Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Obecný metabolismus Metabolismus: Základní pojetí a obsah pojmu.
Cesty přenosu signálu: Úvod do metabolismu informace. Regulace metabolických drah: Glykolýza a glukoneogeneze. Glykogen. Cyklus trikarboxylových kyselin. Oxidatívní fosforylace. Calvinův cyklus a metabolismus fosfátů pentos. Metabolismus mastných kyselin Obrat aminokyselin a jejich biosyntéza a katabolismus.

2
METABOLISMUS Řešení dvou základních otázek
1. Jak buňky získávají energii a redukční sílu ze svého okolí ? 2. Jak buňky syntetizují základní stavební kameny makromolekul a poté samotné makromolekuly ? Oba procesy probíhají jako vysoce organizované chemické reakce - metabolismus.

3
Buňky využívají různé typy energií
- pro mechanický pohyb - k aktivnímu transportu molekul a iontů - k syntéze makromolekul z jednoduchých prekurzorů Volná energie využívané při těchto procesech se získává z vnějšího prostředí První zákon termodynamiky

4
Fotosyntetyzující organismy – energetický základ života na Zemi
Fototrofní organismy – přeměna světelné energie na energii chemických sloučenin. Chemotrofní organismy – získávají chemickou energii oxidací chemických sloučenin vytvořených fototrofy. Metabolismus nebo intermediární metabolismus – v buňkách probíhá v každém okamžiku tisíce transformací energie. Energie je uvolňována z energeticky bohatých sloučenin a využita k biosyntetickým procesům.

5
Metabolismus glukosy - glykolýza

6
Metabolické dráhy

7
Spřažené reakce Termodynamicky nevýhodná reakce je poháněna exergonní reakcí. Obvykle přenos fosfátu z ATP. Kapitola 14 – pohyblivé schéma.

8
Hydrolýza ATP posunuje rovnováhu spřažených reakcí
Termodynamicky nevýhodná reakční sekvence může být převedena na výhodnou spojením s potřebným množstvím molekul ATP v nové reakci (fosforylace substrátů).
.")
9
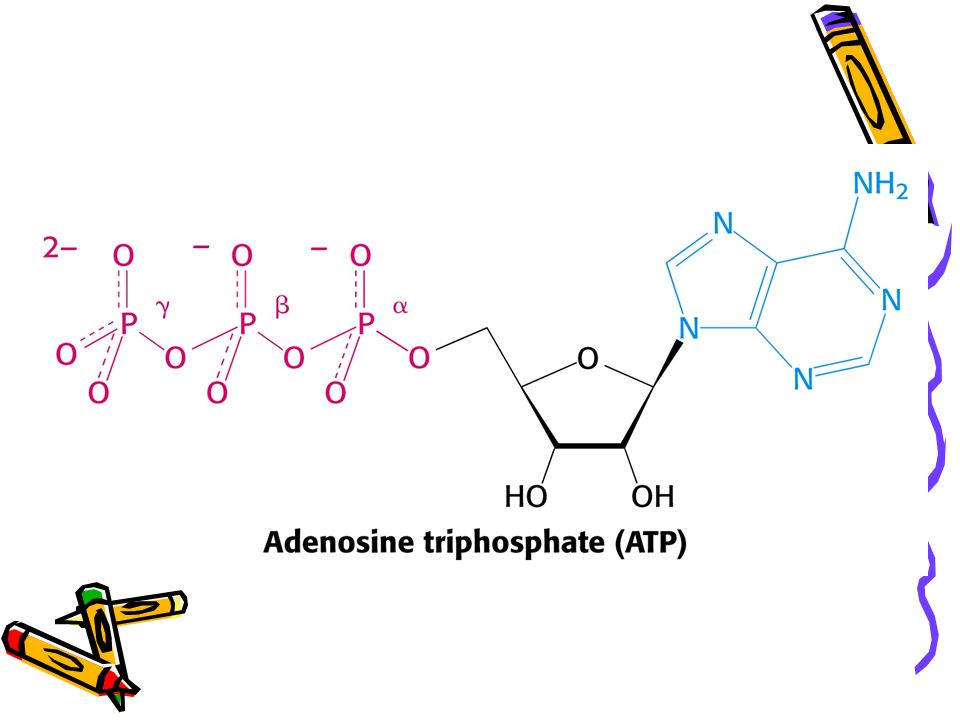
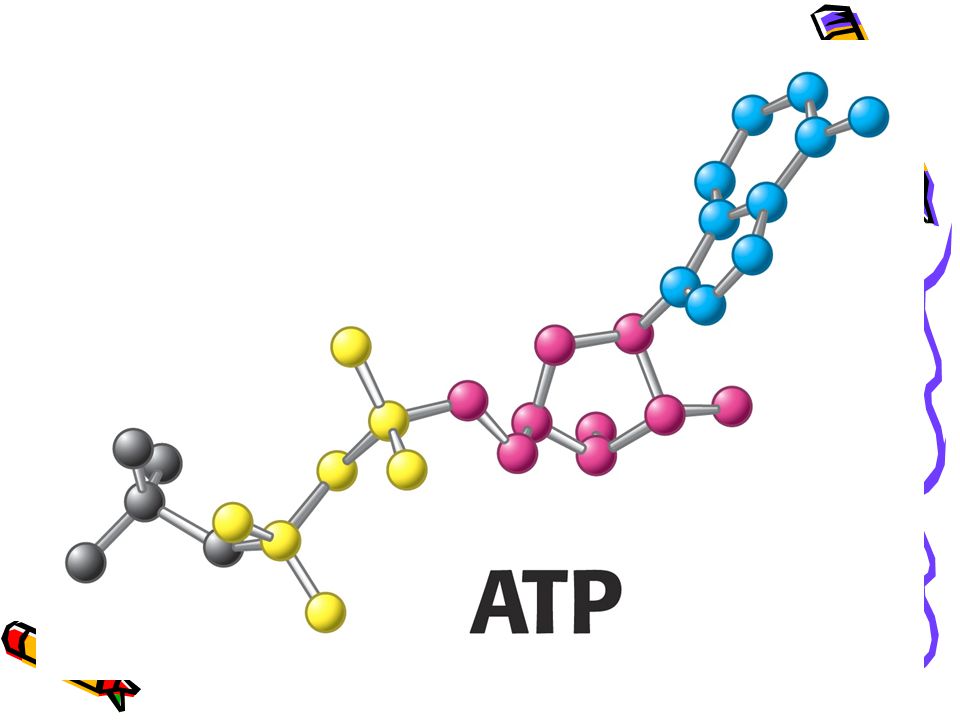
Strukturní základy vysokého potenciálu přenosu fosfátu ATP
Glycerol-3-fosfát + voda = glycerol + fosfát - 9, 2 kJ.mol ATP + voda = ADP + fosfát + H - 30, 5 kJ.mol Rozdíly: rezonanční stabilita; elektrostatické odpuzování; stabilizace hydratací.

10
Celková volná energie chemické reakce je součtem volných energií dílčích reakcí

11
Možné typy hydrolýzy ATP

12
Termodynamicky nevýhodná reakce (25oC)
")
13
Termodynamicky nevýhodná reakce ve spojení s hydrolýzou ATP: A + ATP + H2O = B + ADP + Pi + H+ D Go´= - 13, 8 kJ.mol-1 Rovnovážná konstanta spřažené reakce:

14
Obecně nATP mění rovnováhu reakce násobkem 108n.
Potenciál ATP se může uplatnit např. při přenosu fosfátu na protein (enzym), který je takto aktivován nebo při aktivním transportu látek přes membránu jako je např. symport Na+ a K+ při kterém je fosforylcí aktivována Na+ - K+ pumpa.
, který je takto aktivován nebo při aktivním transportu látek přes membránu jako je např. symport Na+ a K+ při kterém je fosforylcí aktivována Na+ - K+ pumpa.")
18
Glycerol-3-fosfát + H2O = Glycerol + Pi D Go´= - 9, 2 kJ.mol-1

19
Jaký je strukturní základ vysokého potenciálu přenosu fosfátu ATP ?
Změna standardní volné energie závisí na rozdílu volných energií produktů a reaktantů, tedy na jejich struktuře. Zkoumáme ADP a Pi jako produkty hydrolýzy ATP a ATP jako reaktant. Tři podstatné faktory : - rezonanční stabilita - ektrostatické odpuzování - stabilizace hydratací ADP a Pi mají větší rezonannční stabilitu než ATP. Orthofosfát má řadu resonančních struktur, kdežto g-fosfátová skupina ATP menší počet. Při pH 7 má trifosfát čtyři záporné náboje, které se odpuzují. Po hydrolýze ATP repulze mizí! Voda se váže mnohem efektivněji na Pi a ADP než na fosfoanhydridové vazby ATP. Stabilizace ADP a Pi hydratací !!

20
Resonanční struktury orthofosfátu

21
Nepravděpodobná resonanční struktura – dva atomy P s pozitivním nábojem vedle sebe.

22
Sloučeniny s vysokým potenciálem přenosu fosfátu – mohou fosforylovat ADP za tvorby ATP. ATP je meziprodukt mezi těmito vysokoenergetickými sloučeninami a sloučeninami s nižším potenciálem.

23
Změna standardní volné energie hydrolýzy vybraných fosfátových sloučenin.

24
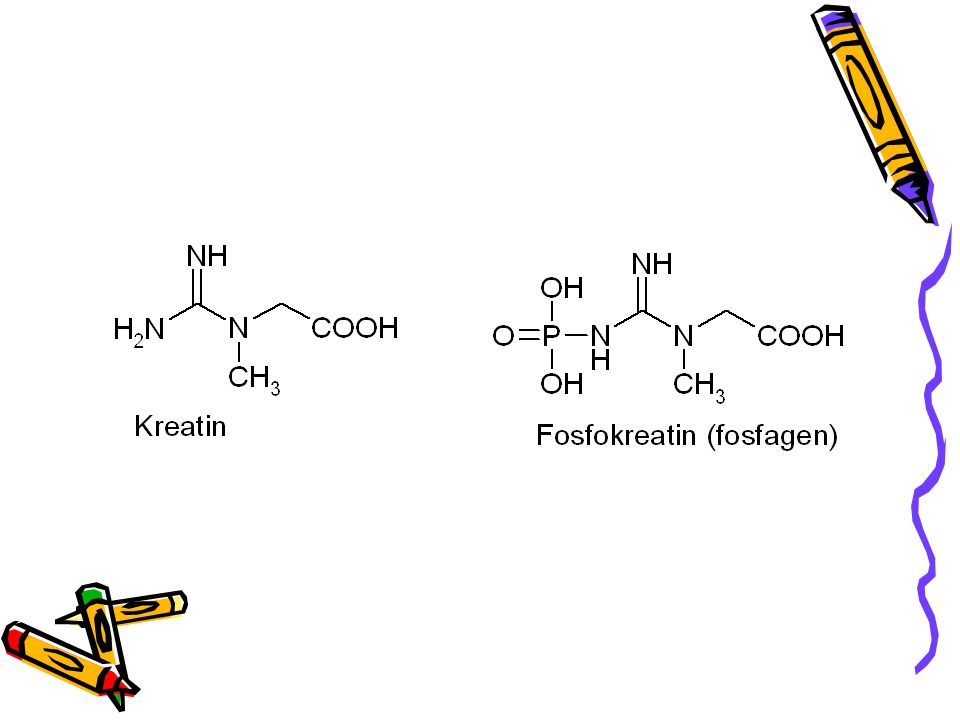
Kreatinfosfát – zásoba energie svalů obratlovců.
Kreatinfosfát slouží u obratlovců jako zásoba svalové energie sloužící k regeneraci ATP z ADP za katalýzy kreatinkinasou: Kreatinfosfát + ADP + H+ = ATP + kreatin Při pH = 7 je D Go´ = - 43, 1 kJ/mol (hydrolýza kreatinfosfátu). Ve srovnání s hydrolýzou ATP (-30,5 kJ/mol) je celková změna rovna – 12,6kJ/mol což odpovídá rovnovážné konstantě = 162. Keq = [ATP] [kreatin] / [ADP] [kreatinfosfát] = 10 D G(stand) / (2,303 RT) = 103/1,36 = 162. Koncentrace v odpočívajícím svalu: [ATP] = 4 mM, [ADP] = 0, 013 mM, [kreatinfosfát] = 25 mM a [kreatin] = 13 mM.
. Ve srovnání s hydrolýzou ATP (-30,5 kJ/mol) je celková změna rovna – 12,6kJ/mol což odpovídá rovnovážné konstantě = 162. Keq = [ATP] [kreatin] / [ADP] [kreatinfosfát] = 10 D G(stand) / (2,303 RT) = 103/1,36 = 162. Koncentrace v odpočívajícím svalu: [ATP] = 4 mM, [ADP] = 0, 013 mM, [kreatinfosfát] = 25 mM a [kreatin] = 13 mM.")
25
Zdroje ATP během tělesného cvičení.

26
Oxidace uhlíkatých sloučenin je důležitým zdrojem
buněčné energie. V typické buňce se ATP spotřebuje do půl minuty po tvorbě. Celkové množství ATP v těle je limitováno 100 g, jeho obrat je vysoký. Odpočívající člověk spotřebuje asi 40 kg ATP/den. Při intenzivní práce to může být až 0, 5 kg/min. Při 2 hod běhu se spotřebuje 60 kg ATP. Tvorba ATP je primární rolí katabolismu. U aerobních organismů je elektronovým akceptorem při oxidaci kyslík a produktem CO2.

27
ATP – ADP cyklus

28
Volná energie oxidace jednoduchých uhlíkatých sloučenin.

29
Mastné kyseliny a sacharidy jsou největšími zdroji energie. Proč ?

30
Fosfáty s vysokým potenciálem spojují oxidaci uhlíku s tvorbou ATP
Fosfáty s vysokým potenciálem spojují oxidaci uhlíku s tvorbou ATP. Jako příklad – glyceraldehyd-3-fosfát.

31
Tvorba ATP na úrovni substrátu !!!!

32
Protonové gradienty.

33
Tři stádia katabolismu.
1. Velké molekuly se štěpí na menší jednotky. 2. Množství malých molekul se odbourává na několik jednoduchých, které hrají centrální roli v metabolismu. Např. acetyl CoA. 3. Kompletní oxidací acetyl CoA se vytváří ATP, CO2 a voda.

34
Tři stupně katabolismu.

35
V metabolických drahách je mnoho se opakujících motivů.
1. Aktivovaní přenašeči elektronů pro oxidaci energeticky bohatých sloučenin (substrátů) 2. Aktivovaní přenašeči elektronů k reduktivním biosyntézám. 3. Aktivovaní přenašeči dvojuhlíkatých štěpů 4. Další různí přenašeči (tabulka) Kinetická stabilita těchto molekul umožňuje bez přítomnosti katalyzátorů jejich biologické funkce a enzymovou kontrolu toku volné energie a redukční síly
 2. Aktivovaní přenašeči elektronů k reduktivním biosyntézám. 3. Aktivovaní přenašeči dvojuhlíkatých štěpů. 4. Další různí přenašeči (tabulka) Kinetická stabilita těchto molekul umožňuje bez přítomnosti katalyzátorů jejich biologické funkce a enzymovou kontrolu toku volné energie a redukční síly.")
36
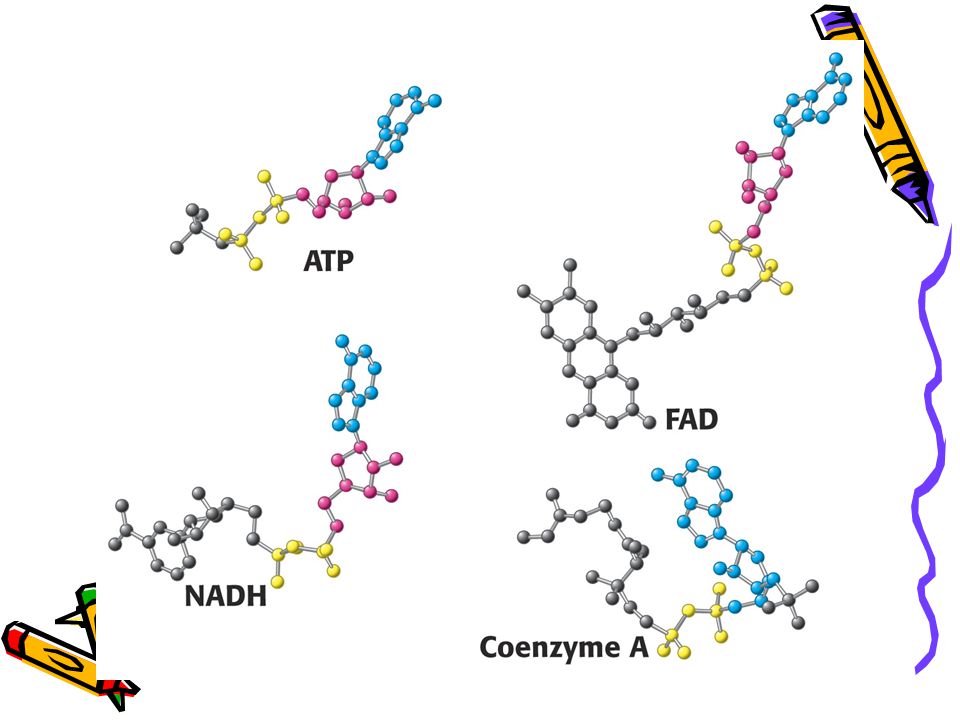
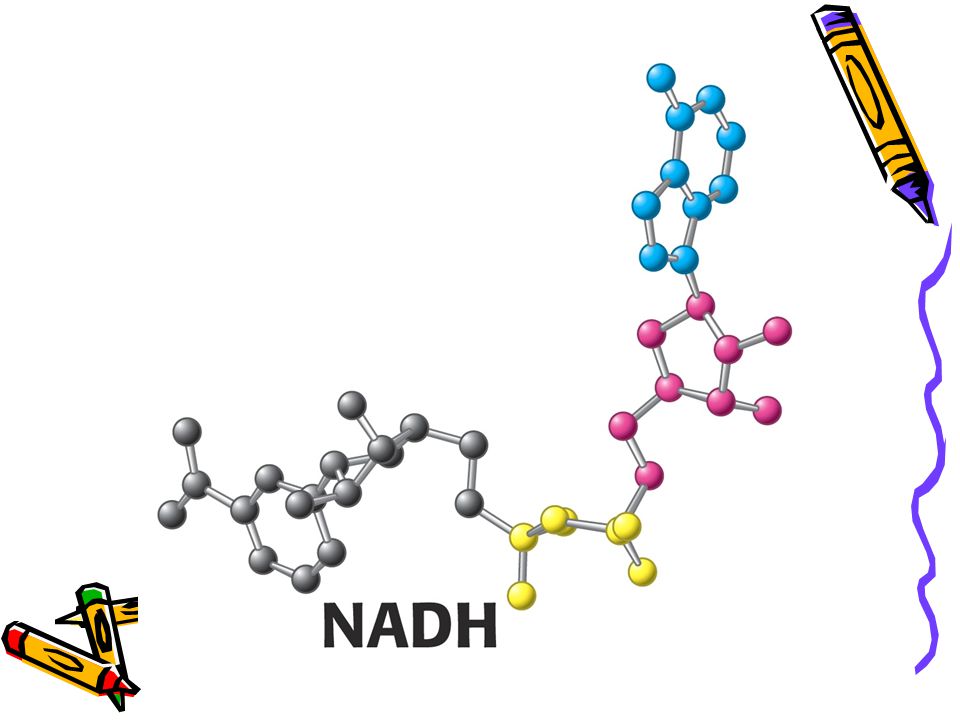
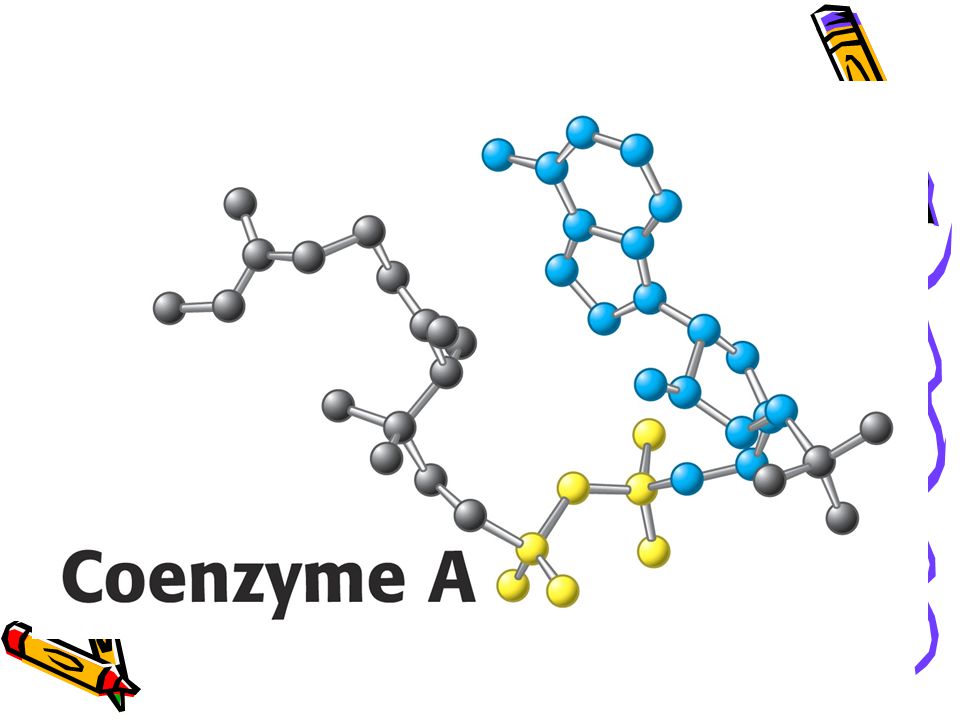
Vybraní aktivovaní přenašeči v metabolismu a jejich prekurzory

39
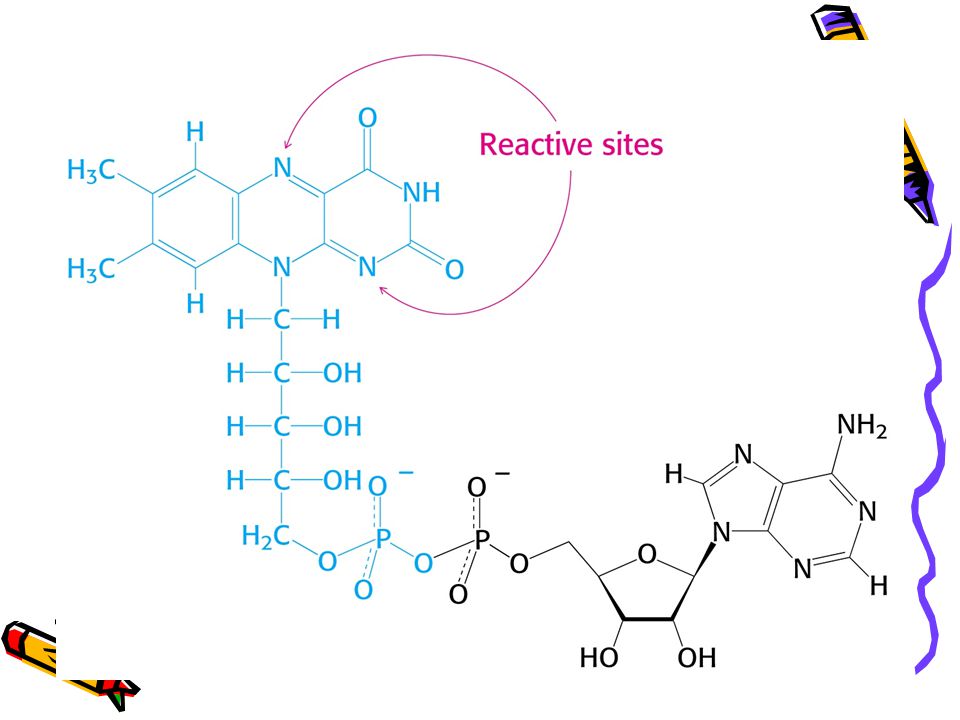
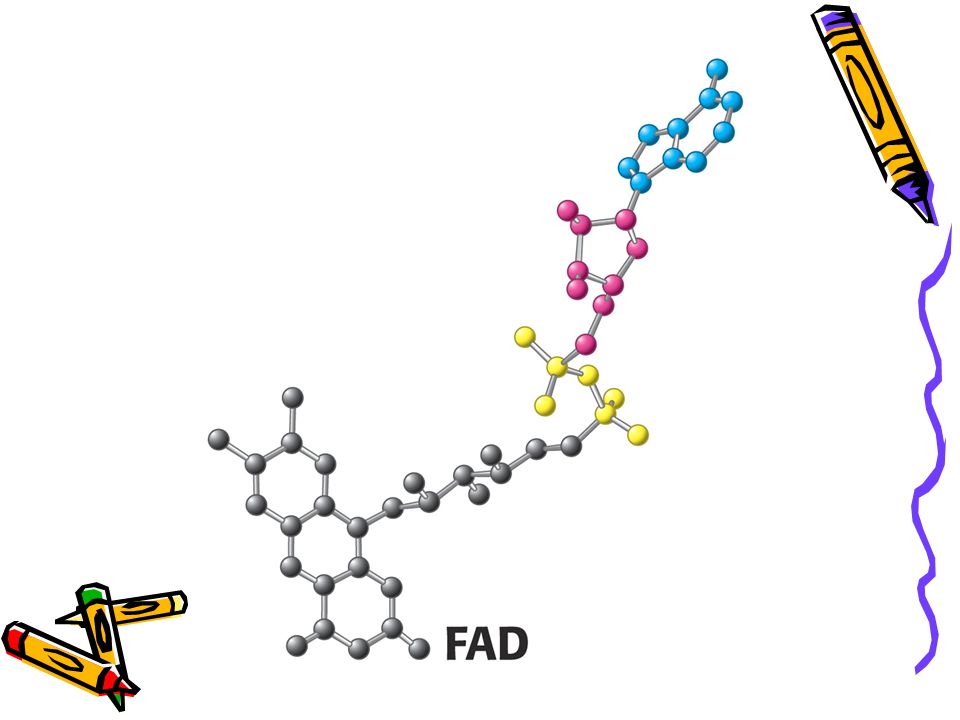
Struktury reaktivních částí FAD a FADH2

40
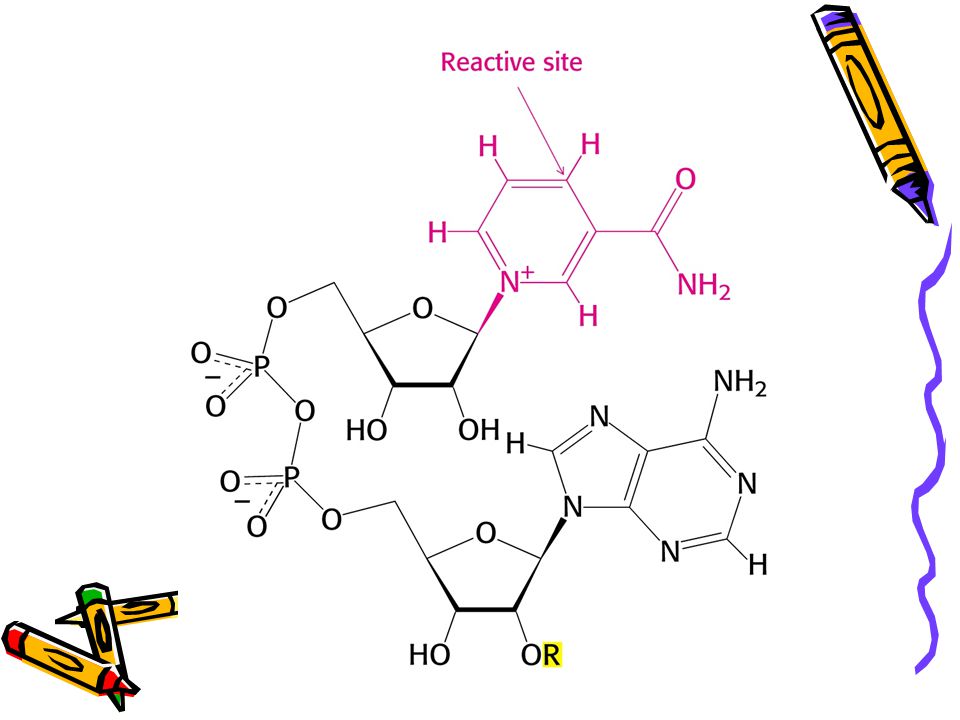
Donorem elektronů při mnoha biosyntézách je NADPH. Např
Donorem elektronů při mnoha biosyntézách je NADPH. Např. biosyntéza mastných kyselin

41
Aktivovaný nosič dvojuhlíkatých štěpů a acylů CoASH
Aktivovaný nosič dvojuhlíkatých štěpů a acylů CoASH. Acetyl CoA + H2O = acetát + CoA + H+. D Go´ = - 31, 4 kJ/mol.

42
Proč je thioester termodynamicky výhodnější než kyslíkatý ester ?

43
Typy chemických reakcí v metabolismu
Typ reakce Popis Oxidačně-redukční Přenos elektronů Tvorba vazeb za účasti ATP Tvorba kovalentních vazeb Izomerizace Přemístění atomů, tvorba izomerů Přenos skupin Přenos funkčních skupin z jedné molekuly na druhou Hydrolýza Štěpení vazby adicí vody Adice a odstranění funkčních sk. Adice na dvojnou vazbu a odštěpení funkční skupiny za tvorby dvojné vazby

44
Oxidačně-redukční reakce. Obě reakce jsou součástí citrátového cyklu

45
Karboxylace – tvorba vazby C – C za účasti ATP. Pyruvátkarboxylasa.

46
Izomerace

47
Přenos skupin – přenos fosfátu.

48
Hydrolýza – peptidasy, proteasy.

49
Adice funkční skupiny na dvojnou vazbu nebo odštěpení funkční skupiny za tvorby dvojné vazby (lyasy)
")
50
Dehydratace

51
Řada metabolických drah má podobné sekvence reakcí
Řada metabolických drah má podobné sekvence reakcí. (ACP = acyl carrier protein)
")
52
Regulace metabolických procesů.
1. Hladinou (množstvím) enzymů. 2. Katalytickou aktivitou enzymů. 3. Dostupností substrátů. 1. Rychlost syntézy a odbourávání, změna rychlosti transkripce. 2. Aktivita enzymů: reversibilní allosterická kontrola, inhibice produktem, kovalentní modifikace, hormony. 3. Kontrola vstupu substrátů do kompartmentu buňky. Obecným principem regulace je kompartmentizace – oddělení dvou protichůdných dějů membránou. Řada reakcí je regulována energetickým nábojem v buňce. ATP generované katabolismem je inhibováno energetickým nábojem, ATP spotřebovávané anabolismem je stimulováno energetickým nábojem.
 enzymů. 2. Katalytickou aktivitou enzymů. 3. Dostupností substrátů. 1. Rychlost syntézy a odbourávání, změna rychlosti transkripce. 2. Aktivita enzymů: reversibilní allosterická kontrola, inhibice produktem, kovalentní modifikace, hormony. 3. Kontrola vstupu substrátů do kompartmentu buňky. Obecným principem regulace je kompartmentizace – oddělení dvou protichůdných dějů membránou. Řada reakcí je regulována energetickým nábojem v buňce. ATP generované katabolismem je inhibováno energetickým nábojem, ATP spotřebovávané anabolismem je stimulováno energetickým nábojem.")
53
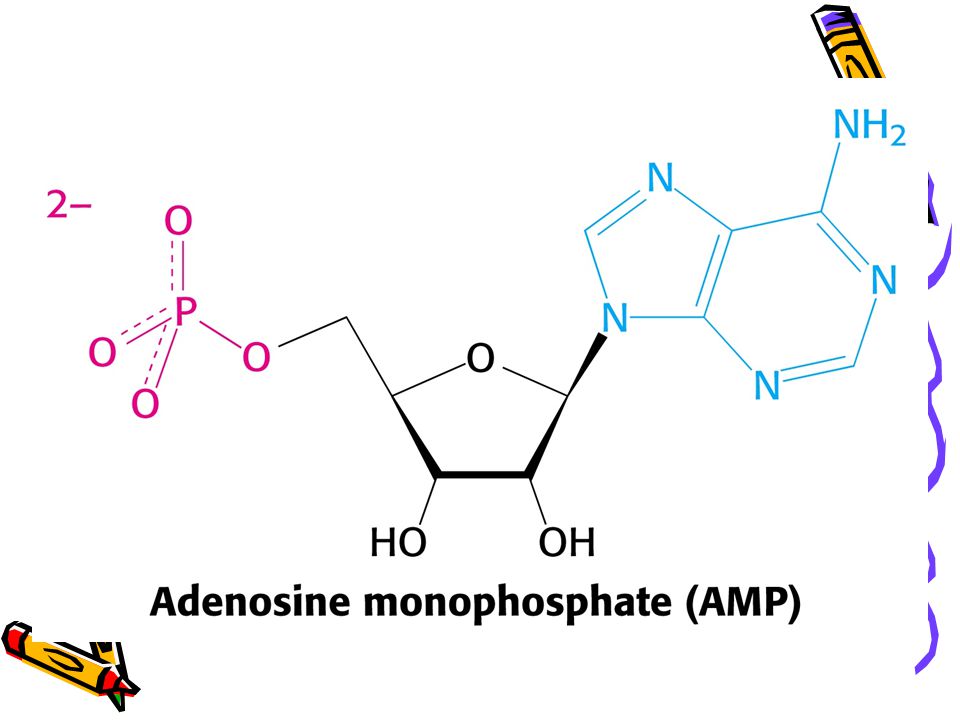
Energetický náboj buňky
Energetický náboj = [ATP] + ½ [ADP] / [ATP] + [ADP] + [AMP]. ATP obsahuje dvě anhydridové vazby, ADP jednu. Energetický náboj buňky je v rozmezí 0 až 1. (U většiny buněk od 0, 80 do 0, 95). Vnitrobuněčná koncentrace ATP se udržuje v rozmezí 2 – 10 mM. Koncentrace ADP a Pi jsou variabilní. Při typické koncentraci [ATP] = 3, 0 mM, konc. [ADP] = 0, 8 mM konc.[ Pi] = 4 mM je volná energie hydrolýzy ATP při 37oC:
. Vnitrobuněčná koncentrace ATP. se udržuje v rozmezí 2 – 10 mM. Koncentrace ADP a Pi jsou variabilní. Při typické koncentraci. [ATP] = 3, 0 mM, konc. [ADP] = 0, 8 mM. konc.[ Pi] = 4 mM je volná energie hydrolýzy ATP při 37oC:")
54
Volná energie hydrolýzy ATP při 37o C (pokračování):
DG = D G o´ + RT ln [ADP] .[ Pi ]/ [ATP] = - 30, 5 kJ.mol-1 + (8, 3145 J. K-1.mol-1)(310 K) ln[( 0, 8 x 10-3 M). (4, 0 x 10-3 M) / 3, 0 x 10-3M)] = (-30, 5 kJ.mol - 17, 6 kJ.mol) = - 48, 1 kJ.mol-1
(310 K) ln[( 0, 8 x 10-3 M). (4, 0 x 10-3 M) / 3, 0 x 10-3M)] = (-30, 5 kJ.mol - 17, 6 kJ.mol) = - 48, 1 kJ.mol-1.")
55
Regulace metabolismu energetickým nábojem
Regulace metabolismu energetickým nábojem. Vysoká koncentrace ATP inhibuje rychlost katabolických dějů při kterých se ATP vytváří a stimuluje děje anabolické.

56
Evoluce metabolických drah.
Aktivovaní přenašeči jako ATP, NADH, FADH2 a koenzym A obsahují adenosin difosfátovou jednotku. Pravděpodobně se odvozují od prvtního RNA katalyzátoru – robozymu.

68
Úloha kreatinfosfátu ve svalech
Kreatinfosfát(KP) + ADP kreatinkinasa ATP + kreatin (K) Při pH 7 je stand. volná energie hydrolýzy KP – 43, 1 kJ/mol. Pro tvorbu ATP z ADP a KP je sve -12, 6 kJ/mol. Což odpovídá rovnovážné konstantě = 162. Typické koncentrace v odpočívajícím svalu: [ATP] = 4 mM; [ADP]= 0, 013 mM; [KP] = 25 mM a [K] = 13 mM. KP je jediný zdroj regenerace ATP při prvních 4 sek. Běhu na 100 m.
 + ADP kreatinkinasa ATP + kreatin (K) Při pH 7 je stand. volná energie hydrolýzy KP – 43, 1 kJ/mol. Pro tvorbu ATP z ADP a KP je sve. -12, 6 kJ/mol. Což odpovídá rovnovážné konstantě = 162. Typické koncentrace v odpočívajícím svalu: [ATP] = 4 mM; [ADP]= 0, 013 mM; [KP] = 25 mM a [K] = 13 mM. KP je jediný zdroj regenerace ATP při prvních 4 sek. Běhu na 100 m.")
Podobné prezentace