Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
T lymfocyty J. Ochotná

2
T lymfocyty T lymfocyty patří mezi buněčnou složku antigenně specifických mechanismů, při vývoji opouštějí kostní dřeň a migrují do thymu, kde dozrávají Existuje několik různých subpopulací T lymfocytů Podílejí se na regulaci imunitních dějů, při likvidaci virem infikovaných buněk či nádorových buněk Rozpoznávají antigen zpracovaný a prezentovaný APC; prostřednictvím TCR rozpoznávají komplex MHC gp-antigenní peptid T lymfocyty jsou po aktivaci stimulovány k pomnožení a diferenciaci v efektorové buňky, část se diferencuje v paměťové buňky

3
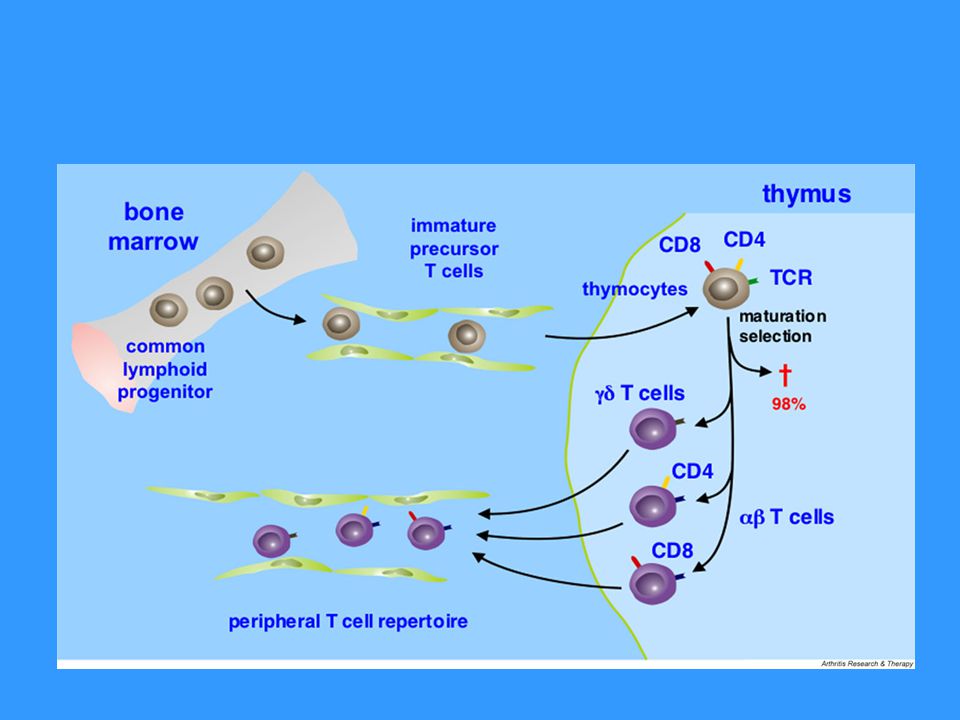
Vývoj T lymfocytů T lymfocyty vznikají v kostní dřeni a pak migrují do thymu, kde dozrávají (abT lymfocyty), konečná diferenciace probíhá až po aktivaci antigenem zpracovaným a prezentovaným APC gdT lymfocyty se mohou vyvíjet i mimo thymus (jde o menšinovou populaci) Pluripotentní hematopoetická kmenová buňka Pro-thymocyt – dostává se z kostní dřeně do thymu, kde se začínají přeskupovat geny pro TCRb, exprimuje na svém povrchu tzv. pre-TCR (složený z b řetězce, pre-TCRa a CD3 komplexu), po té se začínají přeskupovat geny pro TCRa Kortikální thymocyt – exprimuje na svém povrchu TCR (složený z řetězců ab a CD3) a koreceptory CD4 a CD8; v této fázi dochází k selekci autoreaktivních buněk a buněk s nefunkčním TCR Medulární thymocyt (zralá T buňka) – zachovávají si expresi CD4 či CD8, usidlují se v sekundárních lymfoidních orgánech
, konečná diferenciace probíhá až po aktivaci antigenem zpracovaným a prezentovaným APC. gdT lymfocyty se mohou vyvíjet i mimo thymus (jde o menšinovou populaci) Pluripotentní hematopoetická kmenová buňka. Pro-thymocyt – dostává se z kostní dřeně do thymu, kde se začínají přeskupovat geny pro TCRb, exprimuje na svém povrchu tzv. pre-TCR (složený z b řetězce, pre-TCRa a CD3 komplexu), po té se začínají přeskupovat geny pro TCRa. Kortikální thymocyt – exprimuje na svém povrchu TCR (složený z řetězců ab a CD3) a koreceptory CD4 a CD8; v této fázi dochází k selekci autoreaktivních buněk a buněk s nefunkčním TCR. Medulární thymocyt (zralá T buňka) – zachovávají si expresi CD4 či CD8, usidlují se v sekundárních lymfoidních orgánech.")
4
Selekce T lymfocytů Negativní selekce – eliminace autoreaktivních buněk Pokud thymocyt váže svým TCR dostatečně silně komplex MHCgp s normálními peptidy (z autoantigenů) se kterými se setká na povrchu thymových buněk dostane signály vedoucí k apoptotické smrti buňky PAE buňky (peripherial antigen expressing cells) Pozitivní selekce – eliminace buněk s nefunkčním TCR Pozitivně selektovány jsou thymocyty, které s nízkou afinitou rozeznávají MHC gp, ty si pak zachovávají expresi CD4 či CD8 (váže-li příslušný TCR MHC gp I či II) – tyto zralé T bb. (medulární thymocyty) opouštějí thymus a usidlují se v sekundárních lymfoidních orgánech 98% pro-thymocytů v thymu během svého vývoje zahyne
 se kterými se setká na povrchu thymových buněk dostane signály vedoucí k apoptotické smrti buňky. PAE buňky (peripherial antigen expressing cells) Pozitivní selekce – eliminace buněk s nefunkčním TCR. Pozitivně selektovány jsou thymocyty, které s nízkou afinitou rozeznávají MHC gp, ty si pak zachovávají expresi CD4 či CD8 (váže-li příslušný TCR MHC gp I či II) – tyto zralé T bb. (medulární thymocyty) opouštějí thymus a usidlují se v sekundárních lymfoidních orgánech. 98% pro-thymocytů v thymu během svého vývoje zahyne.")
6
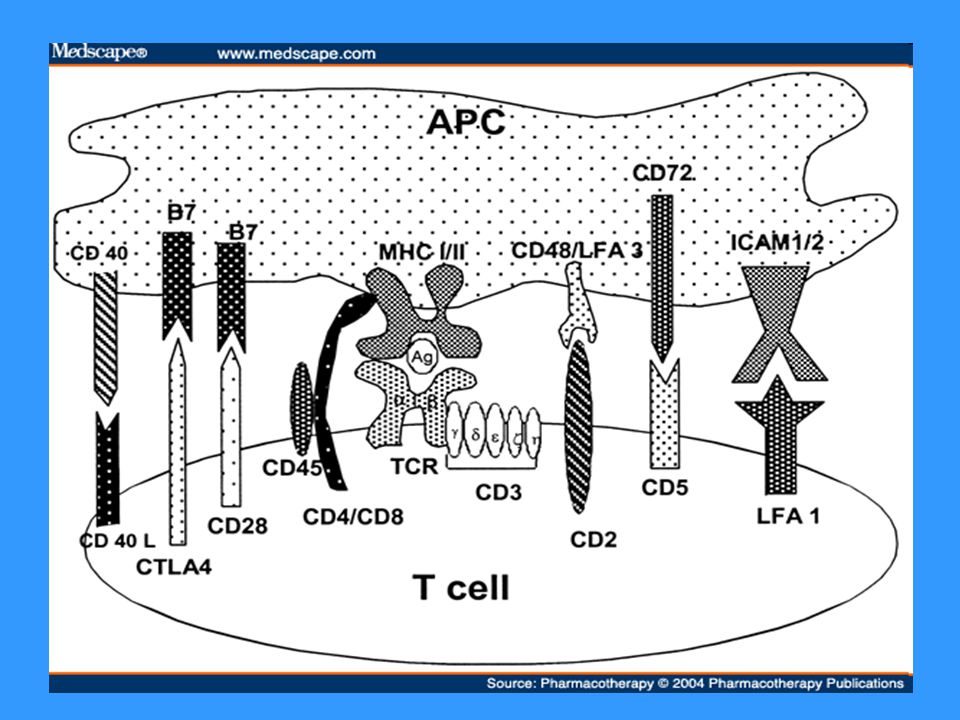
Povrchové znaky T lymfocytů
TCR – rozpoznává Ag peptid v komplexu s MHC gp CD3 – součást TCR, účast při přenosu signálu CD4 nebo CD8 – koreceptory, napomáhají k vazbě na MHC gp CD28 – kostimulační receptor; váže CD80, CD86 CTLA-4 (CD152) – inhibiční receptor; váže CD80, CD86
 – inhibiční receptor; váže CD80, CD86.")
8
Subpopulace T lymfocytů
ab-T lymfocyty – mají TCRab, většinový typ (95%), k vývoji potřebují thymus, rozeznávají antigeny v komplexu MHC gp-peptid gd-T lymfocyty – (5%) mohou se vyvíjet i mimo thymus, některé jsou schopny rozpoznat nativní Ag, uplatňují se při obraně kůže a sliznic Intraepiteliální T lymfocyty NK-T lymfocyty – rozeznávají komplexy CD1 molekul s lipidy
, k vývoji potřebují thymus, rozeznávají antigeny v komplexu MHC gp-peptid. gd-T lymfocyty – (5%) mohou se vyvíjet i mimo thymus, některé jsou schopny rozpoznat nativní Ag, uplatňují se při obraně kůže a sliznic. Intraepiteliální T lymfocyty. NK-T lymfocyty – rozeznávají komplexy CD1 molekul s lipidy.")
9
ab-T lymfocyty TH3 – TGFb
Exprimující koreceptor CD4 (koreceptor pro MHC gp II) Jde o prekurzory pomocných T buněk (TH), ty lze rozdělit podle produkce cytokinů na : TH0 – produkují směs cytokinů jako TH1 a TH2 TH1 – IL-2, IFNg (pomoc makrofágům) TH2 – IL-4, IL-5, IL-6, IL-10 (pomoc B lymfocytům) TH3 – TGFb Treg – regulační T lymfocyty, potlačují aktivitu TH1 a částečně fungují jako TS, potlačování autoreaktivních klonů T lymfocytů; produkují IL-10 a TGFb
 Jde o prekurzory pomocných T buněk (TH), ty lze rozdělit podle produkce cytokinů na : TH0 – produkují směs cytokinů jako TH1 a TH2 TH1 – IL-2, IFNg (pomoc makrofágům) TH2 – IL-4, IL-5, IL-6, IL-10 (pomoc B lymfocytům) TH3 – TGFb. Treg – regulační T lymfocyty, potlačují aktivitu TH1 a částečně fungují jako TS, potlačování autoreaktivních klonů T lymfocytů; produkují IL-10 a TGFb.")
10
ab-T lymfocyty Exprimující koreceptor CD8 (koreceptor pro MHC gp I) Jde o prekurzory cytotoxických T buněk (TC), či supresorových T buněk (TS) TC – rozeznávají buňky napadené viry či jinými intracelulárními parazity a některé nádorové buňky TS – potlačují funkce ostatních lymfocytů
 Jde o prekurzory cytotoxických T buněk (TC), či supresorových T buněk (TS) TC – rozeznávají buňky napadené viry či jinými intracelulárními parazity a některé nádorové buňky. TS – potlačují funkce ostatních lymfocytů.")
11
TCR TCR (T cell receptor) se skládá z modulu rozeznávajícího Ag a asociovaného CD3 komplexu, který je nezbytný pro přenos signálu, je propojen s PTK (Src) Modul rozeznávající Ag je tvořen 2 řetězci a a b (g,d), N-terminální části tvoří vazebné místo pro Ag T lymfocyty (ab) rozeznávají komplex MHC gp-Ag peptid na povrchu APC, při rozpoznání spolupracuje TCR s koreceptory CD4 (pomáhá vázat MHC gp II) nebo CD8 (pomáhá vázat MHC gp I) Aktivace přes TCR a CD28 vede k proliferaci a diferenciaci v efektorové buňky CD28 – receptor kostimulačního signálu, váže ligandy CD80 a CD86, které jsou na povrchu APC
 se skládá z modulu rozeznávajícího Ag a asociovaného CD3 komplexu, který je nezbytný pro přenos signálu, je propojen s PTK (Src) Modul rozeznávající Ag je tvořen 2 řetězci a a b (g,d), N-terminální části tvoří vazebné místo pro Ag. T lymfocyty (ab) rozeznávají komplex MHC gp-Ag peptid na povrchu APC, při rozpoznání spolupracuje TCR s koreceptory CD4 (pomáhá vázat MHC gp II) nebo CD8 (pomáhá vázat MHC gp I) Aktivace přes TCR a CD28 vede k proliferaci a diferenciaci v efektorové buňky. CD28 – receptor kostimulačního signálu, váže ligandy CD80 a CD86, které jsou na povrchu APC.")
12
Spolupráce TCR s koreceptory CD4 a CD8

13
Vznik TCR Obdoba s tvorbou BCR
Řetězce b a d - odpovídají genovému komplexu IgH imunoglobulinů V, D, J, C segmenty Řetězce a a g - odpovídají genům pro L řetězce imunoglobulinů V, J, C segmenty Přeskupování genů probíhá podobně jako u BCR a provádějí je shodné rekombinázy u genů pro TCR nedochází k somatickým mutacím a afinitní maturaci

14
TH1 imunitní odpověď

15
TH1 imunitní odpověď – zánětlivá reakce
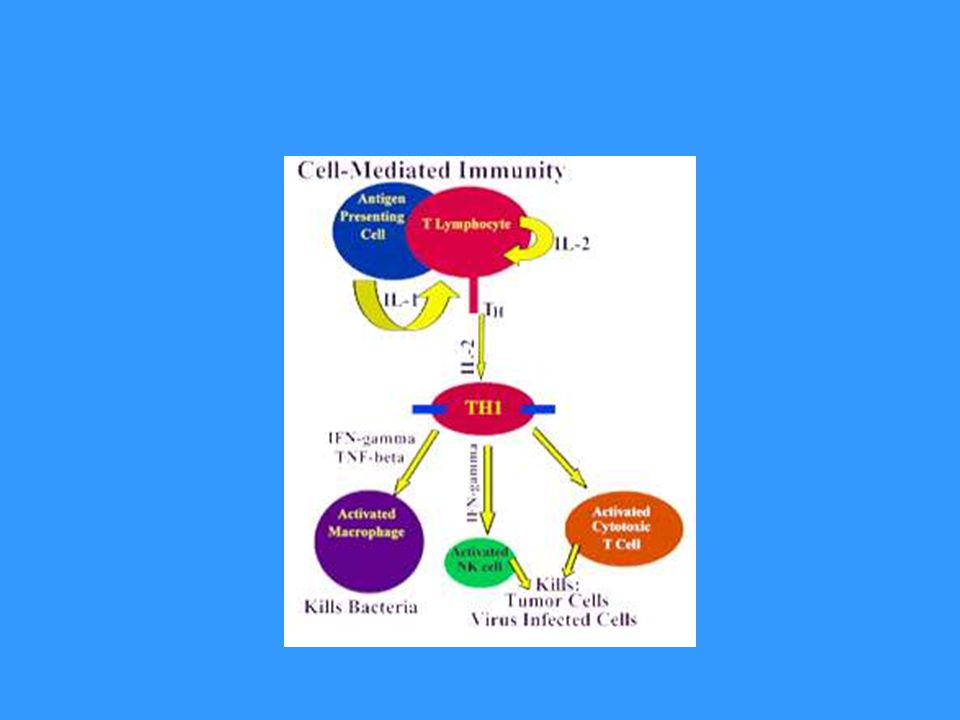
Základní funkcí TH1 buněk je spolupráce s makrofágy a jejich přeměna v aktivované, které jsou schopny produkovat NO, pomocí kterého likvidují své intracelulární parazity Pro přeměnu makrofágů v aktivované jsou nezbytné cytokiny (IFNg) produkované TH1 buňkami Aktivované makrofágy secernují některé cytokiny ( IL-1, TNF…), které napomáhají ke stimulaci T buněk a stimulují lokální zánět, který napomáhá potlačení infekce Vzájemné působení TH1 buněk a makrofágů je základním mechanismem imunopatologické reakce opožděného typu (DTH- delayed type hypersensitivity)
 produkované TH1 buňkami. Aktivované makrofágy secernují některé cytokiny ( IL-1, TNF…), které napomáhají ke stimulaci T buněk a stimulují lokální zánět, který napomáhá potlačení infekce. Vzájemné působení TH1 buněk a makrofágů je základním mechanismem imunopatologické reakce opožděného typu (DTH- delayed type hypersensitivity)")
16
Infikovaný makrofág produkuje fragmenty proteinů pocházející z intracelulárního parazita, některé jsou prezentovány na jeho povrchu pomocí MHC gp II Makrofágy a dendritické buňky stimulované některými mikroorganismy produkují IL-12 Prekurzor TH , který rozpozná infikovaný makrofág a obdrží signály přes TCR, CD 28 a receptor pro IL-12 a další adhezivní a signalizační molekuly proliferuje a diferencuje se na efektorové TH1 buňky, které produkují IFNg a IL-2. IFNg podporuje přeměnu makrofágů v aktivované IL-2 je autokrinní růstový faktor TH1 lymfocytů.

17
Interakce mezi APC a prekurzorem TH

18
Diferenciace TH0

19
Kostimulační signál

21
TH2 imunitní odpověď

22
TH2 imunitní odpověď – pomoc B lymfocytům
Základní funkcí TH2 buněk je spolupráce s B lymfocyty (které byly stimulovány Ag) prostřednictvím cytokinů (IL-4, IL-5, IL-6) a přímého mezibuněčného kontaktu Pro stimulaci B lymfocytů je většinou potřeba spolupráce mezi APC → TH2 buňkou → B lymfocytem V případě tzv. minimálního modelu, pokud se z B lymfocytu stane dobrá APC (CD80, CD86), stačí spolupráce mezi TH2 buňkou → B lymfocytem
 prostřednictvím cytokinů (IL-4, IL-5, IL-6) a přímého mezibuněčného kontaktu. Pro stimulaci B lymfocytů je většinou potřeba spolupráce mezi APC → TH2 buňkou → B lymfocytem. V případě tzv. minimálního modelu, pokud se z B lymfocytu stane dobrá APC (CD80, CD86), stačí spolupráce mezi TH2 buňkou → B lymfocytem.")
23
Prekurzor TH , který rozpozná infikovaný makrofág a dostane signály přes TCR, CD 28, receptor pro IL-4 a receptor pro IL-2 a další adhezivní a signalizační molekuly proliferuje a diferencuje se na efektorové TH2, které poskytují B lymfocytům pomocné signály pomocí sekretovaných cytokinů IL-4, IL-5, IL-6 a prostřednictvím adhezivních molekul CD 40L, které se váží se na kostimulační receptor B lymfocytů CD 40 Interakce mezi CD40 (B lymfocyt) a CD40L (TH2 buňka) je nezbytná pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových buněk IL-4, IL-5, IL-6 : stimulace B lymfocytů
 a CD40L (TH2 buňka) je nezbytná pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových buněk. IL-4, IL-5, IL-6 : stimulace B lymfocytů.")
25
Pomoc B lymfocytům Specifická přímá pomoc B lymfocytům:
TH2 lymfocyt poskytuje pomoc B lymfocytům, které byly stimulovány stejným Ag, který vyvolal vznik TH2 Ke stimulaci sekrece cytokinů TH2 buňce stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný) Jeden klon TH2 buněk může poskytovat specifickou pomoc B lymfocytům různých specifit (musí prezentovat příslušné Ag peptidy pomocí MHC gp II, které rozpoznáváTCR)
 Jeden klon TH2 buněk může poskytovat specifickou pomoc B lymfocytům různých specifit (musí prezentovat příslušné Ag peptidy pomocí MHC gp II, které rozpoznáváTCR)")
26
Přímá pomoc B lymfocytům

27
Pomoc B lymfocytům Nepřímá pomoc B lymfocytům („bystander help“):
TH2 lymfocyt poskytuje pomoc B lymfocytům, které byly stimulovány jiným Ag, než který vyvolal vznik TH2 Kontakt mezi TH2 buňkou → B lymfocytem prostřednictvím adhezivních molekul, sekrece cytokinů, vazba CD40-CD40L Nebezpečí aktivace autoreaktivních B lymfocytů

28
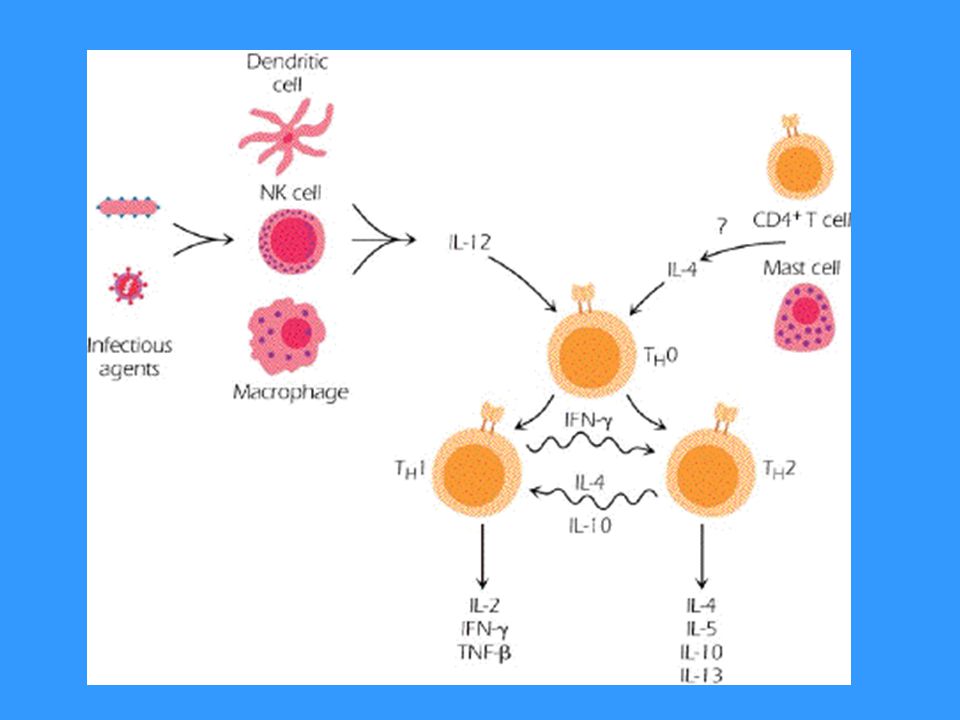
Vzájemná regulace aktivit TH1 versus TH2
Zda se prekurzorové TH lymfocyty budou vyvíjet v TH1 či TH2 rozhodne poměr cytokinů IL-12 a IL-4 IL-12 je produkován makrofágy a dendritickými buňkami stimulovanými některými mikroorganismy IL-4 je produkován bazofily a mastocyty Cytokiny produkované TH1 (hlavně IFNg) inhibují vývoj TH2 a stimulují vývoj TH1 (IL-2 stimuluje i TH2) Cytokiny produkované TH2 (IL-4, IL-10) inhibují vývoj TH1 a stimulují vývoj TH2 Vývoj TH3 je stimulován specifickým cytokinovým prostředím IL-4, IL-10, TGFb; TH3 produkují TGFb, spolupracují s B lymfocyty v MALT
 inhibují vývoj TH2 a stimulují vývoj TH1 (IL-2 stimuluje i TH2) Cytokiny produkované TH2 (IL-4, IL-10) inhibují vývoj TH1 a stimulují vývoj TH2. Vývoj TH3 je stimulován specifickým cytokinovým prostředím IL-4, IL-10, TGFb; TH3 produkují TGFb, spolupracují s B lymfocyty v MALT.")
30
Imunitní reakce založená na TC

31
Stimulace cytotoxických T lymfocytů
TC rozeznávají buňky infikované viry či jinými intracelulárními parazity a některé nádorové buňky Prekurzor TC , který rozpozná komplex MHC gp I- antigenní peptid na povrchu APC přes TCR a dostane signály přes CD 28 proliferuje a diferencuje se na klon zralých efektorových cytotoxických buněk (CTL); toto probíhá za pomoci TH1 lymfocytů produkujících IL-2 Efektorové TC jsou rozneseny krevním oběhem do tkání, k aktivaci cytotoxických mechanismů stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný)
; toto probíhá za pomoci TH1 lymfocytů produkujících IL-2. Efektorové TC jsou rozneseny krevním oběhem do tkání, k aktivaci cytotoxických mechanismů stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný)")
32
Profesionální APC jsou dendritické buňky nebo makrofágy, které jsou infikovány virem, nebo pohltili antigeny z odumřelé virem infikované, nádorové nebo stresované buňky Aby APC mohla aktivovat prekurzor TC ,musí být sama stimulována kontaktem s TH buňkami přes CD 40, poté začne dendritická buňka exprimovat CD 80, CD86 a sekretovat cytokiny (IL-1, IL-12) = přeměna klidové APC v aktivovanou
 = přeměna klidové APC v aktivovanou.")
33
Efektorové mechanismy TC
Cytotoxická granula obsahující perforin a granzymy (perforin vytváří póry v cytoplazmatické membráně napadené buňky,v některých případech může dojít k osmotické lýze napadené buňky, vytvořenými póry se do buňky dostávají granzymy, které aktivují caspázy, což vede k apoptóze napadené buňky Fas-ligand (FasL) – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. (i na povrchu TC) TNFb
 – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. (i na povrchu TC) TNFb.")
35
Rozdílná aktivace Tc a NK

36
Imunitní odpověď založená na protilátkách

37
Protilátková reakce vyvolaná:
Antigeny nezávislými na T lymfocytech Vyvolávají převážně tvorbu IgM protilátek Jedná se zvláště o bakteriální polysacharidy, lipopolysacharidy a polymerní formy proteinů Antigeny závislými na T lymfocytech Reakce na tyto Ag probíhá ve dvou fázích – primární a sekundární odpověď Tyto reakce mohou být od sebe odděleny či mohou na sebe bezprostředně navazovat Vyvolávají tvorbu paměťových buněk a vznik vysokoafinních protilátek

38
T-nezávislá a T-závislá imunitní odpověď

39
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Primární fáze protilátkové reakce - při prvním setkání s Ag Probíhá v sekundárních lymfatických orgánech Stimulace B lymfocytu vazbou Ag na BCR Pohlcení Ag APC a jeho prezentace prostřednictvím MHC gp II prekurzorům TH buněk → vznik klonu antigenně specifických TH2 buněk, které poskytují pomoc příslušným B lymfocytům, což vede k jejich proliferaci, diferenciaci na plazmatické bb (produkují Ab) a na paměťové bb Plazmatické bb jsou rozneseny oběhovým systémem do organismu (zvláště kostní dřeně) Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům
 a na paměťové bb. Plazmatické bb jsou rozneseny oběhovým systémem do organismu (zvláště kostní dřeně) Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy. Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům.")
40
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Sekundární fáze protilátkové reakce – po rozpoznání Ag na FDC Vznikne-li dostatečné množství imunokomplexů na FDC a jsou-li rozpoznány B lymfocyty Germinální reakce - pod vlivem signálů od FDC (Ag) a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí
 a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek. Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí.")
41
V sekundární fázi imunitní reakce vznikají protilátky s vyšší afinitou k Ag a s jinými efektorovými vlastnostmi závislými na izotypu, vznikají také paměťové bb pro další setkání s Ag Protilátky v organismu po primární infekci přetrvávají po dlouhou dobu Kontakt mezi CD40 (B lymfocyt) a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.
 a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.")
43
Protilátková odpověď Primární odpověď
- následuje po prvním kontaktu s antigenem, kdy ještě nejsou přítomny paměťové buňky - tvoří se převážně nízkoafinní IgM, postupně se tvoří ostatní izotypy s vyšší afinitou Sekundární odpověď - následuje po opakovaných setkáních se stejným antigenem, jsou již přítomny paměťové buňky - hladina IgM protilátek je nižší, rychleji se tvoří ostatní izotypy, protilátky mají vyšší afinitu k antigenu, protože jsou aktivovány paměťové buňky

44
Dynamika tvorby protilátek při primární a sekundární odpovědi

45
Fyziologické imunitní regulační mechanismy

46
Regulace antigenem Vyvolání a vyhasnutí imunitní odpovědi
Afinitní maturace B lymfocytů Udržení imunologické paměti Antigenní kompetice Prahová hustota komplexu MHC gp II-Ag na APC

47
Regulace antagonistickými peptidy
Agonista - antigenní peptid, který vyvolá plnohodnotnou odpověď T buněk (proliferace, diferenciace TH či TC a stimulace efektorových funkcí) Antagonista - (částečný agonista) peptid strukturně podobný antigennímu peptidu, který vyvolává kvalitativně jinou odpověď T lymfocytů (např. produkce jen některých cytokinů, nebo anergie...) Negativní signály vyvolané antagonistou mohou překonat pozitivní signály vyvolané agonistou, který je v organismu v nadbytku, toho využívají některé mikroorganismy
 Antagonista - (částečný agonista) peptid strukturně podobný antigennímu peptidu, který vyvolává kvalitativně jinou odpověď T lymfocytů (např. produkce jen některých cytokinů, nebo anergie...) Negativní signály vyvolané antagonistou mohou překonat pozitivní signály vyvolané agonistou, který je v organismu v nadbytku, toho využívají některé mikroorganismy.")
48
Regulace protilátkami
Protilátky kompetují s BCR o antigen (negativní regulátor stimulace B lymfocytů) Imunokomplexy s IgG se váží na B lymfocyt na BCR a FcgR, důsledkem je blokování aktivace B lymfocytů Zatím je nejasný význam regulace pomocí idiotypové sítě
 Imunokomplexy s IgG se váží na B lymfocyt na BCR a FcgR, důsledkem je blokování aktivace B lymfocytů. Zatím je nejasný význam regulace pomocí idiotypové sítě.")
49
Regulace cytokiny a mezibuněčným kontaktem
Interakce APC - T lymfocyt Interakce TH1 – makrofág Interakce TH2 – B lymfocyt Vzájemná regulace aktivit TH1 versus TH2 Vývoj subpopulací leukocytů Negativní regulace efektorových lymfocytů: CTLA-4 inhibiční receptor T lymfocytů, váže ligandy CD80 a CD86 Inhibiční receptory NK buněk Sebedestrukční interakce apoptotického receptoru Fas s ligandem FasL na povrchu aktivovaných T lymfocytů

50
Příklad regulace cytokiny při zánětlivé odpovědi

51
Suprese zprostředkovaná T lymfocyty
Vzájemná negativní interakce TH1 a TH2 zprostředkovaná cytokiny (TH2 lymfocyty produkují IL-4 a IL-10, které potlačují imunitní reakce založené na TH1 buňkách) CD 8+ TS - supresorové T lymfocyty se zatím nepodařilo izolovat jako samostatnou subpopulaci (částečně identické s TC ) - negativně regulují aktivaci jiných T buněk Klonální eliminace či anergizace T lymfocytů po styku s antigenem na povrchu jiných buněk, než APC (chybí kostimulační signály Regulační T lymfocyty (Tr1 CD 4+; podobné TH3) pomáhají udržet toleranci k autoantigenům
 CD 8+ TS - supresorové T lymfocyty se zatím nepodařilo izolovat jako samostatnou subpopulaci (částečně identické s TC ) - negativně regulují aktivaci jiných T buněk. Klonální eliminace či anergizace T lymfocytů po styku s antigenem na povrchu jiných buněk, než APC (chybí kostimulační signály. Regulační T lymfocyty (Tr1 CD 4+; podobné TH3) pomáhají udržet toleranci k autoantigenům.")
52
Neuroendokrinní regulace
Některé neurotransmitery působí na leukocyty (noradrenalin) Přímý kontakt volných nervových zakončení s mastocyty (bolest při degranulaci mastocytů) Některé endokrinní hormony působí na leukocyty (kortikosteroidy, růstový hormon, thyroxin, endorfiny...) Leukocyty produkují řadu hormonů (endorfiny, ACTH, TSH, růstový hormon...) Některé cytokiny působí na nervový systém (IL-1, IL-6, LIF, TNF) Vliv emocionálního stresu na imunitní systém
 Přímý kontakt volných nervových zakončení s mastocyty (bolest při degranulaci mastocytů) Některé endokrinní hormony působí na leukocyty (kortikosteroidy, růstový hormon, thyroxin, endorfiny...) Leukocyty produkují řadu hormonů (endorfiny, ACTH, TSH, růstový hormon...) Některé cytokiny působí na nervový systém (IL-1, IL-6, LIF, TNF) Vliv emocionálního stresu na imunitní systém.")
53
Faktory ovlivňující výsledek imunitní odpovědi
Tentýž antigen může navodit aktivní imunitní odpověď nebo stav aktivní tolerance, výsledek odpovědi závisí na mnoha faktorech: Stavu imunitního systému Vlastnostech antigenu Dávce antigenu Způsobu podání

54
Cytokiny ( tkáňové hormony )
")
55
Cytokiny Regulační proteiny a glykoproteiny produkované leukocyty i jinými buňkami Základní regulátory imunitního systému Uplatňují se i mimo imunitní systém (angiogeneze, regenerace tkání, kancerogeneze, ovlivnění řady mozkových funkcí,embryoníální vývoj…) Cytokiny - sekretované - membránové (zajištěno lokální působení; CD 80, CD86, CD40L, FasL..)
 Cytokiny - sekretované. - membránové (zajištěno lokální působení; CD 80, CD86, CD40L, FasL..)")
56
Pleiotropní účinek Působí v kaskádě Cytokinová síť Cytokinový systém je redundantní Působení cytokinů- autokrinní - parakrinní - endokrinní Jsou označovány jako interleukiny (vyjímka: TNF, lymfotoxin, TGF, interferony, CSF a růstové faktory)
")
57
B lymfocyt komunikuje s makrofágem a T lymfocytem pomocí cytokinů

58
Přehled cytokinů interleukiny ( IL-1 až IL-23 )
chemokiny ( IL-8 a příbuzné molekuly ) interferony ( IFN-, -, - ) transformující růstové faktory ( TGF,TGF ) faktory stimulující kolonie ( G-CSF, M-CSF, GM-CSF ) faktory nekrotizující nádory ( TNF-, lymfotoxin ) jiné růstové faktory (SCF, EPO, FGF, NGF,LIF )
 interferony ( IFN-, -, - ) transformující růstové faktory ( TGF,TGF ) faktory stimulující kolonie ( G-CSF, M-CSF, GM-CSF ) faktory nekrotizující nádory ( TNF-, lymfotoxin ) jiné růstové faktory (SCF, EPO, FGF, NGF,LIF )")
59
Rozdělení cytokinů podle funkce
Prozánětlivé cytokiny (IL-1 a , IL-6, 8, 12, 18, TNF) Protizánětlivé cytokiny (IL-1Ra, IL-4, IL-10, TGF ) Cytokiny s aktivitou růstových faktorů hemopoetických bb. ( IL-2, 3, 4, 5, 6, 7, 9, 11, 14, 15, CSF, SCF, LIF, EPO ) Cytokiny uplatňující se v humorální imunitě TH2 ( IL-4, 5, 9, 13 ) Cytokiny uplatňující se v buněčně zprostředkované imunitě TH1 ( IL- 2, 12, IFN, GM-CSF, lymfotoxin ) Cytokiny s antivirovým účinkem ( IFN-, IFN-, IFN- )
 Protizánětlivé cytokiny (IL-1Ra, IL-4, IL-10, TGF ) Cytokiny s aktivitou růstových faktorů hemopoetických bb. ( IL-2, 3, 4, 5, 6, 7, 9, 11, 14, 15, CSF, SCF, LIF, EPO ) Cytokiny uplatňující se v humorální imunitě TH2 ( IL-4, 5, 9, 13 ) Cytokiny uplatňující se v buněčně zprostředkované imunitě TH1 ( IL- 2, 12, IFN, GM-CSF, lymfotoxin ) Cytokiny s antivirovým účinkem ( IFN-, IFN-, IFN- )")
60
Receptory cytokinů Jsou složeny ze 2 či 3 podjednotek
Jedna váže cytokin, další asociovány s cytoplazmatickými signalizačními molekulami (protein-kinázami) Signalizační podjednotka bývá sdílena několika různými cytokinovými receptory – tzv receptorové rodiny Signalizace přes tyto receptory může vést k proliferaci, diferenciaci, aktivaci efektorových mechanismů či zablokování buněčného cyklu a indukce apoptózy
 Signalizační podjednotka bývá sdílena několika různými cytokinovými receptory – tzv receptorové rodiny. Signalizace přes tyto receptory může vést k proliferaci, diferenciaci, aktivaci efektorových mechanismů či zablokování buněčného cyklu a indukce apoptózy.")
Podobné prezentace













 Mgr. Martin Šmíd.>")




>")




