Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Chemické smysly Chuť Čich Vnitřní chemorecepce Obecný chemický smysl

2
Chuť Vnímá chemické látky rozpuštěné ve vodě
Receptory shloučeny v chuťových pohárcích v papilách Na jazyku Volně na patře Člk – 3 typy papil Houbovitá papila (1-5 pohárků) Listovitá papila (tisíce pohárků) Hrazená papila (velké, okrouhlé, tisíce pohárků)
 Listovitá papila (tisíce pohárků) Hrazená papila (velké, okrouhlé, tisíce pohárků)")
3
Chuťový pohárek obratlovců Pór Pokožka Mikrovilli
Místo trandukce- Podpůrné a bazální buňky nové receptory a trofická funkce Receptory Nerv Aferentní vlákna

4
Základní modality chuti
Specifická chuť vzniká kombinací základních modalit Hořko Slano Kyselo Sladko Topografie sladko/slano – špička jazyka, kyselo – laterální části, hořko – kořen jazyka

5
Charakteristika Hořko spojeno s nebezpečnými stimuly Kyselo – kyseliny
Chemicky heterogenní látky Sladko – nejsladší pro lidi jsou proteiny thaumatin a monellin Strukturálně odlišné krát sladší než glukosa Kyselo – kyseliny Slano

6
Chemická transdukce a buněčná biochemie
Odpovědi chemoreceptorů extrémně variabilní Od přímého působení na iontové kanály Přes zapojení vnitrobuněčných přenašečů Bohatá škála neuronálních odpovědí Využitelné i na jiných úrovních komunikace neuronů

7
Transdukce chuťových stimulů
Molekuly slané a kyselé chuti Přímo otvírají iontové kanály receptorů Molekuly sladké a hořké chuti Působí prostřednictvím vnitrobuněčného přenašeče Existují výjimky pro některé hořké chuti Transdukce aminokyselin – přímo i přes přenašeče

8
Slaná a kyselá transdukce
Kyseliny pronikají membránou chuťových receptorů přímo bez intervence receptorů Slano vzniká průchodem iontů přes napěťově nezávislé kationtové kanály Nevyžaduje přítomnost specifických receptorů

9
A B Transdukce na slaných a kyselých receptorech H+ Uzavření kanálu
V klidu A K+ Hromadění K+ v ICT Depolarizace B Na+ Na+ Na+ Na+ Na+ V klidu Na+ Na+ Depolarizace

10
Sladká transdukce Navrženy dva mechanismy sladké transdukce
Otevření napěťově nezávislých Na+ kanálů Zavření napěťově nezávislých K+ kanálů Není jasné, zda oba mechanismy mohou existovat na jednom receptoru

11
Hořká transdukce Působeni Mechanismus Látka Přímý vliv na kanály
Receptory řízená iontová pumpa Receptory aktivovaný G protein (gustducin) Uzavření K+ kanálů Aktivace Cl- pumpy Aktivace PIP a IP3,zvýšení Ca+ a uzavření K+ kanálů Inhibice PDE a snížení cAMP hladiny Barium Quinin Strychnin Denatonium
 Uzavření K+ kanálů. Aktivace Cl- pumpy. Aktivace PIP a IP3,zvýšení Ca+ a uzavření K+ kanálů. Inhibice PDE a snížení cAMP hladiny. Barium. Quinin. Strychnin. Denatonium.")
12
uzavření kanálu – depolariz. IP3 Vnitřní membrána Ca++ zásoby
Hořký ligand Fosfolipáza C (PDC) Receptor G protein (gustducin) PIP2 DAG B – uzavření kanálu – depolariz. IP3 K+ Vnitřní membrána Ca++ zásoby Proteinkináza C B Ca++ uvolnění DAG=diacylglycerol PIP=fosfatidylinositoldifosfát A - uvolnění mediátoru bez depolarizace
 Receptor. G protein (gustducin) PIP2. DAG. B – uzavření kanálu – depolariz. IP3. K+ Vnitřní membrána Ca++ zásoby. Proteinkináza C. B. Ca++ uvolnění. DAG=diacylglycerol. PIP=fosfatidylinositoldifosfát. A - uvolnění mediátoru. bez depolarizace.")
13
Chuťové dráhy Chuťové info se nekříží
První synapse na chuťových receptorech Jedno aferentní vlákno inervuje více receptorů Vzniká komplexní receptivní pole Pole se překrývají Aferentní vlákna vstupují hlavovými VII, IX a X nervy Druhá synapse je v chuťovém jádře (solitary nuclear complex a parabrachiální jádro mostu) Pak v thalamu Kůra Modality segregovány v průběhu všemi analyzačními centry
 Pak v thalamu. Kůra. Modality segregovány v průběhu všemi analyzačními centry.")
14
Chuťový systém člověka
Somatosensorická kůra Chuťová oblast I Thalamus Insula Chuťová oblast II Chuťové jádro Prodloužená mícha Chuťový systém člověka

15
Parabrachiální jádro Tektum Kůra Mozeček Mozeček Komora Most Sacharin
NaCl HCl Quinin

16
Chuťové kódování 4 separátní kanály pro hlavní chutě existují i v mozkovém kmeni Separace však není absolutní Většina receptorů odpovídá na více než jednu látku Ve směsích jedna látka může modulovat působení látky jiné

17
Chemosensila bezobratlých
Otvůrek na vrchu Sensila Dendrity receptorů Kutikula Dendrity Receptorová lymfa Sůl Cukr Voda Specifické molekuly Podpůrná buňka Smyslový neuron Axony

18
Čich Fylogeneticky nejstarší Emoce a paměť
Receptory odpovídají na diskrétní těkavé látky Nosní sliznice v nosní dutině Receptory Bb podpůrné a bazální

19
Receptory Bipolární neurony
Dendrity tvoří cilie, ty zanořeny v mukoze nosní dutiny Axony končí v čichovém laloku Zpočátku nemyelinizované, později tvoří svazky obalené myelinem, pronikají cribriformní destičkou Receptory savců kontinuálně obnovovány Problematika prorůstání axonů

20
Čichové receptory obratlovců
Nerv Receptory Podůrné buňky Cilie Sliznice

21
Vůně Komplexní směsi těkavých látek
Kvalita dána přítomností různých látek a jejich poměry Lipofilní látky málo rozpustné ve vodě Velká variabilita struktur a funkčních skupin Společný jmenovatel – nízká molekulová hmnotnost

22
Transdukce Vůně nejprve absorbovány mukozou
Přítomnost odorant binding proteinu Různé teorie o jeho funkci Interakce vůně s receptorem – depolarizace G protein aktivace adenylylcyklázy nebo IP3 Na+ Kanály přímo kontrolovány cAMP Ca+ + - IP3

23
Diversifikace vnitřních přenašečů
Vůně mohou aktivovat různé přenašeče Vede se diskuze, zda jeden receptor může využívat alternativních drah k ovládání iontových kanálů Receptory nedávno identifikovány 7 transmembránových smyček Spřažené s G proteinem Extrémně citlivé Jeden receptor exprimuje jeden gen

24
A B Čichová transdukce obratlovců Vůně Receptor cAMP G protein ATP
Adenylylcykláza Na+ Receptor Na+ cAMP G protein ATP Vůně B Fosfolipáza C Receptor Ca++ PIP2 Ca++ IP3 G protein

25
Ligand Receptor Adenylyl cykláza G protein ATP cAMP Fosforylace PKA Desensitizace čichových receptorů

26
Rozpoznávání kvality chemického stimulu
Čich a chuť kódují odlišně Chuť Identifikuje relativně málo látek Používá labeled lines Čich Identifikace extrémního množství látek Používá across fiber pattern Oba systémy principy kombinují

27
Kódování vůní Živočichové rozlišují tisíce vůní Across fiber pattern
Neurony odpovídají na více než jednu látku Jedna látka je vnímána více než jedním neuronem U obratlovců se zdá, že čichové neurony tvoří 4 skupiny a 4 odlišné kanály čichové informace jsou odděleny i v čichovém laloku Identifikace čichových receptorů a genů

28
Čichové receptory 1000 genů u člověka a krysy
Bílkoviny se 7 transmembránovými smyčkami Schopné reagovat s G proteiny Jeden receptor exprimuje jeden gen Vůně aktivuje populaci neuronů schopných interakce Informace je dále vedena jako populační kód Interpretace ve vyšších mozkových centrech

29
Čichový lalok člověka Čichový lalok Mitrální buňky Granulární buňky Chomáčkovité buňky Glomeruly Periglomerulární buňky Receptory

30
Princip čichového kódování – chemotopické uspořádání
Mitrální buňky – projekční neurony Kolik glomerulů tolik receptorových typů Glomeruly Receptorové typy

31
Čichový lalok a glomeruly
Čichové laloky obratlovců, členovců, měkkýšů, červů mají podobnou organizaci –glomeruly (savci glomerulů) Hustě propletená dendritická a axonová zakončení Buněčné typy : Receptory Mitrální buňky – výstup Chomáčové buňky - výstup Periglomerulární buňky Interglomerulární komunikace - inhibiční Granulární buňky Nevstupují do glomerulů - inhibiční
 Hustě propletená dendritická a axonová zakončení. Buněčné typy : Receptory. Mitrální buňky – výstup. Chomáčové buňky - výstup. Periglomerulární buňky. Interglomerulární komunikace - inhibiční. Granulární buňky. Nevstupují do glomerulů - inhibiční.")
33
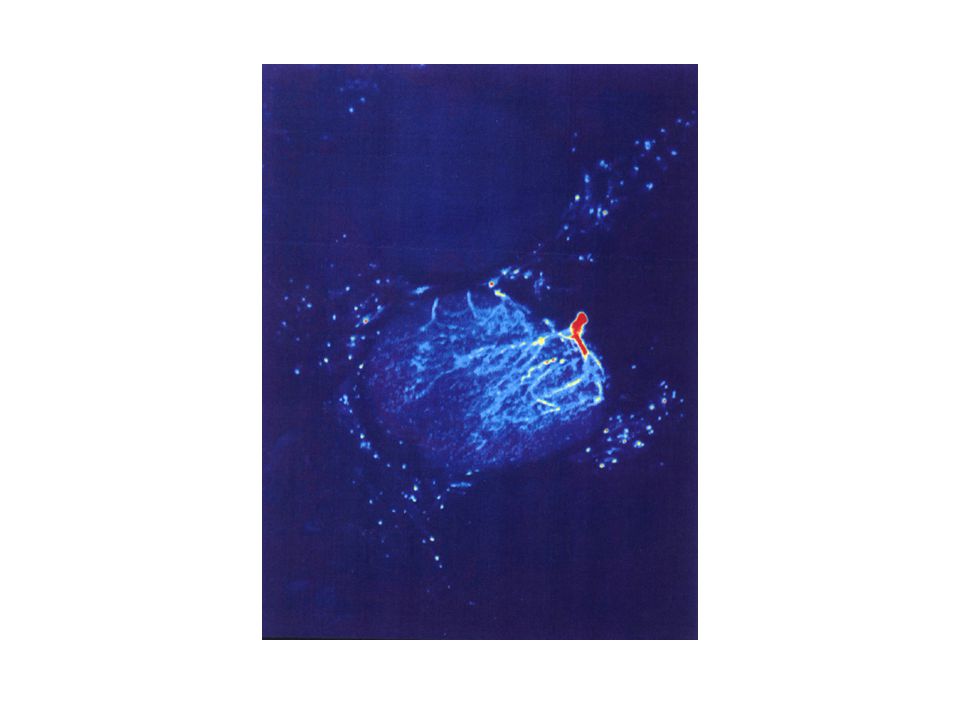
Optické zobrazení glomerulární aktivity v čichovém laloku pomocí Ca sensitivního fluorescenčního barviva Amylacetát Limonene Etyl-n-butyrát

34
Čichový lalok Konvergence – 5-10 tisíc receptorů
Podnět aktivuje specifickou kombinaci glomerulů Z glomerulu vychází cca 35 mitrálních a 80 chomáčových buněk Oscilace projekčních neuronů Svazování signálů, přestavba synaptických dendrodendritických recipročních spojů – čichová paměť? Korové kombinační detektory

35
Dráhy čichového systému člověka
Čichový nerv Čichový lalok Primární čichová kůra (piriformní a entorhinální kůra) Bez přepojení v THALAMU!! Důkaz evolučního stáří smyslu Další projekce do Limbického systému (vůně – emoce) Mediodorsálního jádra thalamu Z thalamu projekce do orbitofrontální kůry (vědomé rozpoznávání vůní)
 Bez přepojení v THALAMU!! Důkaz evolučního stáří smyslu. Další projekce do. Limbického systému (vůně – emoce) Mediodorsálního jádra thalamu. Z thalamu projekce do orbitofrontální kůry (vědomé rozpoznávání vůní)")
36
Čichové mapy 2 čichové mapy – čichový lalok Vyšší centra??
Jak kůra porovnává signály z 1800 glomerulů?

37
Kalózní těleso Thalamus Čichový lalok Orbitofrontální kůra Mozeček Pyriformní kůra Entorhinální kůra

38
Pyriformní kůra Entorhinální kůra Orbitofrontální komplex Frontální lalok Z thalamu Do thalamu Čichový lalok Čichový nerv

39
Čichový neuron obratlovců
Vzduch Dendrit A Póry C Tekutina Vzduch Vzduch Pór Nosní dutina Tělo Tekutina Tekutina Axon Čichový neuron hmyzu Tělo Tělo B Axon Axon Čichový neuron obratlovců Čichový neuron červů

40
Tykadlo švába Pór Dendrity Sensila Kutikula tykadla

41
Čelní pohled na mozek švába
Zrakový lalok Houbovitá tělesa Nerv Tykadlový nerv MGC čichový lalok Glomeruli Jícen

42
Psychofyzika čichu Extrémní citlivost Adaptace Absence modalit
slovníku popisující vůně Klasifikace vůní podle chemických struktur Některé vůně se při správném smíchání vyruší Některé vůně voní jinak při různých koncentracích (indol – psí exkrementy v nízkých koncentracích voní květinově) Schopnost rozpoznávání optických izomerů Neschopnost rozlišení čisté látky a směsi Směsi látek nepříjemných mohou dát příjemnou vůni
 Schopnost rozpoznávání optických izomerů. Neschopnost rozlišení čisté látky a směsi. Směsi látek nepříjemných mohou dát příjemnou vůni.")
43
Bolest, tlak, teplo, chlad, vibrace, šimrání
Somatický smysl Povrch těla Bolest, tlak, teplo, chlad, vibrace, šimrání

44
Somatický smysl - charakteristika
Není soustředěn do specializovaného orgánu Uvnitř a po celém povrchu těla 2 anatomické dráhy 1 mozková oblast Mediuje více modalit Kvalita stimulu (měkký, tvrdý, ostrý, horký, studený…) Lokalizace
 Lokalizace.")
45
Mechanoreceptory savčí kůže
Volná zakončení Merkelův komplex SLOW /* SLOW Receptory chlupových váčků Paciniho tělíska FAST FAST Maissnerova tělíska Ruffiniho tělíska FAST SLOW /* ADAPTACE

46
Smyslové neurony savčí kůže
Merkelův komplex Smyslové neurony savčí kůže Receptor chlupového váčku Volná zakončení Neochlupená kůže Ruffiniho tělísko Paciniho tělíska Potní žláza Meissnerovo tělísko Merkelův komplex Paciniho tělísko Volná zakončení Ochlupená kůže

47
Mechanoreceptory svalu a šlach
Svalové vřeténko Ruffiniho tělísko Svalové vlákno Golgiho šlachové tělísko Paciniho tělísko Šlacha

48
Somatosensorické receptory obratlovců
Různé aferentní dráhy Rozdílná rychlost vedení vzruchu Myelinizované aferenty Ab m/s (6-12 mm) - dotek, chlad, ostrá bolest Ad – m/s (1-5 mm) – teplo, neohraničená bolest, šimrání Nemyelinizované aferenty C – 0,5-2 m/s (0,2-2 mm) Aferentní signály z vnitřních orgánů často podvědomá signalizace Přispívají k vnímání těla v prostoru – kinestéze Vegetativní NS Receptory natažení svalů a šlach
 - dotek, chlad, ostrá bolest. Ad – 5-30 m/s (1-5 mm) – teplo, neohraničená bolest, šimrání. Nemyelinizované aferenty. C – 0,5-2 m/s (0,2-2 mm) Aferentní signály z vnitřních orgánů. často podvědomá signalizace. Přispívají k vnímání těla v prostoru – kinestéze. Vegetativní NS. Receptory natažení svalů a šlach.")
49
Transdukce Specifické iontové kanály – ukotvené
k cytoskeletu buňky K externímu skeletu – uhlovodíkové řetězce Natažením aktivované Otevírají se na začátku stimulu Natažením inhibované Zavírají se na začátku stimulu Externí struktury „tvarují“ odpověď mechanoreceptoru Transdukce na volných nervových zakončeních neznám

50
Vnější lamely Myelinizovaný axon Vnitřní lamely Nemyelinizované nervové zakončení Paciniho tělísko Generátorový potenciál bez obalu Stimulus

51
Diskriminace dotyku Jak rychle a kterým směrem se stimulus pohybuje
Fast adapting receptory (FA) pálí více při vyšší rychlosti než slow adapting (SA) Směrově specifické mechanoreceptory Míra rozlišení – 1,5 mm vzdálené stimuly
 pálí více při vyšší rychlosti než slow adapting (SA) Směrově specifické mechanoreceptory. Míra rozlišení – 1,5 mm vzdálené stimuly.")
52
Rozlišování detailů kožními mechanoreceptory prstu opice
Kůra SA receptory FA receptory Odpovědi receptorů na stimulaci

53
Somatosensorické dráhy
Thalamus Primární somatosensorická kůra Kalózní těleso Střední mozek Střední lemniskus Jádra zadních míšních provazců Prodloužená mícha Somatosensorické dráhy Zadní míšní kořen Mícha

54
Thalamus Primární somatosensorická kůra Kalózní těleso Střední mozek Prodloužená mícha Zadní míšní ganglia Spinothalamický trakt Mícha

55
Korová reprezentace somatosensorické informace
Druhotná somatosensorická kůra v postranním záhybu Primární somatosensorická kůra SI SII Spánkový lalok Mozeček Most Prodloužená mícha

56
Organizace SS kůry SI SII Topografické uspořádání
Vstupy odděleny ve sloupcích SII Hrubé topografické uspořádání Vstupy z thalamu a z SI Abstrakce povrchových vlastností ze složitých podnětů Ne zcela známo

57
Bolest Komplexní vjem s výraznou emocionální složkou
Receptory – nociceptory – info o poškození tkání Volná nervová zakončení bez periferních struktur, mono i polymodální, různé transdukční mechanismy, lze selektivně blokovat Transdukční mechanismus není znám

58
Centrální zpracování – dráhy bolesti
Bolestivé podněty se přepojují V mozkovém kmeni Thalamu Somatosensorické kůře Neví se, jak je bolest v kůřč zpracovávána Na všech centrálních synapsích dochází k modulaci Somatosensorickými vstupy – vrátková teorie bolesti Descendentní kontrolou z mozku Modulace vede ke změnám citlivosti na bolest Fyziologické Patofyziologické změny hyperalgesie

59
Kontrola vnímání bolesti
Somatosensorická Aktivace somatosensorických aferentních vláken s receptivními poli sousedícími s nociceptorem inhibuje nocicepční neurony v thalamu a v kůře – synaptické interakce Centrální sestupné dráhy Opiátová – uvolňuje endogenní opioidní peptidy Distribuce opiátových receptorů v mozku ukazuje naúčast nejen v modulaci bolesti Serotoninová Norepinefrinová

60
Sestupný trakt modulace bolesti Spinothalamický trakt Info z nociceptoru Pre-synaptická modulace Modulační neuron Nociceptor Post-synaptická modulace Tělo neuronu nocicepční dráhy

61
Somatický smysl bezobratlých
Somatosensorické info zpracovávány v gangliích dané úrovně U hmyzu prokázáno topografické uspořádání

62
Vnímání rovnováhy U všech živočichů Vestibulární aparát - obratlovci
Statocysty se statolity Vestibulární aparát - obratlovci Otolitový orgán (párový) utrikulus a sakulus makula – oblast s mechanoreceptory 3 polokruhovité chodby – ortogonálně orientované
 utrikulus a. sakulus. makula – oblast s mechanoreceptory. 3 polokruhovité chodby – ortogonálně orientované.")
63
Vestibulární aparát člověka
Polokruhovité chodby Utrikulus (váček) Sakulus Scala tympani Scala media Hlemýžď Scala vestibuli
 Sakulus. Scala tympani. Scala media. Hlemýžď. Scala vestibuli.")
64
Orgán rovnováhy – utrikulus a sakulus
Mechanoreceptory Vláskovité buňky Anaxonální nespikující neurony Uvolňují mediátor nepřetržitě na aferentní neurony vestibulárních jáder Orgán rovnováhy – utrikulus a sakulus Poloha – horizontálně orientovaná makula utrikulu Pohyb – vertikálně orientovaná makula sakulu

65
Vliv úklonu hlavy na stereocilie utrikulu
Vestibulární nerv Polokruhovité chodby Stereocilie Utrikulus Otolity Makula v klidu Úklon hlavy stranou

66
Polokruhovité chodby Rotace hlavy Vyplněny tekutinou
Detekce vzájemného pohybu tekutiny a sten kanálů Kontrola polohy očí při pohybu hlavy Každá chodba má ztluštěninu – ampula Vláskovité bb zanořeny do gelatinozní kupuly

67
Endolymfa Kupula Zrychlení Odpověď neuronů

68
Odpověď vestibulárního receptoru opice na rotaci celého těla v rovině kanálu
Neměnná rychlost 0 pohyb 0 pohyb Zrychlení Frekvence AP Receptorový potenciál Čas (s)
")
69
Transdukce ve vestibulárních buňkách
Receptory podobné buňkám kochlei Anaxonální Nespikující Stereocilie různé velikosti Ohyb ve směru k nejvyšším - depolarizuje Ve směru k nejnižším – hyperpolarizuje Morfologicky polarizované Mechanické otvírání nebo zavírání nespecifických kationtových kanálů stereocilií

70
Vestibulární buňky ampulí
Řady pravidelně uspořádány endolymfa ohýbá všechny stejným směrem Buňky utrikulu a sakulu Uspořádání do 2 zón Různá orientace stereocilií Citlivost k akceleraci ve všech směrech

71
Polokruhovité chodby Utrikulus Vestibulární nerv Makula Otolit L A Šipky označují směr ohybu stereocilií, který depolarizuje P M

72
Vestibulární dráhy 4 vestibulární jádra v prodloužené míše (superior, lateral, medial, inferior) 2 sestupné trakty Mediální vestibulospinální trakt Laterální vestibulospinální trakt Vzestupné dráhy Mozeček Most nc. abducens – Střední mozek Nc. trochlear Okulomotorické jádro Kůra

73
Okulomotorické jádro Thalamus Superior colliculus Trochleární jádro Inferior colliculus Vestibulární jádra Do mozečku Superior Lateral Nc. abducens Medial Inferior Postranní vestibulospinální trakt Střední vestibulospinální trakt Vestibulární dráhy

74
Elektrický smysl

75
Elektrický smysl Vnímání elektrického pole
Pasivní (nekontrolují svoje) Aktivní (elektrogenní) Obrana před predátory Elektrolokace (navigace nebo orientace) Evolučně odvozen z postranní čáry Ampulární orgány Pod kůží - ampula spojená s povrchem Tuberální orgány Pod kůží – nejsou spojeny s povrchem Reagují na fluktuace napětí
 Aktivní (elektrogenní) Obrana před predátory. Elektrolokace (navigace nebo orientace) Evolučně odvozen z postranní čáry. Ampulární orgány. Pod kůží - ampula spojená s povrchem. Tuberální orgány. Pod kůží – nejsou spojeny s povrchem. Reagují na fluktuace napětí.")
76
Nevodivý objekt Princip elektrolokace Vodivý objekt

77
Ampulární orgán Kůže Elektroreceptory Smyslový nerv Tuberální orgán kůže Elektroreceptory Smyslový nerv

78
Extrémní citlivost receptorů
Aferenty: Prodloužená mícha Tektum Mozeček Kůra Topografické uspořádání podle umístění receptoru na těle Extrémní citlivost receptorů Elektrický gradient 0,005 mV/cm proud generovaný 1,5 V baterií v oceánu s póly 3000 km od sebe!!! Navigace v magnetickém poli země Pohyb napříč pole vyvolává větší elektrický gradient než pohyb podél Vyhledávání kořisti Svaly kořisti produkují el. potenciál

79
Elektrické pole vyvolává
Depolarizaci způsobenou vtokem Ca2++ Receptor se repolarizuje následným výtok K+ Ampulární orgány Receptory anaxonální, nespikují Tonická odpověď Kontinuální uvolňování mediátoru Tuberální orgány Receptory anaxonální, SPIKUJÍ, jiné? obratlovčí chuťové Fázická odpověď (nekódují absolutní intenzitu) Některé přenášejí svou excitaci na neurony vyššího řádu elektricky
 Některé přenášejí svou excitaci na neurony vyššího řádu elektricky.")
80
Magnetický smysl Vnímání přítomnosti a orientace magnetického pole zěmě Bakterie magnetity – směsný oxid železa Magnetity i u jiných druhů, mechanismus neznám Demagnetizace bez vlivu na chování (včely) Holub – magnety po stranách hlavy dezorientují Zrakový systém – speciální fotoreceptory, které využívají Elektron Spin Resonance
 Holub – magnety po stranách hlavy dezorientují. Zrakový systém – speciální fotoreceptory, které využívají Elektron Spin Resonance.")
81
Uspořádání magnetitů v bakterii
Magnetity

82
Elektron Spin Resonance
Odpověď fotoreceptorů modulována orientací v magnetickém poli Důsledkem je rozdílné vidění barev Behaviorální evidence Fyziologické evidence chybí Pravý magnetický smysl Žraloci – detekce změn elektrického pole při pohybu magnetickým polem Jiné smysly Vlhkost (hmyz) Infračervené vlnění (hadi - zrak)
 Infračervené vlnění (hadi - zrak)")
Podobné prezentace