Vazba kyslíku na myoglobin – myoglobin je zpoloviny nasycen kyslíkem při tlaku kyslíku (pO2) 2, 8 torru.
 2, 8 torru.")
78
Fyziologická role myoglobinu.
Rolí myoglobinu je usnadňovat difůzi kyslíku ve svalech při intenzívní svalové činnosti. Rychlost difůze kyslíku z krevních kapilár do tkání je limitována jeho nízkou rozpustností ve vodě (10-4 M v krvi). Myoglobin zvyšuje efekt rozpustnosti kyslíku ve svalových buňkách. Myoglobin jako „sklad“ kyslíku je důležitý u mořských savců jako je např. velryba, jejichž koncentrace myoglobinu ve svalech je 10x vyšší než u suchozemských savců. Myši u kterých byl vyřazen („knocked out“) gen pro myoglobin, mají světlejší svalstvo, ale jinak jsou normální. Myoglobin není pro život suchozemských savců nezbytný. V mozku byl nedávno popsán protein charakteru myoglobinu nazvaný neuroglobin. Funkcí je podpora koncentrace O2 v nervových tkáních, které jsou metabolicky velmi aktivní. Mozek reprezentuje asi 2% hmotnosti lidského těla, ale spotřebovává 20% získaného kyslíku.
. Myoglobin zvyšuje efekt rozpustnosti kyslíku ve svalových buňkách. Myoglobin jako „sklad kyslíku je důležitý u mořských savců jako je např. velryba, jejichž koncentrace myoglobinu ve svalech je 10x vyšší než u suchozemských savců. Myši u kterých byl vyřazen („knocked out ) gen pro myoglobin, mají světlejší svalstvo, ale jinak jsou normální. Myoglobin není pro život suchozemských savců nezbytný. V mozku byl nedávno popsán protein charakteru myoglobinu nazvaný neuroglobin. Funkcí je podpora koncentrace O2 v nervových tkáních, které jsou metabolicky velmi aktivní. Mozek reprezentuje asi 2% hmotnosti lidského těla, ale spotřebovává 20% získaného kyslíku.")
79
Adaptace na výšku. Atmosférický tlak klesá s výškou. Tlak kyslíku ve výšce m je 110 torrů, tj. 70 % hodnoty tlaku na hladině moře. Tlak kyslíku pod 85 torrů je neslučitelný se životem. Adaptace na výšku je spojena se zvýšením množství Hb v erythrocytech a zvýšením počtu erythrocytů. Tento komplexní proces obvykle zahrnuje několik týdnů. Mnohdy stačí pouze jeden den k dosažení dostatečné adaptace. Tento efekt spočívá v rapidním zvýšení množství BPG syntetizovaného v erythrocytech (obvykle ze 4 mM na 8 mM; BPG nemůže procházet membránou erythrocytu). Zvýšená koncentrace BPG způsobuje, že se křivka saturace Hb kyslíkem posunuje oproti křivce na hladině moře do pozice znamenající nižší afinitu !! Na hladině moře je rozdíl pO2 mezi tepennou a žilní krví 70 torrů a Hb uvolňuje 38 % vázaného kyslíku. Když obsah vázaného kyslíku v tepnách klesne na 55 torrů, to je ve výšce m, uvolňuje Hb jen 30 % vázaného kyslíku. Adaptace na výšku (pokles vázaného kyslíku v plících, zvýšení množství kyslíku uvolněného v tkáních) znamená, že Hb přenáší prakticky normální množství kyslíku.
. Zvýšená koncentrace BPG způsobuje, že se křivka saturace Hb kyslíkem posunuje oproti křivce na hladině moře do pozice znamenající nižší afinitu !! Na hladině moře je rozdíl pO2 mezi tepennou a žilní krví 70 torrů a Hb uvolňuje 38 % vázaného kyslíku. Když obsah vázaného kyslíku v tepnách klesne na 55 torrů, to je ve výšce m, uvolňuje Hb jen 30 % vázaného kyslíku. Adaptace na výšku (pokles vázaného kyslíku v plících, zvýšení množství kyslíku uvolněného v tkáních) znamená, že Hb přenáší prakticky normální množství kyslíku.")
80
Koncentrace BPG se zvyšuje v erythrocytech mnohem rychleji než proběhne syntéza Hb (erythrocyty nemají jádro a proto nemohou syntetizovat proteiny). Hladina BPG je mnohem citlivější regulace přenosu kyslíku než změna respirační rychlosti. Hyperventilace, další rychlá odezva na výšku, vede k respirační alkalose. Hyperventilace urychluje vydechování CO2 a způsobuje respirační alkalosu. Alkalosa (pH větší než 7, 6) a acidosa (pH menší než 7, 1) jsou následky nerovnováhy krevních ústojů. Čím je způsobována acidosa a jak se dá potlačit ?? Alkalosa se dá potlačit NaCl nebo KCl (proč?) nebo vdechováním vzduchu obohaceneého CO2.
 a acidosa (pH menší než 7, 1) jsou následky nerovnováhy krevních ústojů. Čím je způsobována acidosa a jak se dá potlačit Alkalosa se dá potlačit NaCl nebo KCl (proč ) nebo vdechováním vzduchu obohaceneého CO2.")
81
Obyvatelé And a Himalájí mají vysokou kapacitu plic, vysokou hladinu Hb a často také větší pravou srdeční komoru což znamená zvýšení objemu čerpané krve. Na rozdíl od adaptace lidí na výšku, většina savců žijících ve vysokých výškách (lamy), má geneticky pozměněný Hb vyznačující se vyšší afinitou ke kyslíku. K adaptaci na výšku vedou dvě cesty: zvýšení i snížení p50 Hb.
, má geneticky pozměněný Hb vyznačující se vyšší afinitou ke kyslíku. K adaptaci na výšku vedou dvě cesty: zvýšení i snížení p50 Hb.")
83
Abnormální hemoglobiny

84
Methemoglobin Také ferrihemoglobin, zkr. metHb, hemoglobin obsahující trojmocné železo (Fe3+ ), není schopen přenášet kyslík. V erythrocytech vzniká za fyziologických podmínek v malém množství přímou oxidací železa kyslíkem; je zde enzymově zpětně redukován na Fe2+ formu. Větší množství metHb vzniká následkem otrav (acetanilid, chlornany, dusitany atd.) nebo vrozených poruch syntézy hemoglobinu. Může se patologicky objevovat v krevní plasmě (methemoglobinemie) nebo v moči (methemoglobinurie). Způsobuje modrání kůže, obecně, sliznic (cyanóza). Talasemie - disproporce tvorby řetězců a a b hemoglobinu. Některé vzácnější formy hereditárních anémií vyskytující se v dospělé populaci v ČR a SR – β−talasemie a nestabilní hemoglobinové varianty V. Divoký, S. Walczysková, D. Pospíšilová, M. Priwitzerová, S. Takáčová, I. Kostelecká, M. Divoká, Š. Rožmanová, M. Jarošová, J. Čermák, K. Indrák a Česko-slovenská kooperativní skupina pro diagnostiku hemoglobinopatií
, není schopen přenášet kyslík. V erythrocytech vzniká za fyziologických podmínek v malém množství přímou oxidací železa kyslíkem; je zde enzymově zpětně redukován na Fe2+ formu. Větší množství metHb vzniká následkem otrav (acetanilid, chlornany, dusitany atd.) nebo vrozených poruch syntézy hemoglobinu. Může se patologicky objevovat v krevní plasmě (methemoglobinemie) nebo v moči (methemoglobinurie). Způsobuje modrání kůže, obecně, sliznic (cyanóza). Talasemie - disproporce tvorby řetězců a a b hemoglobinu. Některé vzácnější formy hereditárních anémií vyskytující se v dospělé populaci v ČR a SR – β−talasemie a nestabilní hemoglobinové varianty. V. Divoký, S. Walczysková, D. Pospíšilová, M. Priwitzerová, S. Takáčová, I. Kostelecká, M. Divoká, Š. Rožmanová, M. Jarošová, J. Čermák, K. Indrák a Česko-slovenská kooperativní skupina pro diagnostiku hemoglobinopatií.")
85
2. Mnohočetné formy enzymů - isozymy (isoenzymy).
L- Laktátdehydrogenasa (EC ; (S)-laktát:NAD+ oxidoreduktasa ) -Isoenzymy H a M -Interakce isoenzymů LDH se substráty a inhibitory. -Isoenzymy LDH v různých fázích vývoje organismu a v různých orgánech Hexokinasa (EC ); ATP:D-hexosa-6-fosfotransferasa
-laktát:NAD+ oxidoreduktasa ) -Isoenzymy H a M. -Interakce isoenzymů LDH se substráty a inhibitory. -Isoenzymy LDH v různých fázích vývoje organismu. a v různých orgánech. Hexokinasa (EC ); ATP:D-hexosa-6-fosfotransferasa.")
86
ISOENZYMY – způsob regulace v různých tkáních a za různých vývojových stádií.
Isozymy nebo isoenzymy jsou enzymy lišící se sekvencí a složením aminokyselin katalyzující stejnou reakci. Obvykle mají různé kinetické parametry, jako Km, nebo různé aktivátory a inhibitory. Isoenzymy od sebe obvykle odlišíme dle jejich různé elektroforetické pohyblivosti. Jako typický příklad lze uvést isoenzymy laktátdehydrogenasy (LDH). LDH se uplatňuje při anaerobní metabolismu glukosy a syntéze glukosy (glukoneogeneze). U člověka rozeznáváme dva isoenzymové polypeptidové řetězce: H isoenzym – vysoký obsah v srdečním svalu a M isoenzym – skeletální svaly. Řetězce jsou ze 75 % shodné. Funkční enzym je tetramer.
. LDH se uplatňuje při anaerobní metabolismu glukosy a syntéze glukosy (glukoneogeneze). U člověka rozeznáváme dva isoenzymové polypeptidové řetězce: H isoenzym – vysoký obsah v srdečním svalu a. M isoenzym – skeletální svaly. Řetězce jsou ze 75 % shodné. Funkční enzym je tetramer.")
87
Lokalizace isoenzymů LDH
LDH-1 (H4) - srdce LDH-2 (H3M1) – retikuloendothel (Retikuloendotel, roztroušené buňky v různých oblastech těla, které jsou schopné zneškodnit choroboplodné látky. LDH-3 (H2M2) - plíce LDH-4 (H1M3) – ledviny, placenta a pankreas LDH-5 (M4) – svaly, játra
 - srdce LDH-2 (H3M1) – retikuloendothel. (Retikuloendotel, roztroušené buňky v různých oblastech těla, které jsou schopné zneškodnit choroboplodné látky. LDH-3 (H2M2) - plíce. LDH-4 (H1M3) – ledviny, placenta a pankreas. LDH-5 (M4) – svaly, játra")
88
Lidská LDH M4 (EC )
")
89
H4 isozym je hojný v srdečním svalu a má větší afinitu k substrátům než M4 isoenzym.
Vysoká hladina pyruvátu allostericky inhibuje H4 a ne M4. Proč ?? Další kombinace, jako je H3M, mají vlastnosti mezi H4 a M4. M4 isozym je optimální za anaerobních podmínek, H4, za aerobních. Přítomnost různých kombinací isoenzymů se mění s vývojem, např. srdce. Přítomnost různých isozymů v krvi je indikátorem poškození tkáně a tento fakt se využívá k diagnostickým účelům. Např. zvyšující se hladina H3M v krevním séru je indikací infarktu myokardu.

90
Isozymy LDH – krysí srdeční sval.
Krysí srdeční LDH isoenzym. Profil se mění s vývojem. H isoenzymy jsou modré kroužky, M jsou červené čtverce.

91
Obsah LDH isozymů v různých tkáních krysy.

92
Troponin = komplex tří regulačních proteinů
Troponin = komplex tří regulačních proteinů. Spojení se svalovou funkcí skeletu a srdce. Není přítomen v hladkém svalstvu.

93
Stanovení AST – EC 2.6.1.1. (aspartátaminotransferasay) v krevním séru
2-oxoglutarát + L-Asp = oxaloacetát + L-Glu Oxaloactát v silně kyselém prostředí dekarboxyluje na pyruvát. Pyruvát + 2,4-dinitrofenylhydrazin = 2,4-dinitrofenylhydrazon pyruvátu Hydrazon v alkalickém prostředí l max = 510 nm.

94
Vysoká hladina pyruvátu allostericky inhibuje H4 a ne M4.
Srdeční sval je aerobní, to znamená, že veškerý pyruvát je pyruvátdehydrogenasovým komplexem převáděn na acetyl-CoA. Bylo by nelogické převádět pyruvát na L-laktát !!! Sval pracuje převážně anaerobně a pyruvát vzniklý glykolýzou je za této situace převáděn na L-laktát, který je transportován Coriho cyklem do jater !! Pokud by nadbytek pyruvátu blokoval LDH, nedostávalo by se energie pro svalovou práci !!

95
Hexokinasa Hexokinasa (EC 2.7.1.1); ATP:D-hexosa-6-fosfotransferasa
Glukokinasa (EC ) ATP + D-hexosa = ADP + D-hexosa-6-fosfát Hexokinasa je enzym katalyzující fosforylaci šestiuhlíkatých sacharidů za tvorby hexosafosfátů. Hexokinasa se nachází u všech organismů počínaje bakteriemi, v kvasinkách, rostlinách až po člověka a obratlovce. Metabolické dráhy s účastí hexokinasy: Metabolismus aminosacharidů, metabolismus fruktosy a manosy, metabolismus galaktosy, glykolýza a glukoneogeneze, metabolismus škrobu a sacharosy, biosyntéza streptomycinu. U různých organismů existuje řada isoforem – isoenzymů, které mají řadu různých funkcí. Většina bakteriálních hexokinas má velikost cca 50 kD.
 ATP + D-hexosa = ADP + D-hexosa-6-fosfát. Hexokinasa je enzym katalyzující fosforylaci šestiuhlíkatých sacharidů za tvorby hexosafosfátů. Hexokinasa se nachází u všech organismů počínaje bakteriemi, v kvasinkách, rostlinách až po člověka. a obratlovce. Metabolické dráhy s účastí hexokinasy: Metabolismus aminosacharidů, metabolismus fruktosy. a manosy, metabolismus galaktosy, glykolýza. a glukoneogeneze, metabolismus škrobu a sacharosy, biosyntéza streptomycinu. U různých organismů existuje řada isoforem – isoenzymů, které mají řadu různých funkcí. Většina bakteriálních hexokinas má velikost cca 50 kD.")
96
Streptomycin je druh antibiotia patřící do skupiny aminoglykosidů
Streptomycin je druh antibiotia patřící do skupiny aminoglykosidů. Streptomycin se váže na ribosom bakterie a způsobuje, že jsou vkládány nesprávné aminokyseliny do buněčné stěny bakterie. Užívá se zejména jako antituberkulotikum - pro léčbu TBC. Získává se z plísně (aktinomyceta) Streptomyces griseus a používá se pro něj zkratka STR či STM.
 Streptomyces griseus a používá se pro něj zkratka STR či STM.")
97
Složení streptomycinu.
Streptomycin se skládá z aminocyklitolu (streptidin). 6-deoxyhexosy (streptosa) a N-methyl-L-glukosaminů, které se tvoří nezávislou biosyntetickou dráhou. Všechny tyto složky mají jako prekurzor D-glukosu.
. 6-deoxyhexosy (streptosa) a N-methyl-L-glukosaminů, které se tvoří nezávislou biosyntetickou dráhou. Všechny tyto složky mají jako prekurzor D-glukosu.")
98
Mnohobuněčné organismy, jako jsou rostliny a živočichové, mají často více hexosakinasových isoenzymů. Většina z nich má velikost 100 kD a je složena ze dvou podjednotek o 50 kD. Existují čtyři savčí hexokinasové isoenzymy lišící se v subcelulární lokalizaci, kinetice, s ohledem na různé substráty, reakční podmínky a fyziologickou funkci. Jsou označovány jako : Hexokinasa I, II, III a IV resp. A, B, C a D. Hexokinasy I, II a III. Hexokinasy I, II a III jsou charakteristické nízkou hodnotou Km. Mají vysokou afinitu ke glukose už při koncentracích pod 1mM. Hexokinasy I a II splňují kinetiku Michaelise a Mentenové při fyziologických koncentracích substrátů. Všechny tři jsou silně inhibovány produktem, kterým je glukosa-6-fosfát. Jejich relativní molekulová hmotnost je kolem 100 kD a jsou složeny ze dvou stejných podjednotek. Jen hexokinasa II má v obou podjednotkách funkční aktivní místa.

99
Hexokinasa I se nachází v savčích tkáních a je považována „provozní enzym“, neovlivňovaný většinou fyziologických, hormonálních a metabolických změn. Hexokinasa III je inhibována nadbytkem glukosy (inhibice substrátem). Hexokinasa IV (glukokinasa); (EC ) Savčí hexokinasa IV má unikátní charakteristické funkce ve srovnání se třemi předchozími. Fosforylace probíhá na subcelulární úrovni za situace, kdy je translokována mezi cytoplsmou a jádrem, např. v jaterních buňkách. K fosforylaci glukosy dochází při její vysoké koncentraci. Km pro glukosu je 100 krát vyšší než u hexokinas I, II a III. Glukokinasa je monomer (50 kD), vykazující pozitivní kooperativitu s glukosou a není allostericky inhibována produktem, glukosa-6-fosfátem.
. Hexokinasa IV (glukokinasa); (EC ) Savčí hexokinasa IV má unikátní charakteristické funkce. ve srovnání se třemi předchozími. Fosforylace probíhá na subcelulární úrovni za situace, kdy je translokována mezi cytoplsmou a jádrem, např. v jaterních buňkách. K fosforylaci glukosy dochází při její vysoké koncentraci. Km pro glukosu je 100 krát vyšší než u hexokinas I, II a III. Glukokinasa je monomer (50 kD), vykazující pozitivní kooperativitu s glukosou a není allostericky inhibována produktem, glukosa-6-fosfátem.")
100
Glukokinasa se nachází v játrech, pankreatu, hypothalmu, tenkém střevě a v některých endokrinních buňkách. Hraje významnou roli při regulaci sacharidového metabolismu. V beta buňkách Langerhansových ostrůvků pankreatu vykonává funkci glukosového senzoru pro uvolňování insulinu. Obdobně kontroluje uvolňování glukagonu z alfa buněk. V jaterních hepatocytech vede její funkce k syntéze glykogenu. Hexokinasa jako součást glykolýzy. Prvním stupněm glykolýzy je hexokinasová fosforylace glukosy. Když už se glukosa dostane do buňky, je nutné ji fosforylovat, kromě jiného, proto, aby nedifundovala z buňky ven. Glukosafosfáty neprochází buněčnou membránou.

101
Asociace hexokinasy s mitochondriemi.
Hexokinasy I, II a III se fyzikálními silami udržují na vnější membráně mitochondrií (porinový kanálek). To umožňuje, aby docházelo k fosforylaci hexos bezprostředně po tvorbě ATP mitochondriemi. Mitochondriální hexokinasy jsou silně zvýšeny u rychle rostoucích maligních buněk – dokonce 200 krát oproti normálním buňkám. Hexokinasa takto vázaná na mitochondrie je pokládána za urychlovače extrémně vysokých glykolytických procesů probíhajících v tumorových buňkách. Popsáno jako Warburgův efekt Otto Warburgem v roce 1930.
. To umožňuje, aby docházelo k fosforylaci hexos bezprostředně po tvorbě ATP mitochondriemi. Mitochondriální hexokinasy jsou silně zvýšeny. u rychle rostoucích maligních buněk – dokonce. 200 krát oproti normálním buňkám. Hexokinasa takto vázaná na mitochondrie je pokládána za urychlovače extrémně vysokých glykolytických procesů probíhajících v tumorových buňkách. Popsáno jako Warburgův efekt Otto Warburgem v roce")
102
3. Regulace enzymové aktivity kovalentní modifikací.
Nejčastější jsou fosforylace a defosforylace Dalším způsobem modifikace je např. acetylace. Acetylovány jsou histony (pomocné proteiny obalující DNA v chromosomech a při regulaci genů). Vysoce acetylované histony spojené s geny jsou aktivně přepisovány. Enzymy jsou acetyltransferasa a deacetylasa a jsou regulovány fosforylací. Modifikace není vždy reversibilní. Např. připojení ubiquitinu k proteinu.
. Vysoce acetylované histony spojené s geny jsou aktivně přepisovány. Enzymy jsou acetyltransferasa a deacetylasa a jsou regulovány fosforylací. Modifikace není vždy reversibilní. Např. připojení ubiquitinu k proteinu.")
103
Kovalentní modifikace proteinů – acetylace vedlejšího řetězce Lys

105
Vysvětlivky k tabulce: Common covalent modification of protein activity
Ras = onkogen – rat sarcoma viruses. Src = onkogen – Rous sarcoma viruses. Myristoyl-CoA = CH3 – (CH2)12 – CO-CoA Farnesylpyrofosfát (3 x prenyl, prenylace) –
12 – CO-CoA. Farnesylpyrofosfát (3 x prenyl, prenylace) –")
106
Fosforylace je vysoce účinný způsob modifikace regulující aktivitu cílového proteinu.

108
Pyruvátdehydrogenasakinasa (PDK) EC 2.7.11.2
PDK inaktivuje pyruvátdehydrogenasu fosforylací za účasti ATP. PDK se podílí na regulaci pyruvátdehdrogenasového komplexu jehož je PD prvním enzymem. PDK a pyruvátdehydrogenasový komplex jsou lokalizovány v matrix mitochondrií u eukaryot. Komplex převádí pyruvát, jako produkt glykolýzy vzniklý v cytoplasmě, na acetyl CoA., který je posléze oxidován v citrátovém cyklu za tvorby energie. PDK snižuje oxidaci pyruvátu v mitochondrii a zvyšuje převod pyruvátu na laktát v cytoplasmě. Opačné působení PDK- defosforylace – a aktivace pyruvátdehdrogenázového komplexu je katalyzováno fofoproteinfosfatasou nazývanou pyruvátdehydrogenasafosfatasa.

109
Regulace pyruvátdehydrogenasového komplexu

110
Pyruvátdehdrogenasakinasa (PDK) EC 2.7.11.2
PDK je stimulována ATP, NADH a acetyl CoA. PDK je inhibována ADP, NAD+ CoA-SH a pyruvátem. PDK je také inhibována farmakem dichloroacetátem, který je sledován jako prostředek k léčení několika metabolických onemocnění – zvláště proti rakovině. PDK má čtyři isozymy – PDK1 až PDK4.

111
Složení pyruvátdehydrogenasového komplexu:
Pyruvátdehydrogenasa (E1) Dihydrolipoyltransacetylasa (E2) Dihydrolipoyldehydrogenasa (E3) Např. komplex E. coli je kD proteinový komplex. Mitochondriální komplex je kD protein, obsahující 20 E2 trimerů obklopených 30 E1 heterotetramery a 12 E3 dimerů. Pyruvátdehydrogenasový komplex katalyzuje sekvencí tří reakcí, sumárně: Pyruvát + CoA + NAD+ → acetyl CoA + CO2 + NADH Komplex využívá pěti různých koenzymů: Thiaminpyrofosfát (TPP), koenzym A (CoA SH), NAD+, FAD a lipoamid. Pyruvátdehydrogenasový komplex, enzymové složení Pyruvátdehydrogenasový komplex, sumární reakce Pyruvátdehydrogenasový komplex, koenzymy 111 111
 Dihydrolipoyltransacetylasa (E2) Dihydrolipoyldehydrogenasa (E3) Např. komplex E. coli je kD proteinový komplex. Mitochondriální komplex je kD protein, obsahující 20 E2 trimerů obklopených 30 E1 heterotetramery a 12 E3 dimerů. Pyruvátdehydrogenasový komplex katalyzuje sekvencí tří reakcí, sumárně: Pyruvát + CoA + NAD+ → acetyl CoA + CO2 + NADH. Komplex využívá pěti různých koenzymů: Thiaminpyrofosfát (TPP), koenzym A (CoA SH), NAD+, FAD a lipoamid. Pyruvátdehydrogenasový komplex, enzymové složení. Pyruvátdehydrogenasový komplex, sumární reakce. Pyruvátdehydrogenasový komplex, koenzymy")
112
Thiaminpyrofosfát – TPP, také thiamindifosfát TDP
Thiaminpyrofosfát – TPP, také thiamindifosfát TDP. Váže se pevně, ale nekovalentně na pyruvátdekarboxylasu. Prekurzorem je vitamin B1 – thiamin.

113
Nekovalentní vazba TPP na pyruvátdekarboxylasu.

114
Pyruvátdehydrogenasa (E1) Pyruvát dekarboxyluje za tvorby hydroxyethyl-TPP meziproduktu.
Pyruvátdehydrogenasový komplex, pyruvátdehydrogenasa 114 114

115
Lipoamid a dihydrolipoamid
Lipoamid a dihydrolipoamid. Lipoová kyselina je vázána na E2 amidovou vazbou přes e-aminoskupinu Lys. Pyruvátdehydrogenasový komplex, lipoamid Pyruvátdehydrogenasový komplex, dihydrolipoamid 115 115

116
Hydroxyethylová skupina je přenesena na dihydrolipoyltransacetylasu (E2). Hydroxyethylový karbanion je současně oxidován na acetyl a lipoamid redukován na disulfid. Pyruvátdehydrogenasový komplex, dihydrolipoyltransacetylasa 116 116

117
E2 (Dihydrolipoyltransacetylasa) poté katalyzuje transesterifikací, při které se acetyl přenese na CoA za tvorby acetyl-CoA. Pyruvátdehydrogenasový komplex, transesterifikace 117 117

118
Regenerace lipoamidu na E2
Regenerace lipoamidu na E2. Reoxidace probíhá přes kovalentně vázaný FAD. Pyruvátdehydrogenasový komplex, regenerace lipamidu 118 118

119
Reoxidace redukovaného E3 (Dihydrolipoyldehydrogenasa )
Reoxidace redukovaného E3 (Dihydrolipoyldehydrogenasa ). Elektrony z FADH2 se přenáší na NAD+ za tvorby NADH. FAD slouží spíše jako vodič elektronů !!! Pyruvátdehydrogenasový komplex, reoxidace dihydrolipoyldehydrogenasy (E3) 119 119
. Elektrony z FADH2 se přenáší na NAD+ za tvorby NADH. FAD slouží spíše jako vodič elektronů !!! Pyruvátdehydrogenasový komplex, reoxidace dihydrolipoyldehydrogenasy (E3)")
120
Aktivní místo dihydrolipoamiddehydrogenasy.
FAD Cys43 Cys48 NAD+ Dihydrolipoyldehydrogenasa (E3), dihydrolipoamiddehydrogenasa Dihydrolipoamiddehydrogenasa, aktivní místo Tyr 181 120
, dihydrolipoamiddehydrogenasa. Dihydrolipoamiddehydrogenasa, aktivní místo. Tyr")
121
Lipoyllysylové raménko E2 (2x)
Raménko přenáší meziprodukty reakce mezi jednotlivými enzymy. Dihydrolipoyltransacetylasa (E2), lipoyllysylové raménko 121 121
, lipoyllysylové raménko")
122
Animace PD

123
Toxicita arsenitanu a organických sloučenin arsenu
Toxicita arsenitanu a organických sloučenin arsenu. Inhibují pyruvátdehydrogenasu a 2-oxoglutarátdehydrogenasu a tím i respiraci. Pyruvátdehydrogenasa, arsenitan Pyruvátdehydrogenasa, organické sloučeniny arsenu 123 123

124
Odbourávání aminokyselin s větveným řetězcem.
Leu je transaminován na odpovídající ketokyselinu a-ketoizokaprovou. Tato a-ketokyselina je oxidativně dekarboxylována na isovaleryl CoA enzymem: komplex dehydrogenasy větvených a-ketokyselin. Isovaleryl CoA je dehydrogenován (FAD, analogie b-oxidace)) na b-methylkrotonyl CoA. Jeho další karboxylací (ATP a CO2) se tvoří 3-hydroxy-3-methylglutaryl CoA (posléze acetyl CoA a acetoacetát) Val a Ile – transaminace, oxidativní dekarboxylace probíhá analogicky jako převod pyruvátu na acetyl CoA a a-ketoglutarátu na sukcinyl CoA. Ile poskytuje acetyl CoA a propionyl CoA, zatímco Val poskytuje CO2 a propionyl CoA.
) na b-methylkrotonyl CoA. Jeho další karboxylací (ATP a CO2) se tvoří 3-hydroxy-3-methylglutaryl CoA (posléze acetyl CoA a acetoacetát) Val a Ile – transaminace, oxidativní dekarboxylace probíhá analogicky jako převod pyruvátu na acetyl CoA a a-ketoglutarátu na sukcinyl CoA. Ile poskytuje acetyl CoA a propionyl CoA, zatímco Val poskytuje CO2 a propionyl CoA.")
125
Dehydrogenasakinasa a-ketokyselin s rozvětveným řetězcem (EC 2. 7. 2
Inaktivace enzymu fosforylací. Dalším zětnovazebným inhibitorem je a-ketoisokaproát (produkt transaminace Leu). Aktivace příslušnou fosfatasou.
. Aktivace příslušnou fosfatasou.")
130
Proteinfosfatasy obrací působení proteinkinas.

131
Fosforylace a defosforylace nejsou procesy samozřejmně reversibilní
Fosforylace a defosforylace nejsou procesy samozřejmně reversibilní. Každý je za fyziologických podmínek ireversibilní. Probíhají pouze za přítomnosti enzymů. Rychlost přechodu fosforylované a nefosforylované formy závisí na relativní aktivitě kinasy a fosfatasy. Výsledek dvou reakcí závisí na hydrolýze ATP na ADP a Pi (- 50 kJ/mol za podmínek buňky).
.")
132
Proč je fosforylace tak efektívní způsob regulace aktivity proteinů ?
1. Fosfát vnáší do molekuly proteinu dva negatívní náboje. Původní elektrostatické interakce jsou rušeny a vznikají nové. Modifikuje vazbu substrátu atp. 2. Fosfátová skupina může tvořit tři nebo více vodíkových vazeb. Tetrahedrální geometrie fosfátových skupin dává těmto vodíkovým vazbám jednoznačný směr. 3. Volná energie fosforylace je vysoká (- 50 kJ/mol). Fosforylace tak posunuje konformační rovnováhu mezi různými funkčními stavy faktorem 104.
. Fosforylace tak posunuje konformační rovnováhu mezi různými funkčními stavy faktorem 104.")
133
4. Fosforylace a defosforylace probíhají v časech kratších než sekunda, maximálně v hodinách. Reaguje se tak rychle na fyziologickou potřebu. 5. Fosforylace obvykle vede k podstatnému zvýšení rychlosti. Kinasa rychle fosforyluje velké množství proteinů. 6. ATP je energetické platidlo buňky. Použití ATP je spojeno s energetickým stavem buňky.

134
Proteinkinasy Proteinkinasy jsou kinasové enzymy modifikující druhé proteiny vstupem fosfátové skupiny – fosforylací. Výsledkem fosforylace je funkční změna cílového proteinu (substrátu) a tím současně změna enzymové aktivity, změna lokalizace proteinu v buňce nebo asociace fosforylovaného proteinu s jinými proteiny. Lidský genom obsahuje 500 proteinkinasových genů, což představuje 2% všech lidských genů. Proteinkinasy jsou obsaženy také v bakteriích a rostlinách. Zhruba 30% lidských proteinů je modifikována kinasovou aktivitou. Kinasy regulují hlavní buněčné dráhy, zvláště ty, které se uplatňují při přenosu signálů. K fosforylaci dochází na vedlejších řetězcích aminokyselin Ser, Thr a Tyr. Existují i proteinkinasy fosforylující His. Kinasy jsou aktivovány a deaktivovány fosforylací (často samotnými kinasami, tzv. cis-fosforylace nebo autofosforylace), vazbou aktivačního proteinu nebo inhibičního proteinu, malými molekulami a kontrolou jejich pozice v buňce ve vztahu k substrátům.
 a tím současně změna enzymové aktivity, změna lokalizace proteinu v buňce nebo asociace fosforylovaného proteinu s jinými proteiny. Lidský genom obsahuje 500 proteinkinasových genů, což představuje 2% všech lidských genů. Proteinkinasy jsou obsaženy také v bakteriích a rostlinách. Zhruba 30% lidských proteinů je modifikována kinasovou aktivitou. Kinasy regulují hlavní buněčné dráhy, zvláště ty, které se uplatňují při přenosu signálů. K fosforylaci dochází na vedlejších řetězcích aminokyselin Ser, Thr a Tyr. Existují i proteinkinasy fosforylující His. Kinasy jsou aktivovány a deaktivovány fosforylací (často samotnými kinasami, tzv. cis-fosforylace nebo autofosforylace), vazbou aktivačního proteinu nebo inhibičního proteinu, malými molekulami a kontrolou jejich pozice v buňce ve vztahu k substrátům.")
135
Rodina lidských proteinkinas se dělí (kinom):
CaM kinasy = obsahující Ca2+ /calmodulin dependentní proteinkinasy. CK1 obsahující kaseinkinasovou skupinu CMGC – obsahující CDK, MAPK, GSK a CLK kinasy. STE – obsahující homology kvasničných kinas Sterile 7, Sterile 11 a Sterile 20. TK – obsahující tyrosinkinasy. CDK = Cyklindependentní kinasy, regulace buněčného cyklu, GSK gĺykogensynthasakinasa; CLK = aktivují tyrosinové fosfatasy. Cyklin Lammer.

136
Buněčný cyklus M=mitosa, S=syntéza, I=interfáze, G=mezifáze (Gap phase)
")
137
MAP kinasy Mitogen-activated protein (MAP) kinases (EC ) jsou serin/threonin specifické proteinkinasy, které jsou odezvou na extracelulární stimuly (mitogeny, osmotický stres, tepelný šok a prozánětlivé cytokiny. Regulují řadu buněčných aktivit, jako jsou: genová exprese, mitosa, difernciace, proliferace a přežití buňky nebo apoptosu.
 kinases (EC ) jsou serin/threonin specifické proteinkinasy, které jsou odezvou na extracelulární stimuly (mitogeny, osmotický stres, tepelný šok a prozánětlivé cytokiny. Regulují řadu buněčných aktivit, jako jsou: genová exprese, mitosa, difernciace, proliferace a přežití buňky nebo apoptosu.")
138
Calmodulin (CaM) (zkratka pro Calcium Modulated protein).
Kyselý protein vážící Ca2+ v eukaryotních buňkách. Váže se a reguluje řadu proteinových molekul a ovlivňuje tak mnoho různých funkcí buněk. CaM ovlivňuje: zánět, metabolismus, apoptosu, kontrakci hladkého svalstva, vnitrobuněčný pohyb, krátkodobou i dlouhodobou paměť, růst nervových vláken a imunitní odpověď.

140
Kuličkový model anorganického fosfátu HPO42-
Kuličkový model anorganického fosfátu HPO42-. Barvy: P (oranžová), O (červená), H (bílá).
, O (červená), H (bílá).")
141
Funkce proteinkinas

142
Specifické Ser/Thr proteinkinasy (EC 2.7.11.1)
Fosforylují OH Ser a Thr zbytků. Aktivita je regulována poškozením DNA, chemickými signály jako jsou cAMP/cGMP, diacylglycerol a Ca2+/calmodulin. Velmi důležitou skupinou proteinkinas jsou MAP kinasy (akronym: „mitogen/microtubule-activated protein kinases). Dva hlavní faktory vlivu aktivity MAP kinas: a) Signály aktivují transmembránové receptory (mohou to být přirozené ligandy nebo zesíťovací látky a proteiny spojené s nimi (mutace simulující aktivní stav). b) Signály aktivující fosfatasy, které ruší MAP. Takové signály zahrnují např. oxidační stres.
. Dva hlavní faktory vlivu aktivity MAP kinas: a) Signály aktivují transmembránové receptory (mohou to být přirozené ligandy nebo zesíťovací látky a proteiny spojené s nimi (mutace simulující aktivní stav). b) Signály aktivující fosfatasy, které ruší MAP. Takové signály zahrnují např. oxidační stres.")
143
Specifické tyrosinové kinasy (EC 2.7.10.1 a EC 2.7.10.2)
Fosforylují vedlejší řetězec Tyr a jsou stejně jako Ser/Thr kinasy součástí přenosu signálu. Působí především jako receptory růstového faktoru. Některé příklady: Epidermální růstový faktor (EGFR) Insulinový receptor
 Insulinový receptor.")
144
Inhibitory proteinkinas.
Deregulace kinasové aktivity je často spojována s onemocněními, zvláště pak s rakovinou. Kinasy regulují mnohé aspekty kontroly růstu buněk, jejich pohybu a smrti. Byla vyvinuta řada farmak k léčení některých onemocnění. Řada se klinicky využívá. Je to např. Gleevec (imatinib) a Iressa (gefitinib), Anthra(1,9-cd)pyrazol-6(2H)-on a Staurosporin.
 a Iressa (gefitinib), Anthra(1,9-cd)pyrazol-6(2H)-on a Staurosporin.")
145
Imatinib (Gleevec) je používán při léčbě chronické myelogenní leukemie (CML), tumorů zažívacího traktu. Inhibuje tyrosinkinasy. Chronická myeloproliferativní onemocnění jsou chronicky (=pomalu a plíživě) probíhající nádorová onemocnění, která vycházejí z buněk kostní dřeně dávajících vzniknout všem krevním buňkám s výjimkou lymfocytů.
 probíhající nádorová onemocnění, která vycházejí z buněk kostní dřeně dávajících vzniknout všem krevním buňkám s výjimkou lymfocytů.")
146
Antibiotikum Staurosporin z bakterie Streptomyces staurosporeus
Antibiotikum Staurosporin z bakterie Streptomyces staurosporeus. Biologická aktivita spočívá v inhibici proteinkinas. Váže se do jejich aktivního místa a zabraňuje tak vazbě ATP. Indukuje apoptosu.

147
Přenos signálu, G-protein(trimer a, b a g), druhý posel, proteinkinasa A.
, druhý posel, proteinkinasa A.")
148
První fáze aktivace proteinkinasy A.

149
Druhá fáze aktivace proteinkinasy A
Druhá fáze aktivace proteinkinasy A. Vytvořený cAMP se váže na regulační podjednotky PKA a uvolňuje aktivní katalytické podjednotky.

150
Regulace proteinkinasy A. (EC 2. 7. 11
Regulace proteinkinasy A. (EC ) Vazbou čtyř molekul cAMP je proteinkinasa A aktivována. Inhibovaný holoenzym (R2C2) disociuje po vazbě cAMP na regulační podjednotku (R2) a katalyticky aktivní podjednotku (C. 150
 Vazbou čtyř molekul cAMP je proteinkinasa A aktivována. Inhibovaný holoenzym (R2C2) disociuje po vazbě cAMP na regulační podjednotku (R2) a katalyticky aktivní podjednotku (C")
151
Cyklický AMP (cAMP) aktivuje proteinkinasu A změnou její struktury.
 aktivuje proteinkinasu A změnou její struktury.")
152
Aktivační a deaktivační mechanismus proteinkinasy A
Aktivační a deaktivační mechanismus proteinkinasy A. PDE = fosfodiesterasa

153
Rovnováha mezi glykolýzou a glukoneogenezí v játrech – vliv hladiny glukosy v krvi.
Hlavním regulátorem glykolýzy a glukoneogeneze v játrech je fruktosa-2,6-bisfosfát. Fruktosa-2,6-bisfosfát aktivuje fosfofruktokinasu a inhibuje fruktosa 2,6-bisfosfatasu !!! Proč ?? Při nízké hlasině glukosy v krvi se z fruktosa-2,6-bisfosfátu uvolňuje fosfát za tvorby fruktosa-6-fosfátu. Fruktosa-6-fosfát je substrátem fosfofruktokinasy. Jakým způsobem je kontrolována koncentrace fruktosa-2,6-bisfosfátu ??? Na kontrole hladiny fruktosa-2,6-bisfosfátu se podílejí dva enzymy.

154
Rovnováha mezi glykolýzou a glukoneogenezí v játrech – vliv hladiny glukosy v krvi.
Fruktosa-2,6-bisfosfát je produktem reakce katalyzované fosfofruktokinasou 2 (PFK2). Odštěpení fosfátu je katalyzované enzymem fruktosabisfosfatasa2 (FBPasa2) Oba enzymy jsou součástí jednoho proteinového řetězce o délce 55 kD – bifunkční enzym !!! Co kontroluje zda bude aktivní PFK2 nebo FBPasa2 ? Za situace hladovění: Při nízké hladině glukosy je vylučován slinivkou hormon glukagon, který spouští uvnitř buněk cAMP kaskadu vedoucí k fosforylaci bifunkčního enzymu proteinkinasou A. Tato kovalentní modifikace aktivuje FBPasu2 a inhibuje PFK2. Převládá glukoneogeneze. V opačném případě, při dostatku glukosy, je fosfát odštěpen, aktivuje se PFK2 a inhibuje FBAsa2. Urychluje se glykolýza!!! Glukagon také aktivuje pyruvátkinasu v játrech.
. Odštěpení fosfátu je katalyzované enzymem fruktosabisfosfatasa2 (FBPasa2) Oba enzymy jsou součástí jednoho proteinového řetězce o délce. 55 kD – bifunkční enzym !!! Co kontroluje zda bude aktivní PFK2 nebo FBPasa2 Za situace hladovění: Při nízké hladině glukosy je vylučován slinivkou hormon glukagon, který spouští uvnitř buněk cAMP kaskadu vedoucí k fosforylaci bifunkčního enzymu proteinkinasou A. Tato kovalentní modifikace aktivuje FBPasu2 a inhibuje PFK2. Převládá glukoneogeneze. V opačném případě, při dostatku glukosy, je fosfát odštěpen, aktivuje se PFK2 a inhibuje FBAsa2. Urychluje se glykolýza!!! Glukagon také aktivuje pyruvátkinasu v játrech.")
155
Fruktosa-2,6-bisfosfát.

156
Bifunkční enzym fosfofruktokinasa2-fosfofruktobisfosfatasa2.

157
Kontrolní mechanismus syntézy a odbourání fruktosa-2,6-bisfosfátu.

158
Pyruvátkinasa (EC 2.7.1.40). Fosfoenolpyruvát + ADP + Pi pyruvát + ATP
Pyruvátkinasa je tetramer (57 kD podjednotka). Existuje řada izoenzymových forem kódovaných různými geny. L typ převažuje v játrech a M ve svalech a mozku. Jaterní enzym podléhá na rozdíl od svalového allosterické regulaci. Katalytická aktivita L formy je kontrolována reversibilní fosforylací. Aktivita M formy ne. Při nízké hladině glukosy funguje glukagon. Spouští se cAMP kaskáda – dochází k fosforylaci pyruvátkinasy a tím ke snížení její aktivity. Tato hormonální regulace zabraňuje spotřebě glukosy játry !!! Glukosa je nutně potřebná k činnosti mozku a svalstva. Při vysoké hladině glukosy se uvolňuje insulin, který aktivuje fosfoproteinfosfatasu, což vede k defosforylce pyruvátkinasy a tím k její aktivaci!!
. Existuje řada izoenzymových forem kódovaných různými geny. L typ převažuje v játrech a M ve svalech a mozku. Jaterní enzym podléhá na rozdíl od svalového allosterické regulaci. Katalytická aktivita L formy je kontrolována reversibilní fosforylací. Aktivita M formy ne. Při nízké hladině glukosy funguje glukagon. Spouští se cAMP kaskáda – dochází k fosforylaci pyruvátkinasy a tím ke snížení její aktivity. Tato hormonální regulace zabraňuje spotřebě glukosy játry !!! Glukosa je nutně potřebná k činnosti mozku a svalstva. Při vysoké hladině glukosy se uvolňuje insulin, který aktivuje fosfoproteinfosfatasu, což vede k defosforylce pyruvátkinasy a tím k její aktivaci!!")
159
Kontrolní systém katalytické aktivity pyruvátkinasy
Kontrolní systém katalytické aktivity pyruvátkinasy. Regulace allosterickou kontrolou aktivátory, inhibitory a kovalentní modifikací.

160
Otázky Uvažujte co se stane, když bude paruvátkinasa v našem organismu deficitní (uvažujte různé typy buněk). Může být pyruvátkinasa současně aktivní s pyruvátkarboxylasou a fosfoenolpyruvátkarboxykinasou ?

161
4. Mnoho enzymů je aktivováno specifickým proteolytickým štěpením.

162
Specifická proteolýza:
1. Trávící enzymy jsou syntetizovány jako zymogeny v žaludku a pankreatu. 2. Srážení krve je modifikováno kaskádou proteolytických aktivací, které jsou rychlou odezvou na poranění. 3. Některé proteinové hormony jsou syntetizovány jako inaktivní prekurzory. Např. insulin je syntetizován jako preproinsulin, proteolyticky se oddělují dva peptidy. 4. Fibrilární protein kolagen, hlavní součást kůže a kloubů, je tvořen z prokolagenu, rozpustného prekurzoru. 5. Mnohé vývojové procesy jako např. metamorfóza pulce na žábu. Velké množství kolagenu je resorbováno z bičíku v několika dnech. 6. Programová smrt buňky, apoptóza, je zprostředkována proteasami jako je kaspasa, syntetizována z prekurzoru prokaspasy.

163
Sekrece zymogenů acinárními (hroznovitými) buňkami pankreatu.
 buňkami pankreatu.")
164
Proteolytická aktivace chymotrypsinogenu
Proteolytická aktivace chymotrypsinogenu. Tři řetězce a-chymotrypsinu jsou vázány dvěma disulfidovými vazbami mezi řetězci (A k B a B k C).
.")
165
Konformace chymotrypsinogenu (červeně) a chymotrypsinu (modře).
Elektrostatická interakce mezi karboxylem Asp 194 a a-aminoskupinou Ile 16 je nutná pro strukturu aktivního chymotrypsinu.

166
Aktivace zymogenů proteolytickým štěpením
Aktivace zymogenů proteolytickým štěpením. Aktivní enzymy jsou žlutě, zymogeny oranžově. Enteropeptidasa iniciuje aktivaci. Trypsin je společný aktivátor všech pankreatických zymogenů. Buňky dvanácterníku vylučují enteropeptidasu, která hydrolyzuje Lys-Ile vazbu trypsinogenu.

167
Inhibitory proteolytických enzymů.
Aktivace zymogenů je ireversibilní. K omezení nebo inaktivaci aktivních enzymů se využívají specifické inhibitory proteas. Např. pankreatický trypsinový inhibitor (antitrypsin) je 6 kD protein inhibující trypsin tak, že se váže velmi pevně do jeho aktivního místa. Inhibiční konstanta je 1 pM !! Komplex nelze oddělit ani denaturací 8 M močovinou nebo 6 M guanidinHCl. Nejznámějším trypsinovým inhibitorem je tzv. Kunitzův inhibitor z hovězího pankreatu (6,5 kD). Inhibuje i chymotrypsin, ale slaběji. Další: Fazol obsahuje šest různých inhibitorů (6-10 kD) Soja obsahuje několik inhibitorů v rozmezí 20,7 až 22,3 kD.
 je 6 kD protein inhibující trypsin tak, že se váže velmi pevně do jeho aktivního místa. Inhibiční konstanta je 1 pM !! Komplex nelze oddělit ani denaturací 8 M močovinou nebo. 6 M guanidinHCl. Nejznámějším trypsinovým inhibitorem je tzv. Kunitzův inhibitor z hovězího pankreatu (6,5 kD). Inhibuje. i chymotrypsin, ale slaběji. Další: Fazol obsahuje šest různých inhibitorů (6-10 kD) Soja obsahuje několik inhibitorů v rozmezí 20,7 až 22,3 kD.")
168
Interakce trypsinu se svým inhibitorem
Interakce trypsinu se svým inhibitorem. Trypsin (žlutý), inhibitor (červený). Lys 15 inhibitoru vstupuje do aktivního místa. Vytváří iontovou vazbu s Asp 189.
, inhibitor (červený). Lys 15 inhibitoru vstupuje do aktivního místa. Vytváří iontovou vazbu s Asp 189.")
169
Proč existuje trypsinový inhibitor ??
Trypsin aktivuje celou řadu zymogenů. Inhibitorem je inaktivována jen velmi malá část trypsinové aktivity. Molekuly trypsinu aktivované ve žlučníku nebo v pankreatu by způsobily poškození tkáně – akutní pankreatitida.

170
Peptidasy, proteinasy (proteasy)…
Peptidasy jsou enzymy ze třídy hydrolas, které štěpí peptidovou vazbu za účasti vody. Do této rozsáhlé skupiny řadíme zejména aminopeptidasy, karboxypeptidasy, dipeptidasy a proteinasy. Neexistuje jasné rozhraní mezi proteinasami a peptidasami. Proteinasy dělíme podle katalytických skupin v aktivním místě: A) Serinové (rozhodující je -OH skupina serinu, např. trypsin, thrombin), B) Aspartátové (skupina -COO-, např. pepsin), C) Cysteinové (-SH, papain), D) Metalloproteasy (často Zn2+, karboxypeptidasa).
 Serinové (rozhodující je -OH skupina serinu, např. trypsin, thrombin), B) Aspartátové (skupina -COO-, např. pepsin), C) Cysteinové (-SH, papain), D) Metalloproteasy (často Zn2+, karboxypeptidasa).")
171
Substrátová specifita proteinas
Trypsin – štěpí peptidovou vazbu na místě karboxylu bázických aminokyselin, zejména Arg a Lys. Chymotrypsin – štěpí peptidy na straně karboxylu Tyr, Trp a Phe, protože tyto aminokyseliny mají aromatický kruh, který vstupuje do hydrofobní kapsy enzymu. Také štěpí další peptidové vazby, hlavně na karboxylové straně Leu. Papain - nejlépe štěpí druhou peptidovou vazbu následující směrem k C konci za zbytkem Phe. Pepsin - nemá velkou specifitu, nejlépe štěpí peptidové vazby na karboxylové straně aromatických aminokyselin (Tyr, Phe). Produktem působení pepsinu na bílkoviny je tzv. pepton, směs peptidů obsahujících obvykle aminokyselinových zbytků. Elastasa (pankreatická) – štěpí elastin, elastinová vlákna, která spolu s kolagenem tvoří mechanické vlastnosti spojovacích tkání. Štěpí peptidové vazby na karboxalové straně Gly, Val a Ala.
. Produktem působení pepsinu na bílkoviny je tzv. pepton, směs peptidů obsahujících obvykle aminokyselinových zbytků. Elastasa (pankreatická) – štěpí elastin, elastinová vlákna, která spolu s kolagenem tvoří mechanické vlastnosti spojovacích tkání. Štěpí peptidové vazby na karboxalové straně Gly, Val a Ala.")
172
Karboxypeptidasa A preferuje štěpení na karboxylovém konci aromatických a větvených aminokyselin.
Karboxypeptidasa B štěpí na karboxylovém konci aminokyseln Arg a Lys. Serinová karboxypeptidasa štěpící C – koncovou aminokyselinu z peptidu se sekvencí –Pro-Xaa (Xaa je libovolná aminokyselina na C konci peptidu) se nazývá „prolylkarboxypeptidasa“. Thrombin – selektivně štěpí peptidovou vazbu Arg-|-Gly fibrinogenu za tvorby fibrinu a a dvou fibrinopeptidů A a B.
 se nazývá „prolylkarboxypeptidasa . Thrombin – selektivně štěpí peptidovou vazbu Arg-|-Gly fibrinogenu za tvorby fibrinu a a dvou fibrinopeptidů A a B.")
173
Hemokoagulace, srážení krve
Principem srážení krve je tvorba nerozpustné sítě tvořené fibrinem, do které se zachytí červené krvinky a krevní destičky. Vzniklý červený trombus, tedy „krevní sraženina“ ucpe defekt ve stěně cévy a zamezí tak krvácení. Většina koagulačních faktorů má charakter serinových proteinas. V plazmě, ale kolují v neaktivní formě, a jejich aktivace spočívá v jejich štěpení enzymem, který byl aktivován v předchozí reakci. Aktivace koagulační kaskády může probíhat dvěma způsoby: A) V případě, že je spouštěcím impulzem poškození nebo adheze na cévní stěnu za vzniku abnormální cévní stěny nebo omezení toku krve, uplatňuje se tzv. vnitřní systém. B) Je-li to odpověď na poškození tkání, vstupuje do hry tzv. vnější systém koagulace. Oba dva systémy nicméně splývají ve společnou dráhu, která vede k samotné tvorbě fibrinové sraženiny.
 V případě, že je spouštěcím impulzem poškození nebo adheze na cévní stěnu za vzniku abnormální cévní stěny nebo omezení toku krve, uplatňuje se tzv. vnitřní systém. B) Je-li to odpověď na poškození tkání, vstupuje do hry tzv. vnější systém koagulace. Oba dva systémy nicméně splývají ve společnou dráhu, která vede k samotné tvorbě fibrinové sraženiny.")
174
Srážení krve se uskutečňuje kaskádou aktivace zymogenů.
Sraženina fibrinu se tvoří souhrou vnitřních a vnějších faktorů a v konečné fázi společných drah. Vnitřní faktor začíná aktivací faktoru XII kontaktem s neobvyklým povrchem způsobeným zraněním. Vnitřním faktorem je trauma aktivující faktor VII a uvolňující lipoprotein z krevních destiček. Inaktivní srážecí faktor je červeně, aktivovaný žlutý. Stimulační proteiny jsou modře. Principem je, že aktivovaný faktor aktivuje faktor následující.

175
Kaskáda srážení krve – kaskáda aktivací zymogenů
Kaskáda srážení krve – kaskáda aktivací zymogenů. Fibrin se tvoří souhrou vnitřních, vnějších a v konečné fázi společných drah. XII- Hagemanův faktor, „tissue factor“ = lipoprotein z krevních destiček

176
Kininogen, kallikrein. Systém kinin-kallikrein, zjednodušeně kininový systém, je systém krevních proteinů, které se uplatňují při zánětech, tlaku krve, kontrole koagulace a bolesti. Kallikreiny (tkáňový a plasmový kallikrein) jsou serinové proteasy uvolňující kininy (bradykinin, kallidin) z kininogenů. Bradykinin (BK) – je nonapeptid s aminokyselinovou sekvencí Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg. BK rozšiřuje (dilatuje) krevní kapiláry a snižuje tak tlak krve (TK). Třída látek zvaná ACE inhibitory, využívaná terapeuticky ke snižování TK, zvyšuje hladinu BK a tím snižují TK. Působení bradykininu spočívá v uvolňování prostacyklinu a oxidu dusnatého.
 jsou serinové proteasy uvolňující kininy (bradykinin, kallidin) z kininogenů. Bradykinin (BK) – je nonapeptid s aminokyselinovou sekvencí Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg. BK rozšiřuje (dilatuje) krevní kapiláry a snižuje tak tlak krve (TK). Třída látek zvaná ACE inhibitory, využívaná terapeuticky ke snižování TK, zvyšuje hladinu BK a tím snižují TK. Působení bradykininu spočívá v uvolňování prostacyklinu a oxidu dusnatého.")
177
ACE inhibitory nebo také inhibitory Angiotensin-Converting-Enzyme,
je skupina farmak používaných prvotně ke snižování TK. Na vedlejším obrázku je Captopryl (Cynt). Angiotensin converting enzyme (ACE, EC ) je exopeptidasa. Katalyzuje štěpení angiotensinu I na angiotensin II, což je „smršťovač cév“ (vasoconstrictor) inaktivující funkci bradykininu (vasodilatátor).
. Angiotensin converting enzyme (ACE, EC ) je exopeptidasa. Katalyzuje štěpení angiotensinu I na angiotensin II, což je „smršťovač cév (vasoconstrictor) inaktivující funkci bradykininu (vasodilatátor).")
178
Dva způsoby aktivace koagulační kaskády
Zevní systém: Při poškození stěny cévy se krevní plazma dostane do styku s tkáňovým faktorem, tromboplastinem (faktor III), který se vyskytuje na povrchu mnoha extravaskulárních buněk. Tromboplastin je kofaktorem faktoru VII v reakci, při níž je aktivován faktor X. Vnitřní systém: Vnitřní dráha začíná stykem s negativně nabitým "aktivačním povrchem“. In vivo jím bývá kolagen, in vitro například sklo. Při styku dojde k aktivaci faktoru XII proteolytickým působením kalikkreinu. Faktor XIIa dále aktivuje faktor IX. Kromě toho štěpí prekalikrein na další kallikrein (pozitivní zpětná vazba) a z vysokomolekulárního kininogenu uvolňuje bradykinin, látku s výraznými vazodilatačními účinky. Faktor IXa, v reakci, ke které jsou nutné ionty vápníku a faktor VIIIa jako kofaktor, štěpí faktor X na Xa.
, který se vyskytuje na povrchu mnoha extravaskulárních buněk. Tromboplastin je kofaktorem faktoru VII v reakci, při níž je aktivován faktor X. Vnitřní systém: Vnitřní dráha začíná stykem s negativně nabitým aktivačním povrchem . In vivo jím bývá kolagen, in vitro například sklo. Při styku dojde k aktivaci faktoru XII proteolytickým působením kalikkreinu. Faktor XIIa dále aktivuje faktor IX. Kromě toho štěpí prekalikrein na další kallikrein (pozitivní zpětná vazba) a z vysokomolekulárního kininogenu uvolňuje bradykinin, látku s výraznými vazodilatačními účinky. Faktor IXa, v reakci, ke které jsou nutné ionty vápníku a faktor VIIIa jako kofaktor, štěpí faktor X na Xa.")
179
Společná dráha Aktivovaný faktor X je serinová proteáza, která štěpí prothrombin (faktor II) na aktivní thrombin. Aktivace se odehrává na povrchu aktivovaných destiček a vyžaduje vznik protrombinasového komplexu aktivátoru protrombinu, skládající se z destičkových aniontových fosfolipidů, Ca2+ faktoru Va a Xa a prothrombinu. Thrombin štěpí fibrinopeptidy fibrinogenu a uvolňuje tak fibrin-monomery, které spontánně polymerují. Kromě toho aktivují faktor VIII, který vzniklou sraženinu stabilizuje vznikem kovalentních vazeb mezi sousedními molekulami fibrin-polymeru. Výsledkem je nerozpustná fibrinová síť.
 na aktivní thrombin. Aktivace se odehrává na povrchu aktivovaných destiček a vyžaduje vznik protrombinasového komplexu aktivátoru protrombinu, skládající se z destičkových aniontových fosfolipidů, Ca2+ faktoru Va a Xa a prothrombinu. Thrombin štěpí fibrinopeptidy fibrinogenu a uvolňuje tak fibrin-monomery, které spontánně polymerují. Kromě toho aktivují faktor VIII, který vzniklou sraženinu stabilizuje vznikem kovalentních vazeb mezi sousedními molekulami fibrin-polymeru. Výsledkem je nerozpustná fibrinová síť.")
181
Tvorba fibrinové sraženiny
Tvorba fibrinové sraženiny. Thrombin (proteolytický enzym) štěpí fibrinopeptidy z centrální globule fibrinogenu. Vytvořené peptidy polymerují na způsob sítě.
 štěpí fibrinopeptidy z centrální globule fibrinogenu. Vytvořené peptidy polymerují na způsob sítě.")
182
Nově vytvořená sraženina se stabilizuje zesíťováním za účasti transglutaminasy (faktor XIIIa).
.")
183
Thrombin je syntetizován jako zymogen z prothrombinu.
Prothrombin obsahuje čtyři domény. Aktivace se uskutečňuje proteolytickým štěpením. Vitamin K je nutný k syntéze prothrombinu. Antagonisté vitaminu K jsou dikumarol a warfarin. Dikumarol se používá v lékařství jako antikoagulant. Warfarin a další antagonisté vit. K slouží také jako jed na hlodavce. Za přítomnosti antagonistů vitaminu K je syntetizován abnormální prothrombin, který neváže Ca2+.

184
Normální prothrombin váže Ca2+ modifikovanou aminokyselinou g-karboxyglutamátem (silná chelatace). Působením antagonistů vit. K dochází k syntéze prothrombinu bez vázaného Ca2+ Vazba Ca2+ umožňuje prothrombinu (zymogenu) vstoupit do fosfolipidové membrány vzniklé z krevních destiček po poranění. Tím se dostává do blízkosti proteinů, které ho aktivují na thrombin.
 vstoupit do fosfolipidové membrány vzniklé z krevních destiček po poranění. Tím se dostává do blízkosti proteinů, které ho aktivují na thrombin.")
185
Úloha vitaminu K a jeho antagonistů při srážení krve.
Vitamin K je nutný k syntéze prothrombinu a dalších faktorů srážení krve. Antagonisty vitaminu K jsou dikumarol a warfarin. Oba antagonisté se používají jako antikoagulancia.

186
Regulace srážení krve Proces srážení krve musí být citlivě regulovaný, aby nedošlo ke vzniku nežádoucích krevních sraženin. Thrombin sám se váže na thrombomodulin na povrchu endotelových buněk, což vede k aktivaci proteinu Ca. Protein Ca společně s proteinem S degradují faktory Va a VIIIa (negativní zpětná vazba). Kromě toho se v plazmě vyskytují další látky, působící proti koagulaci: Antithrombin III: je nejvýznamnějším inhibitorem aktivity trombinu, kromě toho inaktivuje také faktory IXa, Xa, XIa a XIIa. Aktivita antithrombinu III je zvyšována přítomností heparinu, což je principem jeho působení v působení proti vzniku krevních sraženin. Plazmin je serinová proteinasa, která degraduje fibrinogen a fibrin. Koluje v krvi jako neaktivní plazminogen, který je aktivován tkáňovým aktivátorem plazminogenu nebo urokinasou a naopak je rychle degradován α2antiplazminem, takže celý systém je v dynamické rovnováze a nedochází ke krvácení nebo ke vzniku trombózy.
. Kromě toho se v plazmě vyskytují další látky, působící proti koagulaci: Antithrombin III: je nejvýznamnějším inhibitorem aktivity trombinu, kromě toho inaktivuje také faktory IXa, Xa, XIa a XIIa. Aktivita antithrombinu III je zvyšována přítomností heparinu, což je principem jeho působení v působení proti vzniku krevních sraženin. Plazmin je serinová proteinasa, která degraduje fibrinogen a fibrin. Koluje v krvi jako neaktivní plazminogen, který je aktivován tkáňovým aktivátorem plazminogenu nebo urokinasou a naopak je rychle degradován α2antiplazminem, takže celý systém je v dynamické rovnováze a nedochází ke krvácení nebo ke vzniku trombózy.")
187
Hemofilie Hemofilie je geneticky podmíněné onemocnění projevující se poruchou srážlivosti krve, což se navenek projevuje chorobnou krvácivostí – krevními výrony do svalů či kloubů a omezenou schopností organismu zastavit krvácení. Chorobou se zabývá hematologie. Hemofilie A = absence nebo nedostatek Faktoru srážlivosti VIII. Hemofilie B = absence nebo nedostatek Faktoru srážlivosti IX. Přenáší se ve formě vadné alely na chromozomu X, neboli jde o onemocnění gonosomálně recesivní. Obvykle postihuje pouze muže (není příliš pravděpodobné, že by se sešly dvě vadné alely, žena s jednou vadnou alelou je pouze skrytá přenašečka). Muži nemohou být skrytými přenašeči, jsou buďto nemocní (v tom případě jsou všechny jejich dcery přenašečky a všichni synové zdraví – za předpokladu, že matka nebyla přenašečkou), nebo zdraví. Chybný gen se váže na chromozom X, který s sebou nese osudnou informaci.
. Muži nemohou být skrytými přenašeči, jsou buďto nemocní (v tom případě jsou všechny jejich dcery přenašečky a všichni synové zdraví – za předpokladu, že matka nebyla přenašečkou), nebo zdraví. Chybný gen se váže na chromozom X, který s sebou nese osudnou informaci.")
188
Dědičná hemofilie Nejznámější dědičná linie vznikla díky britské královně Viktorii. Než v roce 1901 zemřela, stačila osudové geny předat svému osmému potomku, synu Leopoldovi. Ten trpěl častými krváceními a psalo se o nich i v britských odborných časopisech. Leopold zemřel na krvácení do mozku v jednatřiceti letech. Ještě osudovější bylo dědictví jejích dcer Alice a Beatrice, které rozšířily onemocnění do královských rodin Německa, Španělska a Ruska. Nejproslulejším se stal příběh ruské carské rodiny, kam zanesla poškozený gen německá princezna a vnučka královny Viktorie, Alexandra Fjodorovna. S carem Mikulášem II. měli syna Alexeje, velmi těžkého hemofilika. Časté krvácivé projevy mu prakticky neumožňovaly normální život, byl neustále obklopen bodyguardy a ošetřovateli, žil ve skleníkovém prostředí. I to byl jeden z důvodů obrovského vlivu podvodného mnicha Grigorije Novika, zvaného Rasputin, na ruský dvůr. Byl totiž schopný hypnózou mladému carevičovi ulevit od bolesti, dokonce v některých případech i zastavit krvácení. Alexej se ruského trůnu nedočkal, s celou rodinou ho bolševici zastřelili v červnu 1918. V anglickém exilu zemřeli před druhou světovou válkou také potomci královny Viktorie Evženie, synové vnučky královny Viktorie a Alfonse VIII - dva španělští princové. Oba po banálních zraněních - jeden lehce havaroval autem do telefonní budky, druhý si přivodil, také v autě lehkým nárazem na volant, krvácení do hrudní dutiny.

189
Hemofilie. Při klasické hemofilii (dědičná, A) není v krvi přítomen antihemofilní faktor VIII nebo je jeho aktivita nízká. Antihemofilní faktor (VIII) – stimuluje aktivaci faktoru X faktorem IXa. Léčí se podáním lyofilizovaných koncentrátů plazmy s faktorem VIII.
 není v krvi přítomen antihemofilní faktor VIII nebo je jeho aktivita nízká. Antihemofilní faktor (VIII) – stimuluje aktivaci faktoru X faktorem IXa. Léčí se podáním lyofilizovaných koncentrátů plazmy s faktorem VIII..")
190
K zamyšlení. 1. His v aktivním místě aspartáttranskarbamoylasy je nutný ke stabilizaci přechodového stavu vázaných substrátů. Jaká bude závislost katalytické rychlosti na pH ? Uvažujte, že tato reakce je dominantní pro pH profil enzymu. 2. Představte si, že jste vyizolovali dimerní enzym obsahující dvě stejná aktivní místa. Vazba substrátu do jednoho aktivního místa snižuje afinitu k substrátu druhého aktivního místa. Který allosterický model nejlépe vyjadřuje tuto negativní kooperativitu ? 3. Erythrocyty ptáků a želv obashují regulační molekuly odlišné od 2,3-BPG. Tyto látky jsou také účinné při snižování afinity hemoglobinu ke kyslíku, dokonce i lidského hemoglobinu zbaveného 2,3-BPG. Která z následujících látek by mohl být v tomto smyslu nejúčinnější ? a) Glukosa-6-fosfát; b) inositolhexafosfát; c) HPO42- ; d) Malonát; e) Arginin; f) Laktát.
 Glukosa-6-fosfát; b) inositolhexafosfát; c) HPO42- ; d) Malonát; e) Arginin; f) Laktát.")
191
4. Prostředí ovlivňuje částečně pK kyseliny
4. Prostředí ovlivňuje částečně pK kyseliny. Jaký vliv budou mít následující změny prostředí na pK vedlejší řetězce Glu ? a) Do blízkosti se dostane vedlejší řetězec Lys. b) Do blízkosti se dostane terminální karboxyl proteinu. c) Vedlejší řetězec Glu je posunut z vnějšku proteinu do hydrofobní kapsy uvnitř. 5. Jaký vliv má každá následující změna na afinitu hemoglobinu ke kyslíku in vitro ? a) Zvýšení pH ze 7, 2 na 7, 4; b) Zvýšení pCO2 z 10 na 40 torrů; c) Zvýšení [2,3-BPG] z 0, 2 na 0, 8 mM; d) Disociace a2b2 na monomerní podjednotky. 6. Substrát se váže 100x pevněji na R stav než na T stav allosterického enzymu. Aplikuj na enzym model MWC. a) Jakým faktorem posune vazba jedné molekuly substrátu na enzym poměr koncentrací enzymových molekul v R a T stavech ? b) Uvažujte že, poměr [T] /[R] za nepřítomnosti substrátu je 107 a, že enzym obsahuje čtyři vazebná místa pro substrát. Jaký je poměr enzymových molekul ve stavu R ku stavu T za přítomnosti saturačního množství substrátu ? 7. Allosterický enzym vyhovující modelu WMC má poměr T/R = 300 bez přítomnosti substrátu. Uvažujte mutaci, která poměr obrátí. Jak může taková mutace ovlivnit vztah mezi rychlostí reakce a koncentrací substrátu?
 Do blízkosti se dostane vedlejší řetězec Lys. b) Do blízkosti se dostane terminální karboxyl proteinu. c) Vedlejší řetězec Glu je posunut z vnějšku proteinu do hydrofobní kapsy uvnitř. 5. Jaký vliv má každá následující změna na afinitu hemoglobinu ke kyslíku in vitro a) Zvýšení pH ze 7, 2 na 7, 4; b) Zvýšení pCO2 z 10 na 40 torrů; c) Zvýšení [2,3-BPG] z 0, 2 na 0, 8 mM; d) Disociace a2b2 na monomerní podjednotky. 6. Substrát se váže 100x pevněji na R stav než na T stav allosterického enzymu. Aplikuj na enzym model MWC. a) Jakým faktorem posune vazba jedné molekuly substrátu na enzym poměr koncentrací enzymových molekul v R a T stavech b) Uvažujte že, poměr [T] /[R] za nepřítomnosti substrátu je 107 a, že enzym obsahuje čtyři vazebná místa pro substrát. Jaký je poměr enzymových molekul ve stavu R ku stavu T za přítomnosti saturačního množství substrátu 7. Allosterický enzym vyhovující modelu WMC má poměr T/R = 300 bez přítomnosti substrátu. Uvažujte mutaci, která poměr obrátí. Jak může taková mutace ovlivnit vztah mezi rychlostí reakce a koncentrací substrátu")
192
Odpovědi: 1. Protonizovaná forma His pravděpodobně stabilizuje negativní náboj kyslíkového atomu štěpitelné vazby v přechodovém stavu. Deprotonizace by mohla vést ke snížení aktivity. Lze uvažovat, že rychlost bude polovina maximální při pH kolem 6, 5 (pK His vedlejšího řetězce v proteinu) a poklesne při zvýšení pH. 2. Sekvenční model, model MWC ne. 3. b) Inositolhexafosfát. 4. Hodnota pK se a) snižuje, b) zvyšuje a c) zvyšuje. 5. a) zvyšuje se, b) snižuje se, c) snižuje se a d) zvyšuje se afinita ke kyslíku. 6. a) 100, změna poměru [R]/[T] po vazbě jednoho substrátu musí být shodná s poměrem substrátových afinit obou forem. b) 10, vazba čtyř molekul substrátu mění poměr [R]/[T] faktorem 1004 = 108. Poměr za nepřítomnosti substrátu je Z toho plyne, že poměr v plně nasycené molekule je 108 x 10-7 = 10. 7. Allosterie přejde na jednoduchou kinetiku Michaelise a Mentenové, protože se bude jednat pouze o R stav.
 a poklesne při zvýšení pH. 2. Sekvenční model, model MWC ne. 3. b) Inositolhexafosfát. 4. Hodnota pK se a) snižuje, b) zvyšuje a c) zvyšuje. 5. a) zvyšuje se, b) snižuje se, c) snižuje se a d) zvyšuje se afinita ke kyslíku. 6. a) 100, změna poměru [R]/[T] po vazbě jednoho substrátu musí být shodná s poměrem substrátových afinit obou forem. b) 10, vazba čtyř molekul substrátu mění poměr [R]/[T] faktorem 1004 = 108. Poměr za nepřítomnosti substrátu je Z toho plyne, že poměr v plně nasycené molekule je 108 x 10-7 = Allosterie přejde na jednoduchou kinetiku Michaelise a Mentenové, protože se bude jednat pouze o R stav.")
Podobné prezentace

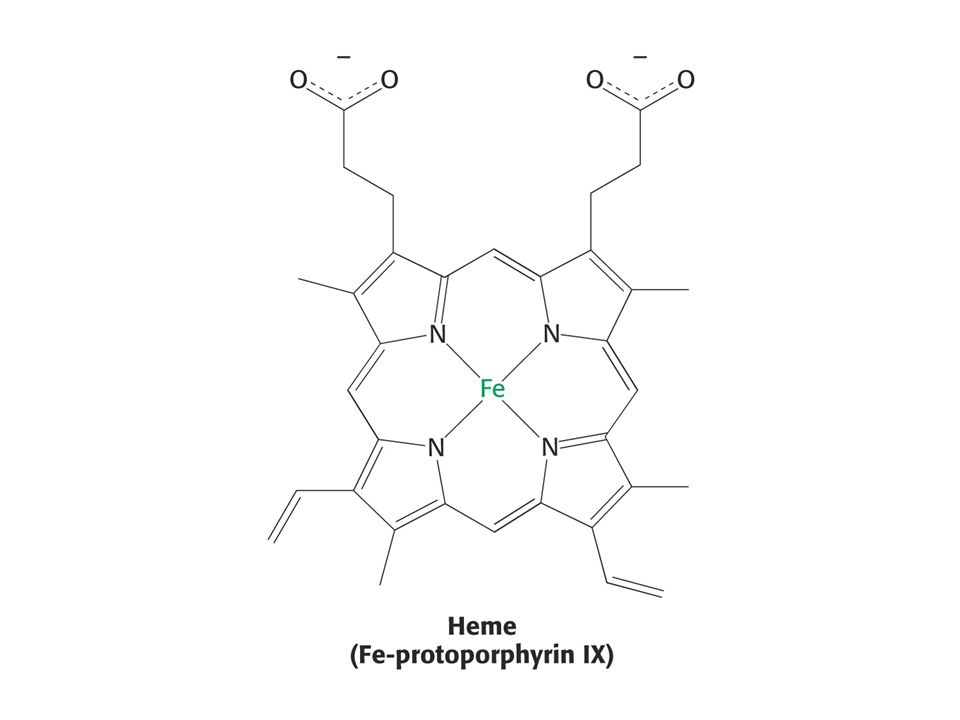
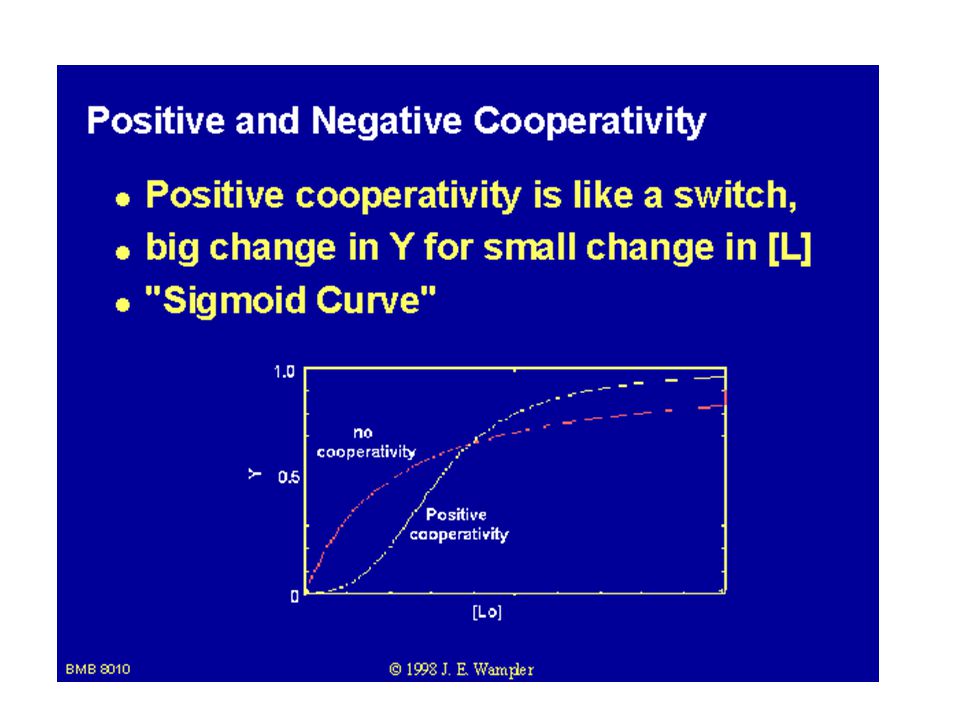
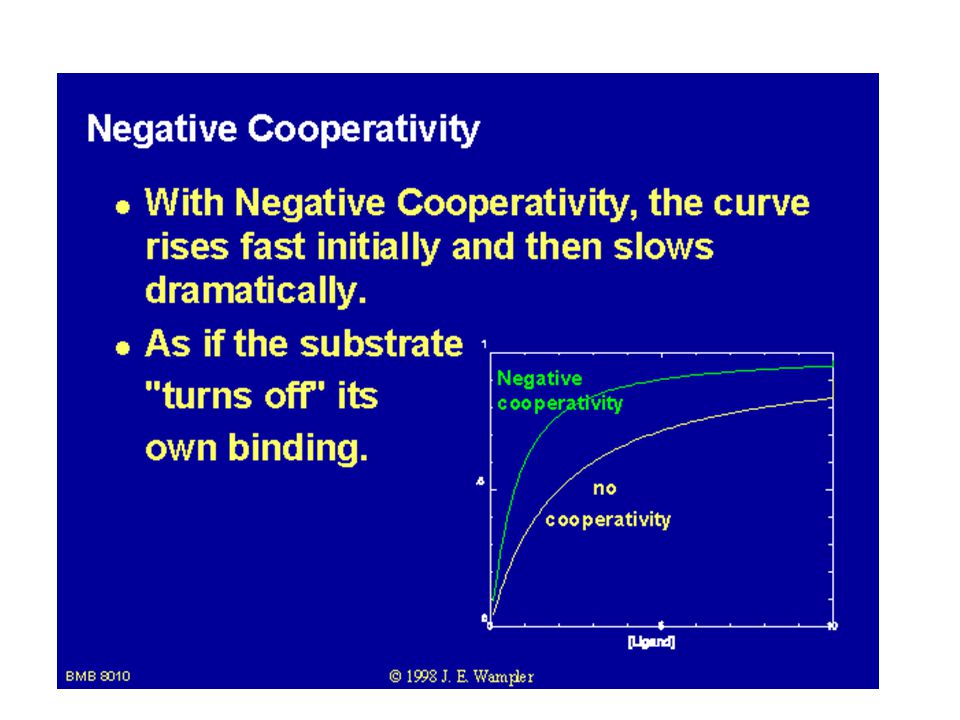
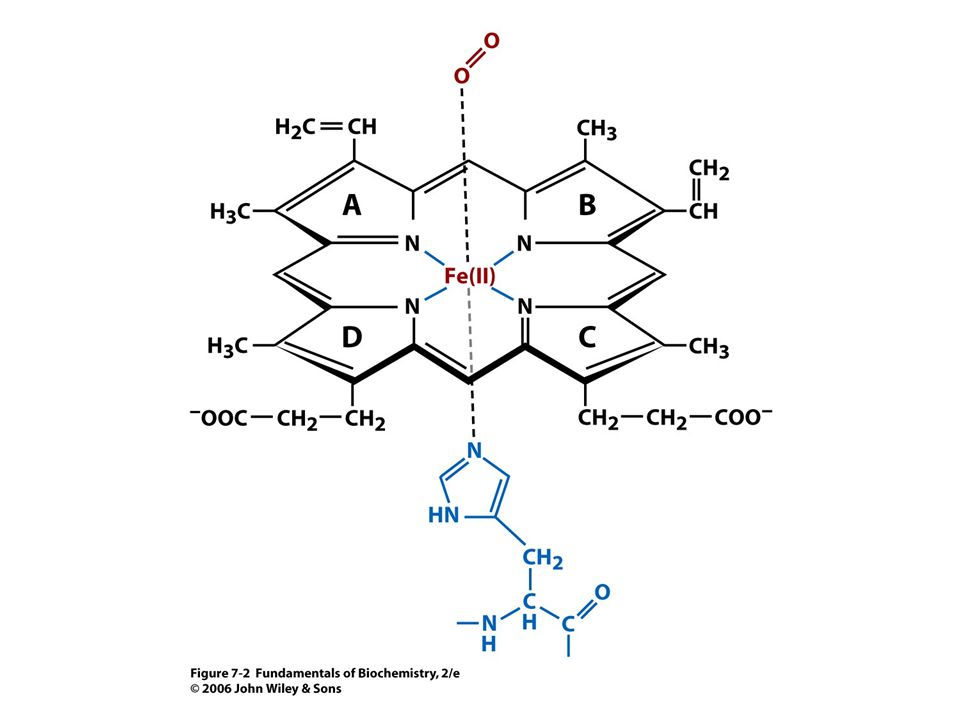
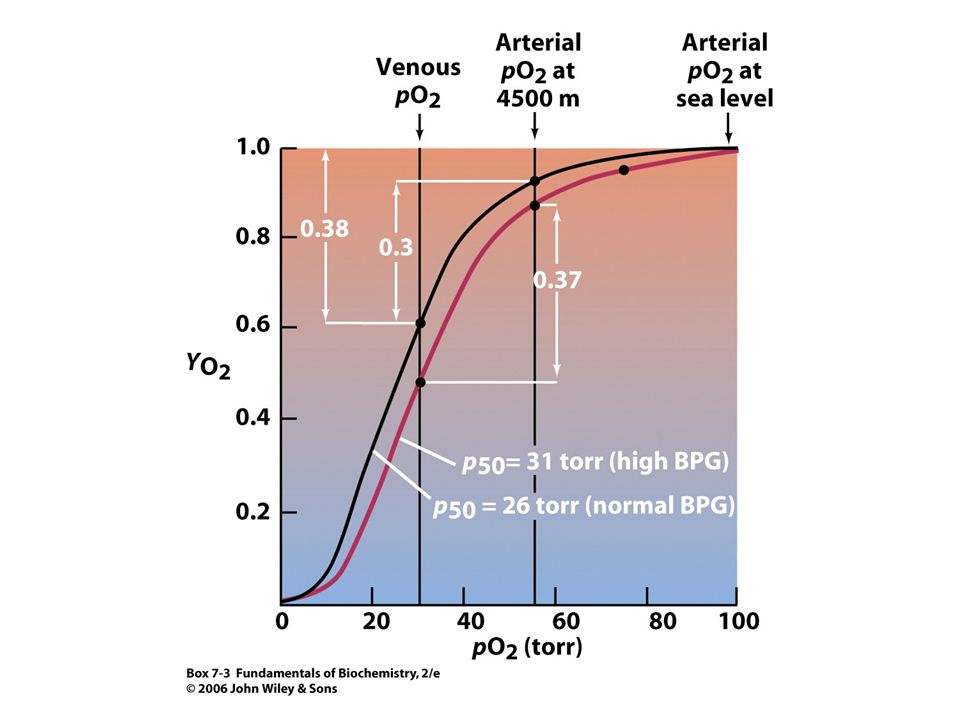
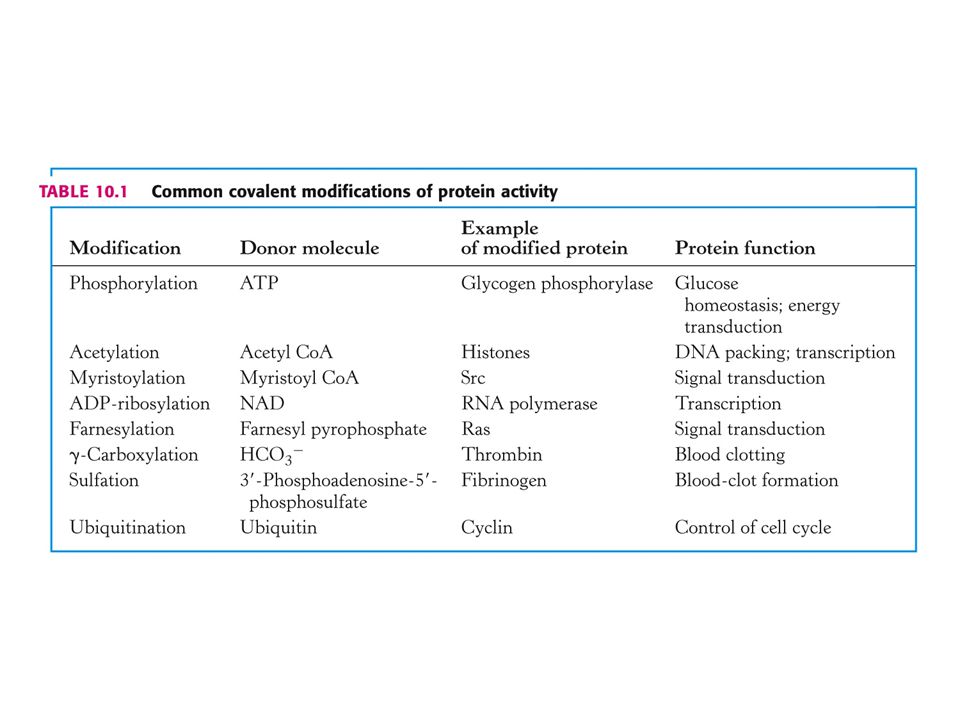




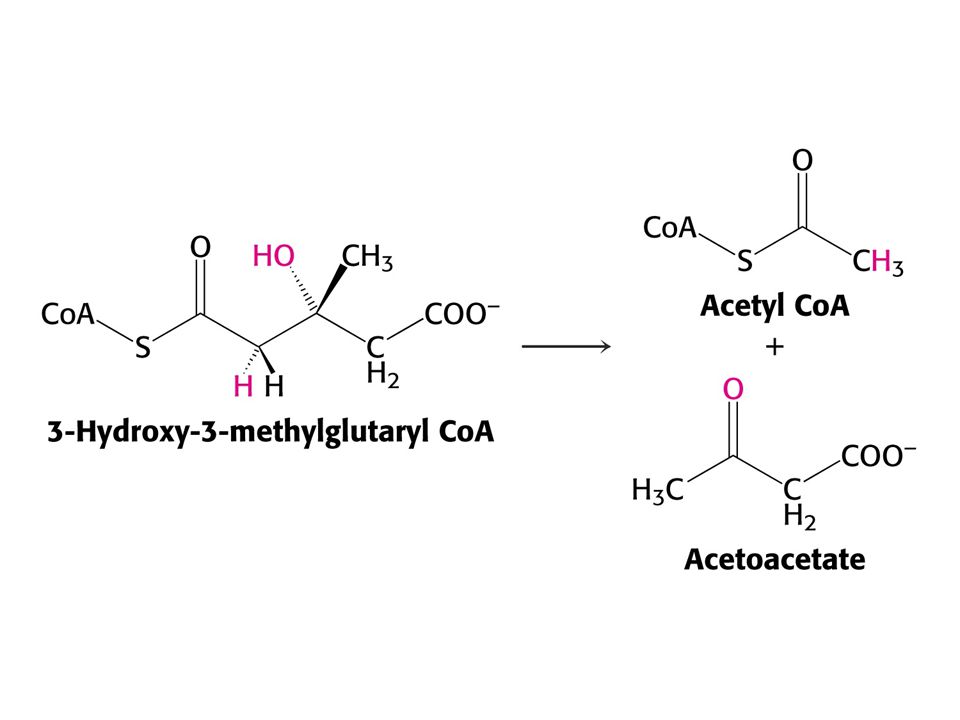

















 vylučované kůrou nadledvinek (aldosteron, kortisol); 2) vylučované pohlavními žlázami (progesteron, testosteron, estradiol)>")


