Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Společenstva a ekosystémy

2
SPOLEČENSTVO - úvod Vlastnosti společenstev souhrnné, kolektivní, vyplývající z vlastností populací a organismů společenstvo tvořících – patří sem charakteristiky jako diversita, biomasa společenstva, produktivita společenstva. ale protože společenstvo není jen sumou organismů, ale navíc zahrnuje i jejich interakce, můžeme hodnotit i vlastnosti, které vykazuje společenstvo jako entita, tzv. vlastnosti emergentní (vynoří se, až když studujeme celé společenstvo, z vlastností prvků je nevyvodíme). Jako když se peče koláč – těžko uhodneme, jak nám bude chutnat (jak je upečený, křupavý, voňavý) z toho, jak vypadá mouka a vejce. Ve společenstvu sem můžeme zařadit namátkou vlastnosti jako třeba hranice podobnosti kompetujících druhů, stabilita potravních řetězců při disturbanci atp. Primárním cílem ekologie společenstev je odhalování obecných vzorů a zákonitostí v kolektivních i emergentních vlastnostech, takových, která by univerzálně platila, byť by se jednotlivá společenstva ve své druhové skladbě všemožně lišila.
. Jako když se peče koláč – těžko uhodneme, jak nám bude chutnat (jak je upečený, křupavý, voňavý) z toho, jak vypadá mouka a vejce. Ve společenstvu sem můžeme zařadit namátkou vlastnosti jako třeba hranice podobnosti kompetujících druhů, stabilita potravních řetězců při disturbanci atp. Primárním cílem ekologie společenstev je odhalování obecných vzorů a zákonitostí v kolektivních i emergentních vlastnostech, takových, která by univerzálně platila, byť by se jednotlivá společenstva ve své druhové skladbě všemožně lišila.")
3
SPOLEČENSTVO A EKOSYSTÉM
Byť se tradičně podle definice oddělují (ekosystém = společenstvo + abiotické složky), v praxi se tyto kategorie dají těžko studovat odděleně – téměř vždy do svých úvah a studií (ať už jsou na úrovni organismu, populace nebo společenstva) nějakým způsobem zahrnujeme abiotické prostředí, ve kterém organismy žijí. Existují sice jevy, které se realizují až v rámci celého ekosystému (koloběh živin, energie), ale i ty nás velmi často zajímají právě z hlediska vlastností společenstev. Co považovat za SPOLEČENSTVO? Společenstvo může být definováno na jakékoliv úrovni hierarchie, v nejrůznějším velikostním měřítku. Jako společenstvo můžeme chápat celý biom – třeba listnatý opadavý les mírného pásu Palearktu, ale můžeme tak chápat klidně též pouze šípákovou doubravu Českého krasu, nebo (v jejím rámci) jen bezobratlé žijící pod kůrou spadlého dubu, nebo mikroflóru a mikrofaunu v žaludku srnce, co tam běhá okolo. Častokrát se z praktických důvodů při studiu určitého území označuje za společenstvo jen určitá taxonomická skupina (těžko bychom sestudovali úplně všechny druhy organismů na určité ploše, od bakterií a plísní přes kvasinky, řasy, sinice, a prvoky, všechny rostliny, bezobratlé – půdní nematoda, vířníky, roztoče, všechen hmyz, apod.)
, v praxi se tyto kategorie dají těžko studovat odděleně – téměř vždy do svých úvah a studií (ať už jsou na úrovni organismu, populace nebo společenstva) nějakým způsobem zahrnujeme abiotické prostředí, ve kterém organismy žijí. Existují sice jevy, které se realizují až v rámci celého ekosystému (koloběh živin, energie), ale i ty nás velmi často zajímají právě z hlediska vlastností společenstev. Co považovat za SPOLEČENSTVO Společenstvo může být definováno na jakékoliv úrovni hierarchie, v nejrůznějším velikostním měřítku. Jako společenstvo můžeme chápat celý biom – třeba listnatý opadavý les mírného pásu Palearktu, ale můžeme tak chápat klidně též pouze šípákovou doubravu Českého krasu, nebo (v jejím rámci) jen bezobratlé žijící pod kůrou spadlého dubu, nebo mikroflóru a mikrofaunu v žaludku srnce, co tam běhá okolo. Častokrát se z praktických důvodů při studiu určitého území označuje za společenstvo jen určitá taxonomická skupina (těžko bychom sestudovali úplně všechny druhy organismů na určité ploše, od bakterií a plísní přes kvasinky, řasy, sinice, a prvoky, všechny rostliny, bezobratlé – půdní nematoda, vířníky, roztoče, všechen hmyz, apod.)")
4
Charakteristiky společenstev
Jedním z nejjednodušších parametrů je prostý počet druhů. V praxi je to ovšem není tak snadné (viz předchozí strana). A i pokud se omezíme na určitou taxonomickou skupinu, záleží ještě na počtu a velikosti vzorků (málokdy můžeme spočítat celou plochu/objem). obrázek 17.2
. A i pokud se omezíme na určitou taxonomickou skupinu, záleží ještě na počtu a velikosti vzorků (málokdy můžeme spočítat celou plochu/objem). obrázek")
5
Charakteristiky společenstev – indexy diverzity apod.
Indexy diversity Diversita – pestrost zahrnuje kromě počtu druhů i informaci, jak jsou tyto druhu relativně početné. Pokud v rámci taxonomické skupiny, stačí počty, pokud přes mnoho skupin, tak lepší přes biomasy. A ještě máme známý problém s modulárními organismy (co je jedinec?). Diversita roste s počtem druhů a s jejich rovnoměrným rozložením. Ekvitabilita – vyrovnanost je mírou rozložení počtů (biomasy) (0 – 1) Indexy Simpsonův Shannonův k nim ekvitabilita
. Diversita roste s počtem druhů a s jejich rovnoměrným rozložením. Ekvitabilita – vyrovnanost je mírou rozložení počtů (biomasy) (0 – 1) Indexy Simpsonův Shannonův. k nim. ekvitabilita.")
6
Charakteristiky společenstev
Ekvitabilita a její efekt na diverzitu

7
Rank-abundance křivky pro dva typy lesa
![]()
8
Rank-abundance křivky pro chrostíky
![]()
9
Rank-abundance křivky pro korálové ryby
![]()
10
Hnojení a diverzita rostlin

11
Pořadí druhů od nejpočetnějšího k nejvzácnějšímu
Rank-abundance – Rozložení početností jednotlivých druhů v rámci společenstva Nejvyšší diverzita je při vyrovnaném počtu jedinců všech druhů – je to ale přirozený stav? (?? má být lišek stejně jako zajíců, štik jako plotic, krahujců jako koňader ??) Broken-stick effect: Pár druhů dominuje, většina je zastoupena středně, pár druhů vzácných. Řeší to (trochu) počítání přes biomasy. Pořadí druhů od nejpočetnějšího k nejvzácnějšímu (abundance rank)
 Broken-stick effect: Pár druhů dominuje, většina je zastoupena středně, pár druhů vzácných. Řeší to (trochu) počítání přes biomasy. Pořadí druhů od nejpočetnějšího k nejvzácnějšímu. (abundance rank)")
12
Charekteristiky společenstev – indexy diverzity apod.
Diverzita dále hierarchicky rozpracována (Whittaker, 1972) na LOKÁLNÍ – diverzitu – na malé ploše na víceméně uniformním stanovišti a REGIONÁLNÍ – diverzitu – unikátní druhy všech stanovišť v regionu Čili by platilo, že pokud jsou všechny druhy přítomny ve všech habitatech regionu, tak -diverzita = diverzita. Pokud má naopak každé stanoviště unikátní (jinde se neopakující) soubor druhů, tak regionální () diverzita je rovna násobku průměrné diverzity krát počet habitatů. A tento násobek je označován jako diverzita. Je to snad např. k tomu, aby se výpočtem stanovil počet unikátních stanovišť, tedy diverzita = diverzita/ diverzita Z čehož vyplývá, že počet druhů/ diverzita se jistojistě mění s velikostí zkoumaného území. O tom bude zvláštní kapitolka v blízké budoucnosti.
 na. LOKÁLNÍ – diverzitu – na malé ploše na víceméně uniformním stanovišti. a. REGIONÁLNÍ – diverzitu – unikátní druhy všech stanovišť v regionu. Čili by platilo, že pokud jsou všechny druhy přítomny ve všech habitatech regionu, tak -diverzita = diverzita. Pokud má naopak každé stanoviště unikátní (jinde se neopakující) soubor druhů, tak regionální () diverzita je rovna násobku průměrné diverzity krát počet habitatů. A tento násobek je označován jako diverzita. Je to snad např. k tomu, aby se výpočtem stanovil počet unikátních stanovišť, tedy. diverzita = diverzita/ diverzita. Z čehož vyplývá, že počet druhů/ diverzita se jistojistě mění s velikostí zkoumaného území. O tom bude zvláštní kapitolka v blízké budoucnosti.")
13
Podoba společenstev v prostoru
Co ovlivňuje druhovou skladbu společenstva? Gradientová analýza (kritika – výběr gradientu je skoro vždy subjektivní)
")
15
Podoba společenstev v prostoru
Ordinace a klasifikace Ordinace – hledání (pomocí metod multidimensionální statistiky) faktorů/gradientů, jež vysvětlují podobnost/rozdílnost jednotlivých společenstev. Umožňuje predikce a formulování dalších hypotéz. Klasifikace – považuje společenstva za oddělené entity a třídí je do skupin podle shody znaků.
 faktorů/gradientů, jež vysvětlují podobnost/rozdílnost jednotlivých společenstev. Umožňuje predikce a formulování dalších hypotéz. Klasifikace – považuje společenstva za oddělené entity a třídí je do skupin podle shody znaků.")
16
Podoba společenstev v prostoru
Mnohorozměrné statistiky Hledající vztahy mezi proměnnými prostředí a výskytem druhů. PCA CCA (Korespondeční analýza) A další V českých krajích oblíbené Canoco
 A další. V českých krajích oblíbené Canoco.")
17
Podoba společenstev v čase - SUKCESE
Sukcese je definována jako „nesezónní, směrovaný a kontinuální proces kolonizace a zániku populací jednotlivých druhů na určitém místě“. Tato obecná definice zahrnuje všemožné sukcesní sekvence v nejrůznějších časových škálách, a často i různých mechanismů. Degradační sukcese je sukcesí společenstev na mrtvé organické hmotě. Alogenní sukcese je sukcese vyvolaná vnějšími geochemickofyzikálními vlivy

18
Podoba společenstev v čase - SUKCESE
AUTOGENNÍ sukcese je sukcese, ke které dochází na nově vytvořených stanovištích, bez vnější změny abiotických podmínek. Můžeme rozlišit primární (úplně panenská půda –např. nový sopečný ostrov) sekundární (např. opuštěné pole) sukcesi. Raně sukcesní druh(y) – první kolonista(é) mohou tak změnit podmínky (zdroje), že usnadňují kolonizaci dalším druhům = facilitace. Jednou z principiálních změn je změna vlastností půdy (akumulace dusíku, humusu atp.). Směr sukcese ovšem může záležet na lokálních podmínkách mikrohabitatů (např. na svahu se nedrží voda les, na rovině se drží voda kyselé rašeliniště s borovicí). V některých situacích může dojít k opaku facilitace – k inhibici, to v případě, kdy úspěšný a rychlý kolonista zabere pro sebe rozhodující zdroje. (příklad Ulva nedovolí ruduchám osídlit příbojové skály). Photos courtesy of Dr. Brent McInnes, Chief Scientist from the Commonwealth Scientific and Industrial Research Organization (CSIRO) in Australia
 sekundární (např. opuštěné pole) sukcesi. Raně sukcesní druh(y) – první kolonista(é) mohou tak změnit podmínky (zdroje), že usnadňují kolonizaci dalším druhům = facilitace. Jednou z principiálních změn je změna vlastností půdy (akumulace dusíku, humusu atp.). Směr sukcese ovšem může záležet na lokálních podmínkách mikrohabitatů (např. na svahu se nedrží voda les, na rovině se drží voda kyselé rašeliniště s borovicí). V některých situacích může dojít k opaku facilitace – k inhibici, to v případě, kdy úspěšný a rychlý kolonista zabere pro sebe rozhodující zdroje. (příklad Ulva nedovolí ruduchám osídlit příbojové skály). Photos courtesy of Dr. Brent McInnes, Chief Scientist from the Commonwealth Scientific and Industrial Research Organization (CSIRO) in Australia.")
19
Erupted May 18 1980 AUTOGENNÍ PRIMÁRNÍ SUKCESE
The eruption of Mt. St. Helens created a huge landscape where in some regions, all vegetation and traces of vegetation were eliminated Actually created a wide number of substrates – pyroclastic flows,mudflows, blast deposits, debris avalanches Combined with variations in micro-climate associated with elevation and orographic rainfall, there are a very large number of micro-climates that occurred in this region Many of the sites are undergoing primary succession One of the unique things about many sites at MSH undergoing primary succession is the absence of adjacent seedbanks for recolonization In many cases of primary succession (those we will discuss later), sites that are created are actually adjacent to sites that are occupied by plant communities Key factor in primary succession (create list on board) Availability of seeds for colonization Dispersal capabilities and patterns are very important
, sites that are created are actually adjacent to sites that are occupied by plant communities. Key factor in primary succession (create list on board) Availability of seeds for colonization. Dispersal capabilities and patterns are very important.")
20
The rates of recolonization of mudflow deposits above Treeline at MSH have been extremely slow
Here we see that 17 years after the blast, there is very little vegetation present at these sites Importance of chance in early establishment Once established provides a local seed source, plus shelter and stablelized substrate.

21
AUTOGENNÍ PRIMÁRNÍ SUKCESE

22
20 km N Glacier Bay Alaska Glacier Bay Direction of glacial retreat
Očekávaná sukcesní řada: Půda exponovaná méně než 20 let: Vrby a Dryas 20 km Půdy exponované let: olše, roztroušeně topol N Půdy exponované 100 let: olše, Roztroušeně smrk Figure: 50.12a Caption: (a) Several distinctive plant communities are found in Glacier Bay, Alaska. The first hypothesis about succession at Glacier Bay proposed that these communities represented stages in the same successional pathway. Glacier Bay Půdy exponované let: Nahusto smrk (Picea sitchensis) a Tsuga Alaska Glacier Bay Direction of glacial retreat
 Several distinctive plant communities are found in Glacier Bay, Alaska. The first hypothesis about succession at Glacier Bay proposed that these communities represented stages in the same successional pathway. Glacier Bay. Půdy exponované let: Nahusto smrk (Picea sitchensis) a Tsuga. Alaska. Glacier. Bay. Direction of. glacial retreat.")
24
Primární sukcese po ústupu ledovce – viz předchozí obrázek
Holý ledovcový till Mechy, vrba, dryádka, vrbovka, topol během 1-10 prvních let Po deseti letech jsou tato stanoviště obsazena olší, která během 50 let doroste v hustém zápoji do výšky až 9 metrů Pak nastupuje smrk Picea sitchensis, který během dalších 120 let vytvoří hustý zapojený les Smrkové lesy jsou pak invadovány tsugou – po dalších 80letech je tak dosaženo (na dobře odvodňovaných místech) klimaxového stadia. Na zamokřených místech tvorba rašelinišť. Další možná varianta/vývoj – kterak se v (klimaxovém) lese hromadí mrtvá organická hmota (jehličí), tak se půdní horizont „utěsňuje“, a zhoršuje se odvodňování rašeliniště. What we are seeing on the glacial till sites is a distinct successional pattern or chronosequence that occurs over a period of about 250 to 300 years Pokles pH půdy Nárůst dusíku v půdě po kolonizaci olší Pokles dusíku v půdě po „vystřídání“ olše Zamokření a a okyselení tam, kde voda neotéká – obsazeno rašeliníkem
 klimaxového stadia. Na zamokřených místech tvorba rašelinišť. Další možná varianta/vývoj – kterak se v (klimaxovém) lese hromadí mrtvá organická hmota (jehličí), tak se půdní horizont „utěsňuje , a zhoršuje se odvodňování rašeliniště. What we are seeing on the glacial till sites is a distinct successional pattern or chronosequence that occurs over a period of about 250 to 300 years. Pokles pH půdy. Nárůst dusíku v půdě po kolonizaci olší. Pokles dusíku v půdě po „vystřídání olše. Zamokření a a okyselení tam, kde voda neotéká – obsazeno rašeliníkem.")
25
The soils of glacial tills are very basic because of the carbonates present from the parent rocks
However, leaves and vegetation, when decomposed, form acids, which when leached, decrease the acidity of the soil Alder leaves are particularly acidic, and after the alder forests form, there is a dramatic decrease in soil acidity Important point – additional of vegetation and their resulting decomposition results in acidification of the soil, a pattern found in most cases of primary succession

26
One of the key steps in succession at this site is the invasion by alders
Alders support root mychorizae in nodules, these mychorizae are nitrogen fixers Thus, the alder result in a tremendous increase in total nitrogen in both the organic soil (top soil) and the mineral soil This nitrogen addition facilitates the growth of other plant species Note that when the alder die out, and spruce come in, there is a decrease in soil nitrogen (nitrogen fixers are absent). However, after a point, the nitrogen content stabilizes Nitrogen is available to the system through decomposition of litter, e.g., dead plant material that is added to the forest floor Important point – nitrogen fixers are important components in early succession During primary succession, soil nutrients increase under all conditions
 and the mineral soil. This nitrogen addition facilitates the growth of other plant species. Note that when the alder die out, and spruce come in, there is a decrease in soil nitrogen (nitrogen fixers are absent). However, after a point, the nitrogen content stabilizes. Nitrogen is available to the system through decomposition of litter, e.g., dead plant material that is added to the forest floor. Important point – nitrogen fixers are important components in early succession. During primary succession, soil nutrients increase under all conditions.")
27
AUTOGENNÍ SEKUNDÁRNÍ SUKCESE
Staré pole Disturbance končí, stanoviště je osídleno krátkověkými druhy (letničkami) Pionýrské druhy Krátkověké druhy jsou vystřídány déle žijícími bylinami a trávami. Společenstvo ranné sukcese Začínají se uchycovat keře a krátkověké stromy. Střední sukcese Figure: 50.11 Caption: This sequence of photos shows how succession leads to the development of a temperate forest from a disturbed state (in this case, an agricultural field). Krátkověké stromy dospívají, začínají se uchycovat dlouhověké stromy. Pokročilá sukcese Dlouhověké stromy dospívají. Klimax
 Pionýrské druhy. Krátkověké druhy jsou vystřídány déle žijícími bylinami a trávami. Společenstvo. ranné sukcese. Začínají se uchycovat keře a krátkověké stromy. Střední sukcese. Figure: Caption: This sequence of photos shows how succession leads to the development of a temperate forest from a disturbed state (in this case, an agricultural field). Krátkověké stromy dospívají, začínají se uchycovat dlouhověké stromy. Pokročilá sukcese. Dlouhověké stromy dospívají. Klimax.")
28
AUTOGENNÍ SEKUNDÁRNÍ SUKCESE
What are some other examples of primary and secondary succession? Požár 1988 v Yellowstone AUTOGENNÍ SEKUNDÁRNÍ SUKCESE

29
Autogenní sukcese Příklady degradační sukcese a autogenní sukcese

30
Autogenní sukcese Typický sled dominantní vegetace na opuštěných polích jednoleté plevely bylinné trvalky keře raně sukcesní druhy stromů pozdně sukcesní druhy stromů. Při sledování sukcese na opuštěných polích amerického východu (farmáři šli dál na středozápad) jako první nastoupily pionýrské druhy, zejména ambrózie (Ambrosia) (ta potřebuje ke klíčení disturbanci – její semena „čekají“ v půdě, po expozici na povrchu rychle klíčí, pak nastoupily jednoleté ozimé rostliny (hlavně hvězdnicovité). Jejich růžice přezimují a na jaře už okupují zdroje (stíní). Ovšem pionýrské rostliny nevydrží v kompetici s dalšími – ke své realizaci potřebují stále nové „pionýrské“ plochy. Rostliny dalšího sledu mají jednu z výhod v tom, že tolerují větší zastínění. (viz obrázek po kliknutí). Jejich listoví je často mnohovrstevné. První kolonizující stromy mají oproti pozdějším často snadno se šířící malá semena. Pozdní kolonisté pak velká semena, delší juvenilní fázi a horší dispersi.
 jako první nastoupily pionýrské druhy, zejména ambrózie (Ambrosia) (ta potřebuje ke klíčení disturbanci – její semena „čekají v půdě, po expozici na povrchu rychle klíčí, pak nastoupily jednoleté ozimé rostliny (hlavně hvězdnicovité). Jejich růžice přezimují a na jaře už okupují zdroje (stíní). Ovšem pionýrské rostliny nevydrží v kompetici s dalšími – ke své realizaci potřebují stále nové „pionýrské plochy. Rostliny dalšího sledu mají jednu z výhod v tom, že tolerují větší zastínění. (viz obrázek po kliknutí). Jejich listoví je často mnohovrstevné. První kolonizující stromy mají oproti pozdějším často snadno se šířící malá semena. Pozdní kolonisté pak velká semena, delší juvenilní fázi a horší dispersi.")
31
Mechanismy v pozadí autogenní sukcese
(Pro hypotetické lesní společenstvo) Hornův maticový model vycházející ze dvou předpokladů potřebujeme vědět, jaká je pravděpodobnost nahrazení jednoho jedince určitého druhu jiným jedincem stejného nebo jiného druhu (za nějakou časovou jednotku). Potřebujeme vědět počáteční druhové složení Výsledek modelu (po kliknutí) – konečné složení (výsledek sukcese) je nezávislý na počátečních poměrech. Někde může fungovat, ale protože transientní funkce je pořád stejná a protože neuvažuje ani pořadí vstupu druhů, v mnoha případech fungovat nebude.
 Hornův maticový model vycházející ze dvou předpokladů. potřebujeme vědět, jaká je pravděpodobnost nahrazení jednoho jedince určitého druhu jiným jedincem stejného nebo jiného druhu (za nějakou časovou jednotku). Potřebujeme vědět počáteční druhové složení. Výsledek modelu (po kliknutí) – konečné složení (výsledek sukcese) je nezávislý na počátečních poměrech. Někde může fungovat, ale protože transientní funkce je pořád stejná a protože neuvažuje ani pořadí vstupu druhů, v mnoha případech fungovat nebude.")
32
Mechanismy v pozadí autogenní sukcese
Další tři modely kromě predikce i vysvětlují – Connel & Slayter. Pracují s facilitací, inhibicí a tolerancí. Schema viz Begon str. 641. V modelu facilitace první připravují cestu dalším (a zároveň zhoršují podmínky pro sebe sama), v modelu tolerance je sekvence druhů predikována jejich kompetičními schopnostmi: pozdní druhy jsou schopny tolerovat nízké úrovně zdrojů, přežít a nakonec v kompetici zvítězit, v modelu inhibice sice první druhy odolávají kompetici, ale vymírají vinou vnějších vlivů. Na uvolněné místo pak nastupují (v tuto chvíli již) kompetičně silnější druhy. Významným příspěvkem pro porozumnění sukcesi je i Tilmanova hypotéza o poměru zdrojů.
, v modelu tolerance je sekvence druhů predikována jejich kompetičními schopnostmi: pozdní druhy jsou schopny tolerovat nízké úrovně zdrojů, přežít a nakonec v kompetici zvítězit, v modelu inhibice sice první druhy odolávají kompetici, ale vymírají vinou vnějších vlivů. Na uvolněné místo pak nastupují (v tuto chvíli již) kompetičně silnější druhy. Významným příspěvkem pro porozumnění sukcesi je i Tilmanova hypotéza o poměru zdrojů.")
33
Mechanismy v pozadí autogenní sukcese
Dalším pokusem (Noble & Slayter) byla snaha determinovat ty vlastnosti, které určují pozici/úlohu daného druhu v sukcesní řadě = vitální atributy. Ze dvou nejvýznamnějších: metoda obnovení po disturbanci vegetativní šíření V obnovení z banky semen S obnovení z okolí účinnou disperzí D bez speciálního mechanismu, jen skromná disperse a malá semenná banka N schopnost reprodukce za podmínek kompetice tolerantní T netolerantní I Druh tak může být precizně klasifikován (např. plevel typu Ambrosie je SI, buk je VT nebo NT, a pokud tak oklasifikujeme všechny, měly by být možné přesné predikce.
 byla snaha determinovat ty vlastnosti, které určují pozici/úlohu daného druhu v sukcesní řadě = vitální atributy. Ze dvou nejvýznamnějších: metoda obnovení po disturbanci. vegetativní šíření V. obnovení z banky semen S. obnovení z okolí účinnou disperzí D. bez speciálního mechanismu, jen skromná disperse a malá semenná banka N. schopnost reprodukce za podmínek kompetice. tolerantní T. netolerantní I. Druh tak může být precizně klasifikován (např. plevel typu Ambrosie je SI, buk je VT nebo NT, a pokud tak oklasifikujeme všechny, měly by být možné přesné predikce.")
34
Mechanismy v pozadí autogenní sukcese
Určité vitální atributy se z hlediska působení evoluce budou vyskytovat pospolu – a jsme opět u r/K koncepce. Buď se druhy budou snažit kompetici vydržet (K) anebo utéct (r) ze sukcese jinam na místa bez kompetice = dobří kolonisté.
 anebo utéct (r) ze sukcese jinam na místa bez kompetice = dobří kolonisté.")
35
Sukcese Jak patrno z předchozího, sukcese se bere jak věc v zásadě botanická. Je to celkem pochopitelné, protože rostliny tvoří nejvíce biomasy a fyzickou podobu/strukturu mnoha společenstev. A taky se pomalu rozkládají, přitom ovšem nejvíce přispívají do nekromasy. Ta se při pomalejším rozkladu hromadí jako hrabanka nebo rašelina. Nicméně i dřevnaté kmeny a větve stromů jsou z většiny nekromasou, oporou pro aktivní moduly. U živočichů analogicky koráli. Ovšem i živočichové mohou směr sukcese ovlivňovat – třeba například rozdíl mezi působením listožravých a kořenožravých herbivorů na vytrvalé trávy vs. vytrvalé byliny. Když defoliace – rozvoj trav. Když po kořenech, tak rozvoj bylin. Nebo savana stromy x sloni. Ale mnohem častěji živočichové jen pasivně odrážejí rostlinnou sukcesi.

36
Klimax Má sukcese konec? Je jasné, že pokud vymírající jedinci budou nahrazováni jedinci toho samého druhu, bude vytvořena určitá stabilní rovnováha (viz Hornův model) Koncepce klimaxu jako konečného stadia sukcese má dlouhou historii. Frederick Clements (1916) hájil myšlenku jednoho typu klimaxu v rámci daného klimatické oblasti (čili by bylo jedno, jestli to začne na písečné duně, opuštěném poli nebo zazeměním tůně). Tuto myšlenku monoklimaxu řada jiných napadala, s tím, že podoba konečného stadia bude záležet na mnoha lokálních faktorech (polyklimax). V realitě je velmi těžké označit nějaký stav jako stabilní klimax. Už proto, že k nějakým změnám dochází neustále (přírodní katastrofy, polomy, požáry), nebo že se klima neustále vyvíjí (změny od poslední glaciace, globální oteplování). A pokud zmenšíme měřítko na úroveň „lesa za humny“, tak v něm se odehrává v každém okamžiku kvanta miniaturních sukcesí.
 Koncepce klimaxu jako konečného stadia sukcese má dlouhou historii. Frederick Clements (1916) hájil myšlenku jednoho typu klimaxu v rámci daného klimatické oblasti (čili by bylo jedno, jestli to začne na písečné duně, opuštěném poli nebo zazeměním tůně). Tuto myšlenku monoklimaxu řada jiných napadala, s tím, že podoba konečného stadia bude záležet na mnoha lokálních faktorech (polyklimax). V realitě je velmi těžké označit nějaký stav jako stabilní klimax. Už proto, že k nějakým změnám dochází neustále (přírodní katastrofy, polomy, požáry), nebo že se klima neustále vyvíjí (změny od poslední glaciace, globální oteplování). A pokud zmenšíme měřítko na úroveň „lesa za humny , tak v něm se odehrává v každém okamžiku kvanta miniaturních sukcesí.")
37
Faktory ovlivňující strukturu společenstva
Mezidruhová kompetice – může determinovat kolik a jakých druhů může koexistovat, viz přednáška o mezidruhové kompetici. Je ovšem otázkou, do jaké míry a také pro jaké skupiny je její efekt významný. Například ze studií fytofágního hmyzu (= cca 25% známých druhů organismů) jen 20% ukazuje vliv vnitrodruhové kompetice, což znamená, že mezidruhová kompetice je ještě mnohem slabší.
 jen 20% ukazuje vliv vnitrodruhové kompetice, což znamená, že mezidruhová kompetice je ještě mnohem slabší.")
38
Faktory ovlivňující strukturu společenstva
PREDACE a DISTURBANCE Disturbance – hovoříme-li o „vnějších vlivech“, „náhodných změnách“, ale též o antropogenním působení (zemědělství, lesní hospodářství, hnojení, znečišťování, rekreace, lov) – drží populace mimo intenzivní vliv kompetice, pod nosnou kapacitou. I v případě aktuální kompetice dochází během času k neustálým změnám stanoviště, což může bránit ustavení nějaké stabilní rovnováhy. DISTURBANCI je možno definovat jako změnu procesu nebo narušení ustáleného stavu. Z hlediska struktury společenstva je narušovaným procesem mezidruhová kompetice, a ustáleným stavem je taková struktura společenstva, které by dosáhlo při neměnnosti podmínek. Disturbance je tedy potom jakákoliv událost v čase, která odstraňuje organismy, nebo jinak mění společenstvo z hlediska dostupnosti prostoru, zdrojů nebo fyzikálních podmínek. Disturbance má za následek obvykle uvolnění prostoru nebo uvolnění zdrojů, které jsou pak k dispozici jiným organismům. PREDACE z hlediska svého působení na společenstvo výše uvedené podmínky velmi dobře splňuje.
 – drží populace mimo intenzivní vliv kompetice, pod nosnou kapacitou. I v případě aktuální kompetice dochází během času k neustálým změnám stanoviště, což může bránit ustavení nějaké stabilní rovnováhy. DISTURBANCI je možno definovat jako změnu procesu nebo narušení ustáleného stavu. Z hlediska struktury společenstva je narušovaným procesem mezidruhová kompetice, a ustáleným stavem je taková struktura společenstva, které by dosáhlo při neměnnosti podmínek. Disturbance je tedy potom jakákoliv událost v čase, která odstraňuje organismy, nebo jinak mění společenstvo z hlediska dostupnosti prostoru, zdrojů nebo fyzikálních podmínek. Disturbance má za následek obvykle uvolnění prostoru nebo uvolnění zdrojů, které jsou pak k dispozici jiným organismům. PREDACE z hlediska svého působení na společenstvo výše uvedené podmínky velmi dobře splňuje.")
39
Faktory ovlivňující strukturu společenstva
PREDACE a DISTURBANCE PREDACE z hlediska svého působení na společenstvo výše uvedené podmínky velmi dobře splňuje.

40
PREDACE – efekt pastvy Už Darwin (1859) si všiml, že sekaný trávník je druhově bohatší než trávník neudržovaný. Totéž třeba králíci introdukovaní do Británie, ale když moc silná predace, tak extinkce. Podobně i jiné příklady (plž Littorina a řasy, ovce a trávy)
 si všiml, že sekaný trávník je druhově bohatší než trávník neudržovaný. Totéž třeba králíci introdukovaní do Británie, ale když moc silná predace, tak extinkce. Podobně i jiné příklady (plž Littorina a řasy, ovce a trávy)")
41
PREDACE – efekt masožravých predátorů
Odstranění hvězdic ze skalních útesů – rozvoj kompetičně dominantních slávek a celkový počet počet druhů z 15 na 8. Podobný efekt silný příboj. PREDACE – efekt změny potravy a hustotně závislá selekce Predátor málokdy působí tak, že by nejdříve vyhubil jeden druh, a pak teprve se vrhnul na další. Obvykle žere víc druhů naráz, anebo s úbytkem jedno-ho typu kořisti se „skokem“ začne zajímat o další. Takové chování by teoreticky vedlo ke koexistenci velkého počtu rela-tivně vzácných druhů. V praxi je to pozorováno jen vzácně (ale je!). Naopak – je hodně vyhraněných specialistů, nebo jsou případy, kdy s introdukcí predátora do „nového“ prostředí dramaticky klesl počet druhů (Boiga irregularis na ostrovy v Ocenánii)
. Naopak – je hodně vyhraněných specialistů, nebo jsou případy, kdy s introdukcí predátora do „nového prostředí dramaticky klesl počet druhů (Boiga irregularis na ostrovy v Ocenánii)")
42
PARAZITI A NEMOCI Výskyt parazita anebo nemoci často rozhoduje o bytí a nebytí (výskytu) konkrétního hostitele na lokalitě. Až 50% všech ptačích druhů na Hawaiských ostrovech zmizelo vinou introdukce ptačích patogenů (malárie a ptačích nešťovic). Parazité – nemoci mohou zlikvidovat určitou populaci na rozsáhlém území. Ale je otázkou, jak je ovlivněna celková diversita – po ustanovení nové „stability“. K tomu je potřeba vylučovacích pokusů (podobně jako s predátory). Jiným způsobem je sledování vlivu nemoci/parazita ve zjednodušených společenstvech. Například oslabení kompetice jednoho z komeptitorů.
 konkrétního hostitele na lokalitě. Až 50% všech ptačích druhů na Hawaiských ostrovech zmizelo vinou introdukce ptačích patogenů (malárie a ptačích nešťovic). Parazité – nemoci mohou zlikvidovat určitou populaci na rozsáhlém území. Ale je otázkou, jak je ovlivněna celková diversita – po ustanovení nové „stability . K tomu je potřeba vylučovacích pokusů (podobně jako s predátory). Jiným způsobem je sledování vlivu nemoci/parazita ve zjednodušených společenstvech. Například oslabení kompetice jednoho z komeptitorů.")
43
PREDÁTOŘI, PARAZITI A NEMOCI - SOUHRN
Selektivní predátoři budou (pravděpodobně) zvyšovat diversitu v případě, že jejich kořist je kompetičně silná. Rovněž působení mnoha neselektivních predátorů (generalistů) má pravděpodobně za následek zvýšení diversity – i v tomto případě jsou silní kompetitoři postiženi relativně nejvíce. Nejvyšší diversitu můžeme očekávat při „střední“ intenzitě predace, protože příliš nízká intenzita nepotlačí silné kompetitory, a příliš intenzivní predace může preferovanou kořist vyhubit. Efekt predace, parazitace a nemocí na strukturu společenstva je patrně relativně méně intenzivní tam, kde jsou fyzické podmínky nepredikovatelné, variabilní a kruté. Působení živočichů na strukturu společenstva se neomezuje toliko na pouhé „odebírání“ organismů. Například živočichové žijící v půdě či hrabající nory vytvářejí „disturbance“, stejně tak velcí predátoři „vyrábí“ mozaiku lokálně bohatých stanovišť.
 zvyšovat diversitu v případě, že jejich kořist je kompetičně silná. Rovněž působení mnoha neselektivních predátorů (generalistů) má pravděpodobně za následek zvýšení diversity – i v tomto případě jsou silní kompetitoři postiženi relativně nejvíce. Nejvyšší diversitu můžeme očekávat při „střední intenzitě predace, protože příliš nízká intenzita nepotlačí silné kompetitory, a příliš intenzivní predace může preferovanou kořist vyhubit. Efekt predace, parazitace a nemocí na strukturu společenstva je patrně relativně méně intenzivní tam, kde jsou fyzické podmínky nepredikovatelné, variabilní a kruté. Působení živočichů na strukturu společenstva se neomezuje toliko na pouhé „odebírání organismů. Například živočichové žijící v půdě či hrabající nory vytvářejí „disturbance , stejně tak velcí predátoři „vyrábí mozaiku lokálně bohatých stanovišť.")
44
VARIACE PODMÍNEK V ČASE
Simulace Lotka-Volterra, periodické snižování o 50%.

45
VARIACE PODMÍNEK V ČASE
Simulace se 6 druhy –disturbance různě často (Huston 1979), prostřední frekvence nejvyšší diversita.
, prostřední frekvence nejvyšší diversita.")
46
VARIACE PODMÍINEK V ČASE
Logicky a teoreticky – rychlost kompetitivního vyloučení je vyšší v přípa-dech, kdy jsou vyšší sa-motné rychlosti růstu. Neboli – (nekonečná) koexistence je možná při určité frekvenci disturban-cí, ale jen pokud nejsou rychlosti růstu populace příliš vysoké. V realitě: často pozoruje-me pokles diversity (dru-hové bohatosti) na urči-tých produktivních stanovi-štích (hnojené pastviny, eutrofní jezera). Neprodu-ktivní stanoviště by na-opak měla mít vyšší diver-situ – tak tomu je třeba na živiny chudých stanoviš-tích ve Fybosu v Jižní Africe nebo v Austrálii.
 koexistence je možná při určité frekvenci disturban-cí, ale jen pokud nejsou rychlosti růstu populace příliš vysoké. V realitě: často pozoruje-me pokles diversity (dru-hové bohatosti) na urči-tých produktivních stanovi-štích (hnojené pastviny, eutrofní jezera). Neprodu-ktivní stanoviště by na-opak měla mít vyšší diver-situ – tak tomu je třeba na živiny chudých stanoviš-tích ve Fybosu v Jižní Africe nebo v Austrálii.")
47
DISTURBANCE A „PATCH-DYNAMIC“ koncept
Vznik „mezer“ (gaps, patches) je ve společenstvech velmi častým jevem, a je podstatný hlavně pro přisedlé, nemobilní organismy. Existuje řada modelů dívajících se na společenstvo jako na mozaiku stejných „patches“, které jsou náhodně osidlovány. Interakce mezi druhy (popisujeme pomocí Lotka-Volterra) vedou k lokálním kompetitivním vyloučením, ale protože možnost emigrace a imigrace mezi patches, v celku možná koexistence bez kompetitivního vyloučení. = OTEVŘENÝ SYSTĚM. Při tomto pohledu můžeme společenstva rozlišovat na regulovaná dominancí a regulovaná zakladatelem. Společenstva regulovaná DOMINANCÍ Společenstva regulovaná dominancí – druh kolonizující uvolněné místo na něm nemusí nutně vydržet, může být díky kompetici vytlačen a na tomto konkrétním místě „vyhynout“, ale na lokalitě jako celku díky migraci a rekolonizaci vydržet („pravděpodobnostní refugium“). V těchto společenstvech disturbance (jež otevírají nové „mezery“) vedou k víceméně předpověditelné sekvenci druhů (= minisukcese). Disturbance tedy vlastně vrací stanoviště do dřívějších sukcesních stadií .
 je ve společenstvech velmi častým jevem, a je podstatný hlavně pro přisedlé, nemobilní organismy. Existuje řada modelů dívajících se na společenstvo jako na mozaiku stejných „patches , které jsou náhodně osidlovány. Interakce mezi druhy (popisujeme pomocí Lotka-Volterra) vedou k lokálním kompetitivním vyloučením, ale protože možnost emigrace a imigrace mezi patches, v celku možná koexistence bez kompetitivního vyloučení. = OTEVŘENÝ SYSTĚM. Při tomto pohledu můžeme společenstva rozlišovat na regulovaná dominancí a regulovaná zakladatelem. Společenstva regulovaná DOMINANCÍ. Společenstva regulovaná dominancí – druh kolonizující uvolněné místo na něm nemusí nutně vydržet, může být díky kompetici vytlačen a na tomto konkrétním místě „vyhynout , ale na lokalitě jako celku díky migraci a rekolonizaci vydržet („pravděpodobnostní refugium ). V těchto společenstvech disturbance (jež otevírají nové „mezery ) vedou k víceméně předpověditelné sekvenci druhů (= minisukcese). Disturbance tedy vlastně vrací stanoviště do dřívějších sukcesních stadií .")
48
Společenstva regulovaná DOMINANCÍ
Frekvence tvoření „mezer“ a druhová bohatost Modelové předpovědi „nejlepšího“ výsledku střední frekvence disturbancí na druhovou pestrost potvrzují i pozorování a experimenty.

49
Společenstva regulovaná DOMINANCÍ
Různé velikosti „mezer“ a jejich zaplňování Velikost mezer ovlivňuje řadu věcí – například lze očekávat, že čím větší je „mezera“, tím větší šanci tam mají druhy s propagulemi šířenými na velké vzdálenosti. U malých „mezer“ je mobilita propagulí méně důležitá, protože stejně budou osidlovány z nejbližšího okolí. Příklad se slávkou – uvolněná místa od velikosti jedné škeble až po plochu 38m2. Místa menší než 100 cm2 jsou zaplněna škeblemi ze sousedství velmi rychle, větší místa (3500 cm2) až za několik let a proto invaze jiných druhů, slávka dominuje zase až po 4 letech. Na největších prázdných plochách se kolonizace děje přes larvy, a ty než vyrostou, trvá dlouho, a stejně tak dlouho trvá období vyšší diversity. Navlas stejné schéma u „mezer“ vytvořených „spálením“ vegetace zvířecí močí, nebo v okolí nor hlodavců.
 až za několik let a proto invaze jiných druhů, slávka dominuje zase až po 4 letech. Na největších prázdných plochách se kolonizace děje přes larvy, a ty než vyrostou, trvá dlouho, a stejně tak dlouho trvá období vyšší diversity. Navlas stejné schéma u „mezer vytvořených „spálením vegetace zvířecí močí, nebo v okolí nor hlodavců.")
50
Společenstva regulovaná ZAKLADATELEM
U předchozího typu (kontrolovaná dominancí) šlo o běžnou r- a K- dichotomii. Na zakladatelem regulovaných společenstvech se předpokládají druhy, které jsou nejen dobrými kolonizátory, ale také dobrými kompetitory. Proto, pokud se objeví „mezera“, nedá se očekávat sukcese. Pak záleží na pravděpodobnostech a náhodě – kompetitivní loterie – v případě, že se mezery objevují neustále a náhodně, udržuje se vysoká diversita. K tomu je další podmínkou stejnocenný přístup potomků na uvolněné místo (čili mělo by být jedno, kolik kdo jich má – pokud by to jedno nebylo, tak vyhraje ten s nejvíc.)
 šlo o běžnou r- a K- dichotomii. Na zakladatelem regulovaných společenstvech se předpokládají druhy, které jsou nejen dobrými kolonizátory, ale také dobrými kompetitory. Proto, pokud se objeví „mezera , nedá se očekávat sukcese. Pak záleží na pravděpodobnostech a náhodě – kompetitivní loterie – v případě, že se mezery objevují neustále a náhodně, udržuje se vysoká diversita. K tomu je další podmínkou stejnocenný přístup potomků na uvolněné místo (čili mělo by být jedno, kolik kdo jich má – pokud by to jedno nebylo, tak vyhraje ten s nejvíc.)")
51
Společenstva regulovaná ZAKLADATELEM
Někde to funguje, ale ne vždy jsou „mezery“ identické a ne vždy se objevují ve stejnou dobu, což je podstatné hlavně u mezer s krátkou nebo sezónní dobou výskytu.

52
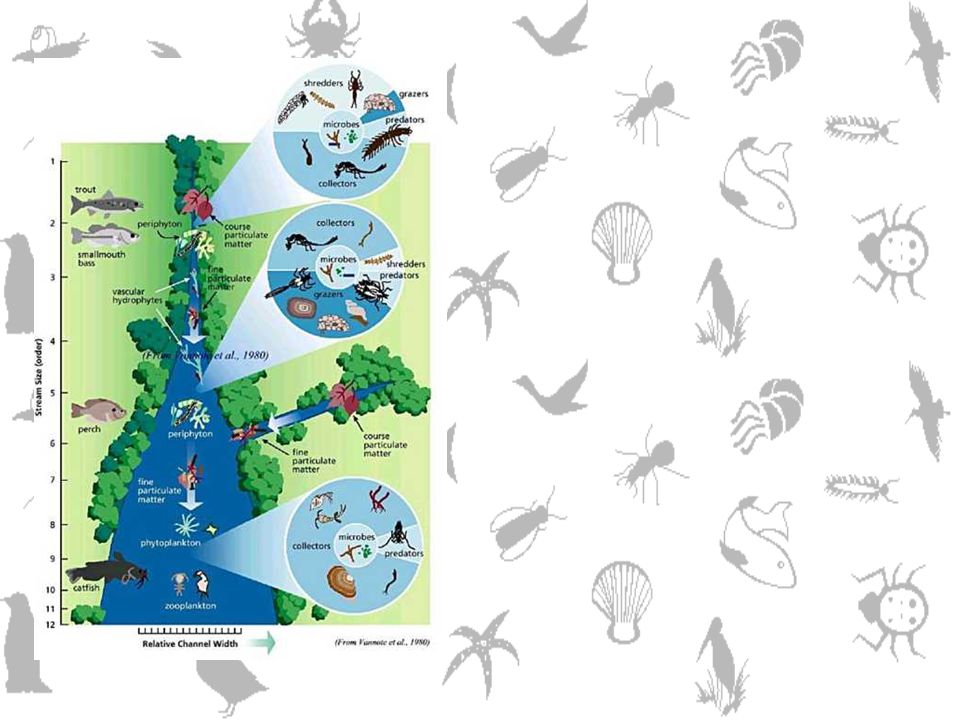
Potravní řetězce – food webs
Při popisu společenstva dosud v rámci vztahu kompetitor-kompetitor (stejná trofická úroveň) nebo predátor-kořist (sousedící trofické úrovně). Ale je jasné, že vliv interakcí zasahuje nepřímo, zprostředkovaně, i další organismy. Jak moc? Jsou v těchto vazbách nějaké druhy důležitější než jiné? Je struktura společenstva ovlivňována směrem od konzumenta k producentovi (top-down), nebo naopak (bottom-up)? Souvisí nějak struktura potravních řetězců (sítí) se stabilitou společenstva?
 nebo predátor-kořist (sousedící trofické úrovně). Ale je jasné, že vliv interakcí zasahuje nepřímo, zprostředkovaně, i další organismy. Jak moc Jsou v těchto vazbách nějaké druhy důležitější než jiné Je struktura společenstva ovlivňována směrem od konzumenta k producentovi (top-down), nebo naopak (bottom-up) Souvisí nějak struktura potravních řetězců (sítí) se stabilitou společenstva")
53
Potravní řetězce – food webs
Nepřímé efekty: Přímé jsou zřejmé – odstraníme-li predátora, můžeme očekávat nárůst jeho kořisti. Odstraníme-li kompetitora, vzroste počet jeho soupeře. Ovšem někdy tomu může být naopak – po odstranění kompetitora klesne početnost jeho konkurenta. Podobně s predátorem. To v případě, že existuje nějaký třetí (a další) člen v těchto vztazích, a že jeho interakce s kompetirorem (kořistí) je silnější než ta přímo pozorovaná. Klíčové druhy – keystone species Některé druhy jsou v potravních sítích „zamotány pevněji“ než jiné – jsou silnými „interaktory“. Proto jejich odstranění způsobí významné změny šířící se celou potravní sítí (viz příklad hvězdic na útesech, Paine, 1966). Predátoři jsou jasní, ale co třeba rostliny jako keystones? Když odstraníme duby z dubového lesa, tak se určitě drasticky změní celé společenstvo. Stejně tak je najdeme mezi herbivory, nebo detritovory.
 člen v těchto vztazích, a že jeho interakce s kompetirorem (kořistí) je silnější než ta přímo pozorovaná. Klíčové druhy – keystone species. Některé druhy jsou v potravních sítích „zamotány pevněji než jiné – jsou silnými „interaktory . Proto jejich odstranění způsobí významné změny šířící se celou potravní sítí (viz příklad hvězdic na útesech, Paine, 1966). Predátoři jsou jasní, ale co třeba rostliny jako keystones Když odstraníme duby z dubového lesa, tak se určitě drasticky změní celé společenstvo. Stejně tak je najdeme mezi herbivory, nebo detritovory.")
54
Potravní řetězce – food webs
Přímé a nepřímé efekty jsou pozorovatelné v rámci více trofických úrovní naráz: pokus s odrušením vlivu ptačích predátorů (racků a ústřičníků) na přílipky a následně řasy. Tři druhy přílipek – světlá (sedá na vilejše (jsou světlí), tmavá, sedá na slávky (jsou tmavé), a třetí, ta si nevybírá). Když bez predace, tak prostor pro vilejše a tím i pro „jejich“ přílipku = celkově víc přílipek, a protože ty žerou řasy, tak méně řas.
 na přílipky a následně řasy. Tři druhy přílipek – světlá (sedá na vilejše (jsou světlí), tmavá, sedá na slávky (jsou tmavé), a třetí, ta si nevybírá). Když bez predace, tak prostor pro vilejše a tím i pro „jejich přílipku = celkově víc přílipek, a protože ty žerou řasy, tak méně řas.")
55
Potravní řetězce – food webs
TOP-DOWN nebo BOTTOM-UP? Na předchozím příkladu jasný příklad kontroly struktury společenstva směrem shora dolů (od nejvyšší trofické hladiny k nižším) = top-down. Klíčový rys systémů determinovaných predátorem Kdybychom se ovšem na společenstvo podívali z hlediska toku energie a biomasy, tak dostaneme opačný obrázek – bottom-up. Klíčový rys systémů limitovaných dostupností zdrojů, tedy populace uvnitř jedné trofické hladiny jsou kontrolovány kompeticí, nikoliv predací. Povídání k obrázku následuje na další straně
 = top-down. Klíčový rys systémů determinovaných predátorem. Kdybychom se ovšem na společenstvo podívali z hlediska toku energie a biomasy, tak dostaneme opačný obrázek – bottom-up. Klíčový rys systémů limitovaných dostupností zdrojů, tedy populace uvnitř jedné trofické hladiny jsou kontrolovány kompeticí, nikoliv predací. Povídání k obrázku následuje na další straně.")
56
TOP-DOWN nebo BOTTOM-UP?
Vliv počtu trofických hladin Jedna trofická hladina (pouze producenti): Z definice chybí predátor, a proto determinovaná dostupností zdrojů - kompeticí. Dvě trofické hladiny: Na ostrovech – obří želvy a rostliny – v ohradách bez želv mnohem větší diversita a biomasa. A jiné příklady. Tři trofické hladiny: fytoplankton – zooplankton – planktonožravé ryby. Normálně zooplankton „drží na uzdě“ fytoplankton, ale když ryby žerou zooplankton, tak fytoplankton omezen zdroji. Podobný příklad s artemií a dravým hmyzem – viz obrázek. Čtyři trofické hladiny fytoplankton – zooplankton – planktonožravé ryby – dravé ryby
: Z definice chybí predátor, a proto determinovaná dostupností. zdrojů - kompeticí. Dvě trofické hladiny: Na ostrovech – obří želvy a rostliny – v ohradách bez želv mnohem větší diversita a biomasa. A jiné příklady. Tři trofické hladiny: fytoplankton – zooplankton – planktonožravé ryby. Normálně zooplankton „drží na uzdě fytoplankton, ale když ryby žerou zooplankton, tak fytoplankton omezen zdroji. Podobný příklad s artemií a dravým hmyzem – viz obrázek. Čtyři trofické hladiny. fytoplankton – zooplankton – planktonožravé ryby – dravé ryby.")
57
BOTTOM UP TOP DOWN TOP-DOWN Podle (ne)přítomnosti / množství predátora na vrcholu se biomasa „přelévá“ z jedné trofické hladiny do druhé = kaskádový efekt P, N

58
Globálně spíš TOP-DOWN nebo BOTTOM-UP?
Země je zelená Hairston et al.(1960) – „rostlinná biomasa se může kumulovat a udržet, protože predátoři drží herbivory na uzdě“. = třístupňový systém. Murdoch (1966) – Země je zelená, protože „je ostnitá a nechutná“ Oksanen (1988) – ... někdy je taky bílá. (na neproduktivních stanovištích není co žrát) Zdá se ovšem, že neplatí, že více produktivní společenstva mohou „unést“ více trofických skupin. pro rozhodnutí zda B-U nebo T-D je klíčová úroveň primární produkce
 – „rostlinná biomasa se může kumulovat a udržet, protože predátoři drží herbivory na uzdě . = třístupňový systém. Murdoch (1966) – Země je zelená, protože „je ostnitá a nechutná Oksanen (1988) – ... někdy je taky bílá. (na neproduktivních stanovištích není co žrát) Zdá se ovšem, že neplatí, že více produktivní společenstva mohou „unést více trofických skupin. pro rozhodnutí zda B-U nebo T-D je klíčová úroveň primární produkce.")
59
STABILITA SPOLEČENSTVA a potravní řetězce
Stabilita má různé komponenty Pružnost (resilience) – rychlost návrácení se do původního stavu. Odolnost (resistance) – schopnost vyhnout se (odolat) narušení. Lokální x globální stabilita – reakce na malé x velké narušení. Křehkost x pevnost společenstvo, které je stabilní pouze v rámci úzkého rozmezí nějakých podmínek (ať už faktorů prostředí, zdrojů, nebo přítomnosti určitých organismů), je tzv. dynamicky křehké. Naopak společenstvo fungující v širokém rozmezí podmínek je dynamicky robustní – pevné. obrázek 23.1 český begon (22.5. angl) VIDEO
 – rychlost návrácení se do původního stavu. Odolnost (resistance) – schopnost vyhnout se (odolat) narušení. Lokální x globální stabilita – reakce na malé x velké narušení. Křehkost x pevnost. společenstvo, které je stabilní pouze v rámci úzkého rozmezí nějakých podmínek (ať už faktorů prostředí, zdrojů, nebo přítomnosti určitých organismů), je tzv. dynamicky křehké. Naopak společenstvo fungující v širokém rozmezí podmínek je dynamicky robustní – pevné. obrázek 23.1 český begon (22.5. angl) VIDEO.")
60
STABILITA SPOLEČENSTVA a potravní řetězce
V ekologii dlouho existovala (a v obecném povědomí stále existuje) konvenční představa, že čím je společenstvo složitější/komplexnější, tím je také stabilnější. (?) První to formuloval Elton, po něm trochu jinak McArthur – tok energie je rozdělen do více (mnoha) drah, proto když se jedna přeruší, tak se celek nezhroutí. Ale dnešní studie a modely tuto představu nepotvrzují: Pro modeláře je stabilita x komplexita velmi oblíbené téma, proto modelů celá řada. Typickým představitelem je třeba Mayův (1972) model zahrnující mnoho druhů, který hodnotí, jak kolísá velikosti populace každého druhu okolo své stabilní početnosti (lokální stabilita). Každý druh byl ovlivněn interakcemi s ostatními druhy – míra interakce (vliv početnosti druhu j na růstovou rychlost druhu i) se měřila pomocí parametru β. βij je nulová, pokud interakce nemá žádný efekt, pro kompetující druhy bude βij a βji negativní, pro predační vztah bude pro predátora (i) βij pozitivní a pro kořist (j) βji negativní. May „náhodně sestavil“ potravní síť a parametry samoregulačních vztahů βii a βjj nastavil na –1, a ostatní náhodně, zahrnující i nulové hodnoty. Model pak měl tři parametry: počet druhu S, propojenost C a β jako průměrná hodnota interakce (bez ohledu na znaménko). Výsledná „společenstva – potravní sítě“ by mohla být stabilní pouze pokud β(SC)1/2 < 1. Takový výsledek znamená, že zvýšení S nebo C nebo β zvyšuje nestabilitu! Ale třeba je takto založený model nerealistický, umělý.
 konvenční představa, že čím je společenstvo složitější/komplexnější, tím je také stabilnější. ( ) První to formuloval Elton, po něm trochu jinak McArthur – tok energie je rozdělen do více (mnoha) drah, proto když se jedna přeruší, tak se celek nezhroutí. Ale dnešní studie a modely tuto představu nepotvrzují: Pro modeláře je stabilita x komplexita velmi oblíbené téma, proto modelů celá řada. Typickým představitelem je třeba Mayův (1972) model zahrnující mnoho druhů, který hodnotí, jak kolísá velikosti populace každého druhu okolo své stabilní početnosti (lokální stabilita). Každý druh byl ovlivněn interakcemi s ostatními druhy – míra interakce (vliv početnosti druhu j na růstovou rychlost druhu i) se měřila pomocí parametru β. βij je nulová, pokud interakce nemá žádný efekt, pro kompetující druhy bude βij a βji negativní, pro predační vztah bude pro predátora (i) βij pozitivní a pro kořist (j) βji negativní. May „náhodně sestavil potravní síť a parametry samoregulačních vztahů βii a βjj nastavil na –1, a ostatní náhodně, zahrnující i nulové hodnoty. Model pak měl tři parametry: počet druhu S, propojenost C a β jako průměrná hodnota interakce (bez ohledu na znaménko). Výsledná „společenstva – potravní sítě by mohla být stabilní pouze pokud β(SC)1/2 < 1. Takový výsledek znamená, že zvýšení S nebo C nebo β zvyšuje nestabilitu! Ale třeba je takto založený model nerealistický, umělý.")
61
STABILITA SPOLEČENSTVA a potravní řetězce
Ale třeba je takto založený model nerealistický, umělý, protože vazby jsou náhodně sestavené – tj. mohou tam být biologické nesmysly (predátor bez kořisti, nebo A žere B žere C žere A). Pokud se model v tomto smyslu opraví, tak dává o něco stabilnější výsledky a také přechod od stability k nestabilitě není tak vyhraněný (jako v předchozím případě). Pokud se model upraví i v tom smyslu, že konzument neovlivňuje početnost zdroje (βij > 0, ale βji = 0), tak výsledkem modelu je systém se stabilitou buď neovlivněnou, nebo dokonce rostoucí (společenstvo kontrolované dárcem – detritovoři, ale též možná hmyz živící se nektarem, nebo parazitoidi, nebo různý fytofágní hmyz). Modelů je celá řada a ve své většině předpovídají pokles stability s rostoucí složitostí společenstva Ale modely pracují s náhodně vytvořenými společenstvy, zatímco reálná společenstva jsou jistě nenáhodná. V realitě se parametry modelu velmi špatně měří. Proto se bere β jako konstanta, a hodnotí se jen součin SC. Podle modelu by mělo platit, že aby byla stabilita zachována, musela by se se zvýšením počtu druhů snížit propojenost C. Jinými slovy – společenstva s mnoha druhy by měly mít méně vnitřních vazeb. Obrázek ukazuje protichůdné výsledky.
. Pokud se model v tomto smyslu opraví, tak dává o něco stabilnější výsledky a také přechod od stability k nestabilitě není tak vyhraněný (jako v předchozím případě). Pokud se model upraví i v tom smyslu, že konzument neovlivňuje početnost zdroje (βij > 0, ale βji = 0), tak výsledkem modelu je systém se stabilitou buď neovlivněnou, nebo dokonce rostoucí (společenstvo kontrolované dárcem – detritovoři, ale též možná hmyz živící se nektarem, nebo parazitoidi, nebo různý fytofágní hmyz). Modelů je celá řada a ve své většině předpovídají pokles stability s rostoucí složitostí společenstva. Ale modely pracují s náhodně vytvořenými společenstvy, zatímco reálná společenstva jsou jistě nenáhodná. V realitě se parametry modelu velmi špatně měří. Proto se bere β jako konstanta, a hodnotí se jen součin SC. Podle modelu by mělo platit, že aby byla stabilita zachována, musela by se se zvýšením počtu druhů snížit propojenost C. Jinými slovy – společenstva s mnoha druhy by měly mít méně vnitřních vazeb. Obrázek ukazuje protichůdné výsledky.")
62
STABILITA SPOLEČENSTVA a potravní řetězce
Takže co si o tom vlastně myslet? Není to moc jasné. Zdá se, že složitější společenstva jsou celkově méně stabilní. A také se zdá, že složitá, avšak křehká společenstva obývají relativně stabilnější prostředí, a jsou náchylnější k „vykolejení“ vnějším, nepřirozeným narušením více než jednodušší, pevnější společenstva, která jsou na disturbance „více zvyklá“ Pravděpodobně bude též důležitá i paralela mezi vlastnostmi společenstva a vlastnostmi jeho populací. Ve stabilních společenstvech budou spíše K-druhy , zatímco ve variabilních podmínkách spíše r-druhy. U K-druhů se dá očekávat, že jsou odolnější vůči narušení (jsou „silné a otužené“ z kompetice) , ale jakmile už narušeny jsou, mají malou pružnost. U r-druhů naopak, nechají se snáze „vykolejit“, ale rychle se vrátí zpět.
 , ale jakmile už narušeny jsou, mají malou pružnost. U r-druhů naopak, nechají se snáze „vykolejit , ale rychle se vrátí zpět.")
63
EMPIRICKÉ VLASTNOSTI POTRAVNÍCH SÍTÍ
POČET TROFICKÝCH ÚROVNÍ je důležitým parametrem trofických sítí. V ukázaném příkladě se pohybuje mezi 3 a 4 (průměr = 3.32) Klasickou otázkou je „kolik je možných trofických hladin?“. V přírodě typicky mezi 2 a 5. Proto další otázka – PROČ TAK MÁLO?
 Klasickou otázkou je „kolik je možných trofických hladin . V přírodě typicky mezi 2 a 5. Proto další otázka – PROČ TAK MÁLO")
64
VLASTNOSTI POTRAVNÍCH SÍTÍ - POČET TROFICKÝCH HLADIN
1. Málo jich je vinou nízké účinnosti přenosu energie na vyšší hladinu. Jen malá část radiační energie se váže fotosyntézou a je pak dostupná pro herbivory či detritovory. Navíc množství reálně konzumované je mnohem menší (hodně energie padne na metabolismus producentů). Mezi heterotrofy je účinnost nízká, od max. 30% k pouhému 1% v rámci jedné vazby. Je tedy možné, že na další potenciální trofickou hladinu se prostě energie už nedostává. ALE lze to empiricky testovat, protože v takovém případě by měly mít produktivní společenstva víc článků než neproduktivní (např. oligotrofní x eutrofní jezera či moře, pouště x tropy). Tak tomu ovšem není (snad až na výjimky). 2. Málo jich je proto, že dlouhé řetězce jsou méně pružné vůči disturbanci Modelování (pomocí Lotka-Voltera) ukazuje, že dlouhé řetězce jsou méně stabilní a daleko více fluktuují, až k vyhynutí vrcholných predátorů. Proto čím více článků, tím je potřeba stabilnější podmínky, a takové nejsou v realitě běžně k mání. K tomu obrázek (click). 3. Málo jich je proto, že design a chování predátora jsou omezeny vlastnostmi reálného světa. Lze předpokládat, že predátor by měl být větší než kořist (aby ji zdolal, ale neplatí universálně, viz fytofágní hmyz). Proto by se s každou úrovní měl příslušně zvětšovat, a lze čekat, že narazí na fyzické a fyzikální meze. Viz (např.) kniha „Proč má slon velké uši“.
. Mezi heterotrofy je účinnost nízká, od max. 30% k pouhému 1% v rámci jedné vazby. Je tedy možné, že na další potenciální trofickou hladinu se prostě energie už nedostává. ALE lze to empiricky testovat, protože v takovém případě by měly mít produktivní společenstva víc článků než neproduktivní (např. oligotrofní x eutrofní jezera či moře, pouště x tropy). Tak tomu ovšem není (snad až na výjimky). 2. Málo jich je proto, že dlouhé řetězce jsou méně pružné vůči disturbanci. Modelování (pomocí Lotka-Voltera) ukazuje, že dlouhé řetězce jsou méně stabilní a daleko více fluktuují, až k vyhynutí vrcholných predátorů. Proto čím více článků, tím je potřeba stabilnější podmínky, a takové nejsou v realitě běžně k mání. K tomu obrázek (click). 3. Málo jich je proto, že design a chování predátora jsou omezeny vlastnostmi reálného světa. Lze předpokládat, že predátor by měl být větší než kořist (aby ji zdolal, ale neplatí universálně, viz fytofágní hmyz). Proto by se s každou úrovní měl příslušně zvětšovat, a lze čekat, že narazí na fyzické a fyzikální meze. Viz (např.) kniha „Proč má slon velké uši .")
65
VLASTNOSTI POTRAVNÍCH SÍTÍ - POČET TROFICKÝCH HLADIN
4. Je jich málo, protože „predátor si vybírá nejvýhodnější kořist“ Vcelku logická úvaha: proč by evoluce měla podporovat vznik superpredátora, který bude umět „lovit lvy“, když i pro takového predátora bude zcela určitě výhodnější (= méně energeticky namáhavé) lovit tu samou kořist co lev; je ji víc, a oproti lvům se bude o poznání snadněji ulovitelná.
 lovit tu samou kořist co lev; je ji víc, a oproti lvům se bude o poznání snadněji ulovitelná.")
66
VLASTNOSTI POTRAVNÍCH SÍTÍ
Pružnost společenstva může záviset na rychlosti toku energie a pohybu živin. Modelově O‘Neil Čím je tok vyšší, tím rychleji se následky disturbance ze systému „vypláchnou“.

Podobné prezentace














 douda@fzp.czu.cz.>")





