Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Bioelektrické jevy a jejich měření
1. přednáška

2
Doporučená literatura
From Neuron to Brain (John. G. Nichols a kol.) Přehled lékařské fyziologie (W. Ganong) Lékařská fyziologie (Trojan a kol.) Molekulární podstata buněčné dráždivosti (F. Vyskočil a L. Vyklický Jr.) Biofyzikální chemie (V. Karpenko a M. Kodíček) Základy fysikální chemie (R. Brdička, J. Dvořák) Fyzikální chemie pro biologické vědy (Z. Vodrážka) Fyzikální chemie (W. Moore) přednášky
 Přehled lékařské fyziologie (W. Ganong) Lékařská fyziologie (Trojan a kol.) Molekulární podstata buněčné dráždivosti (F. Vyskočil a L. Vyklický Jr.) Biofyzikální chemie (V. Karpenko a M. Kodíček) Základy fysikální chemie (R. Brdička, J. Dvořák) Fyzikální chemie pro biologické vědy (Z. Vodrážka) Fyzikální chemie (W. Moore) přednášky.")
4
Bioelektrické jevy jsou integrální projev živých organismů, a to nejen vzrušivých tkání, ale všech živých buněk. Základem bioelektrických jevů jsou toky iontů (přenos náboje) Jde o toky iontů přes iontové kanály nebo toky aktivním přenosem (pumpy); důsledkem jsou polarizace různých struktur (např. tkání: stěna střeva aktivně transportující ionty z jedné strany na druhou; ale i polarizace jednotlivých buněk, celých orgánů a celých živých organismů) existence membrán Zásadní metodou studia bioelektrických jevů je elektrofyziologie. Elektrofyziologie zkoumá elektrické stavy živých organismů (polarizace buněk, tkání...) mechanismy vzniku a zániku napětí a proudů v těchto strukturách vliv elektřiny na živou hmotu elektrické charakteristiky biologických objektů a struktur (vodivost iontových kanálů, elektrogenní příspěvky iontových pump...) Aplikovaná elektrofyziologie využívá měření elektrických projevů živých systémů k funkční, morfologické a anatomické analýze zkoumaných systémů (EEG, EMG, ERG, EKG, ENG).
 Jde o toky iontů přes iontové kanály nebo toky aktivním přenosem (pumpy); důsledkem jsou polarizace různých struktur (např. tkání: stěna střeva aktivně transportující ionty z jedné strany na druhou; ale i polarizace jednotlivých buněk, celých orgánů a celých živých organismů) existence membrán. Zásadní metodou studia bioelektrických jevů je elektrofyziologie. Elektrofyziologie zkoumá. elektrické stavy živých organismů (polarizace buněk, tkání...) mechanismy vzniku a zániku napětí a proudů v těchto strukturách. vliv elektřiny na živou hmotu. elektrické charakteristiky biologických objektů a struktur (vodivost iontových kanálů, elektrogenní příspěvky iontových pump...) Aplikovaná elektrofyziologie využívá měření elektrických projevů živých systémů k funkční, morfologické a anatomické analýze zkoumaných systémů (EEG, EMG, ERG, EKG, ENG).")
5
Historické základy elektrofysiologie
„Již staří Řekové...“ polovina 18. století – Stephen Gray – popis elektrické indukce; nervový přenos může být elektrickým projevem, člověk je prostorový vodič; : žáček Charterhouse School zavěšen na popruhy z hedvábí, skleněnou tyčí přiloženou u nohou naindukován záporný náboj na nekrytých částech chlapce, ty začaly přitahovat kousky sušených lístků trávy , dtto Charles François de Cisternay du Fay a (Francie) či Christian Augustus Hausen (raději děvčata, Lipsko, Německo) 1745 – první elektrický kondensátor (Leyden jar, Lydenská láhev): sklenice naplněná vodou a obalená tenkou kovovou fólií uvolnění většího náboje pokusy Jeana Antiona Nolleta s velkým počtem osob zapojených do obvodu
 či Christian Augustus Hausen (raději děvčata, Lipsko, Německo) 1745 – první elektrický kondensátor (Leyden jar, Lydenská láhev): sklenice naplněná vodou a obalená tenkou kovovou fólií uvolnění většího náboje pokusy Jeana Antiona Nolleta s velkým počtem osob zapojených do obvodu.")
6
1780 – Luigi Galvani primitivní elektrický obvod s kovovými elektrodami spojenými žabím nervosvalovým preparátem Pro tento jev razil pojem „živočišná elektřina“. Uznával i „umělou elektřinu“ či „vzdušnou elektřinu“ (žabí svaly stimulovány uměle kovovými drátky na zábradlí balkónu, dlouhý drát trčící z páteřího kanálu k nebi záškuby stehýnek po úderu blesku ). „I dissected and prepared a frog and laid it on a table, on which, at some distance to the frog was an electric machine. It happened by chance that one of may assistants touched the inner crural nerve of the frog with the point of scalpel: whereupon at once the muscles of the limbs were violently convulsed. Another of those who used to help me in the electrical experiments thought he had noticed that at this instant a spark was drawn from the conductor of the machine. I myself was at the time occupied with a totally different matter; but when he drew my attention to this, I greatly desired to try it myself and discover the hidden principle.“
. „I dissected and prepared a frog and laid it on a table, on which, at some distance to the frog was an electric machine. It happened by chance that one of may assistants touched the inner crural nerve of the frog with the point of scalpel: whereupon at once the muscles of the limbs were violently convulsed. Another of those who used to help me in the electrical experiments thought he had noticed that at this instant a spark was drawn from the conductor of the machine. I myself was at the time occupied with a totally different matter; but when he drew my attention to this, I greatly desired to try it myself and discover the hidden principle.")
7
Vzdušná (bouřková) elektřina): dlouhý drát trčící z páteřího kanálu k nebi záškuby stehýnek po úderu blesku ).
 elektřina): dlouhý drát trčící z páteřího kanálu k nebi záškuby stehýnek po úderu blesku ).")
8
Giovanni Aldini – demonstrace „živočišné elektřiny“ na tělech popravených zločinců

9
Allessandro Guiseppe Antonio Anastasio Volta
1791 – polemika Galvani - Volta Galvani: záškuby žabích stehýnek jsou projevem bioelektřiny v živé struktuře x Volta: důvod záškubů je čistě fyzikálně-chemický, kdy v důsledku kontaktu dvou kovů vzniká tzv. voltův potenciál 1800 – v podstatě jako důsledek sporů s Galvanim vyvinul Volta tzv. voltův článek: první zdroj stabilního elektrického proudu; jeho zdrojem je vodivé spojení dvou rozdílných kovů (Zn/Ag) Nejprve používal jednotlivé obvody spojené do série, kdy obvodem byl pohár na víno naplněný roztokem soli, do něhož byly ponořeny elektrody. Později použil jako pojítko hmotu nasáklou tímto roztokem a sestrojoval sloupcové články; velikost proudu limitovala výška sloupců.
 Nejprve používal jednotlivé obvody spojené do série, kdy obvodem byl pohár na víno naplněný roztokem soli, do něhož byly ponořeny elektrody. Později použil jako pojítko hmotu nasáklou tímto roztokem a sestrojoval sloupcové články; velikost proudu limitovala výška sloupců.")
10
1819 (1820) – Hans Christian Øersted – dánský fyzik a chemik, konstrukce galvanometru, objevil magnetické účinky elektrického proudu; oersted – jednotka magnetické indukce 1825 – Leopold Nobili – prokázal toky elektrických proudů ze svalů do míchy žáby či z jednoho žabího těla spojeného s druhým ne dráty, ale bavlnou namočenou ve fysiologickém roztoku (podpora „živočišné elektřiny); konstrukce vlastního (astatického) galvanometru 1845 – Carlo Matteuci [-uči] – Nobiliho žák, preparát „rheostatické žáby“: podráždil svalová vlákna, na nich ležel vypreparovaný nerv jiného svalu (měl nervosvalový preparát jako citlivý bioindikátor přítomnosti biopotenicálů v jiném kontrahujícím svalu); ukázal, že každý úder srdce doprovází elektrický proud; pokus o měření vedení nervů (málo citlivý galvanometr)
; konstrukce vlastního (astatického) galvanometru – Carlo Matteuci [-uči] – Nobiliho žák, preparát „rheostatické žáby : podráždil svalová vlákna, na nich ležel vypreparovaný nerv jiného svalu (měl nervosvalový preparát jako citlivý bioindikátor přítomnosti biopotenicálů v jiném kontrahujícím svalu); ukázal, že každý úder srdce doprovází elektrický proud; pokus o měření vedení nervů (málo citlivý galvanometr)")
11
Živočišná elektřina: „sloup“ rozpůlených stehýnek Carla Matteuciho
Elektroda umístěná na intaktním povrchu byla spojena s elektrodou na povrchu rozříznutém: proud měřený ve svalech nevznikal kontaktem kovových elektrod galvanometru a žabí tkáně. Jeho intenzita rostla s počtem půlstehýnek, zapojených do sloupce = do série, zatímco počet kontaktů kov-sval zůstával stejný.

12
1849 – Emil Du Bois-Reymon [di boá rejmon, vlevo] – vyvinul techniku stimulace a registrace pomocí galvanometru vlastní výroby (5 km drátu, 24 tisíc otáček); demonstroval klidový membránový potenciál a akční potenciál ve svalu 1868 – Hermann von Helmholtz – změřil rychlost šíření akčního potenciálu v sedacím nervu žáby (cca 30 m/s) vs – m/s - 1872 (1873) – Gabriel Lippmann – kapilární elektrometr a rtuťový galvanometr, jemnější studie (tenká skleněná trubička naplněná rtutí převrstvenou kyselinou sírovou; meniskus rtuti se pohybuje v závislosti na elektrickém proudu), NC za fyziku 1908 1883 N.E. Vvedensky – užil zrcátkový galvanometr měřící proudy až v rozsahu pA, malý pohyb cívky galvanometru přenášel na vzdálené stínítko, pohled se zvětšil; studoval aktivitu lidských svalů (telefon pro poslech AP, 1884) 1902 – Julius Bernstein – biopotenciály nervových a svalových buněk existují díky membráně, která je selektivně propustná pro draslík, kvantitativně odpovídající Nernstově vztahu (tehdy mluvil ještě o hypotetické membráně) – v tom se ale mýlil 1902 – Ernest Overton – svaly ztrácí dráždivost v roztocích bez sodných iontů, hypotéza výměny intracelulárního draslíku za extracelulární sodík (měnil složení ECS) 1910 – J. Bernstein a L. German popsali nervový impuls jako „putující potenciál aktivity“
![1849 – Emil Du Bois-Reymon [di boá rejmon, vlevo] – vyvinul techniku stimulace a registrace pomocí galvanometru vlastní výroby (5 km drátu, 24 tisíc otáček); demonstroval klidový membránový potenciál a akční potenciál ve svalu](http://slideplayer.cz/slide/3390207/11/images/12/1849+%E2%80%93+Emil+Du+Bois-Reymon+%5Bdi+bo%C3%A1+rejmon%2C+vlevo%5D+%E2%80%93+vyvinul+techniku+stimulace+a+registrace+pomoc%C3%AD+galvanometru+vlastn%C3%AD+v%C3%BDroby+%285+km+dr%C3%A1tu%2C+24+tis%C3%ADc+ot%C3%A1%C4%8Dek%29%3B+demonstroval+klidov%C3%BD+membr%C3%A1nov%C3%BD+potenci%C3%A1l+a+ak%C4%8Dn%C3%AD+potenci%C3%A1l+ve+svalu.jpg "1868 – Hermann von Helmholtz – změřil rychlost šíření akčního potenciálu v sedacím nervu žáby (cca 30 m/s) vs – m/s - 1872 (1873) – Gabriel Lippmann – kapilární elektrometr a rtuťový galvanometr, jemnější studie (tenká skleněná trubička naplněná rtutí převrstvenou kyselinou sírovou; meniskus rtuti se pohybuje v závislosti na elektrickém proudu), NC za fyziku N.E. Vvedensky – užil zrcátkový galvanometr měřící proudy až v rozsahu pA, malý pohyb cívky galvanometru přenášel na vzdálené stínítko, pohled se zvětšil; studoval aktivitu lidských svalů (telefon pro poslech AP, 1884) 1902 – Julius Bernstein – biopotenciály nervových a svalových buněk existují díky membráně, která je selektivně propustná pro draslík, kvantitativně odpovídající Nernstově vztahu (tehdy mluvil ještě o hypotetické membráně) – v tom se ale mýlil – Ernest Overton – svaly ztrácí dráždivost v roztocích bez sodných iontů, hypotéza výměny intracelulárního draslíku za extracelulární sodík (měnil složení ECS) 1910 – J. Bernstein a L. German popsali nervový impuls jako „putující potenciál aktivity")
13
Bernstein: záznam eleketrické aktivity a její rychlosti:
(a) „diferenciální rheotom“ sloužící ke stimulaci nervu a nahrávání odpovědi v různých intervalech po aplikaci stimulu (b) první publikovaný záznam časové konstanty akčního proudu v nervu (“negative variation”). Maximum elektrické aktivity zde představuje negativní výchylka, základní čára je klidový membránový potenciál (c) rozložení amplitud AP na nervovém vlákně v čase
 „diferenciální rheotom sloužící ke stimulaci nervu a nahrávání odpovědi v různých intervalech po aplikaci stimulu (b) první publikovaný záznam časové konstanty akčního proudu v nervu ( negative variation ). Maximum elektrické aktivity zde představuje negativní výchylka, základní čára je klidový membránový potenciál (c) rozložení amplitud AP na nervovém vlákně v čase.")
14
1936 – William Grey Walter – lokalizace nádorů v mozku pomocí EEG
1903 (1901 a dál); – Willem Einthoven – první záznamy EKG, strunný galvanometr vlastní konstrukce, určil vztahy mezi končetinovými svody, r obdržel NC za popis vztahu záznam EKG – poruchy srdeční činnosti) 1922 – Erlanger + Gassser (jeho žák) – katodový osciloskop, elektronkový zesilovač vlastní Gasserovy konstrukce, elektrofyziologicky rozlišili různé typy nervových vláken (elektroneurogram; oba za to dostali NC) (1924) 1929 – Hans Berger – použil strunný galvanometr k objevu EEG, pracoval nejdříve na obnažených mozcích, pak stříbrné drátky pod kůží; jako první popsal alfa (Bergerovy) vlny 1935 – Gibbs, Davis a Lennox – objevili epileptické rytmy v EEG (grand mal lze detekovat na EEG ještě předtím, než se plně rozvine) 1936 – William Grey Walter – lokalizace nádorů v mozku pomocí EEG První záznam EEG Bergerem z roku 1929
; – Willem Einthoven – první záznamy EKG, strunný galvanometr vlastní konstrukce, určil vztahy mezi končetinovými svody, r obdržel NC za popis vztahu záznam EKG – poruchy srdeční činnosti) 1922 – Erlanger + Gassser (jeho žák) – katodový osciloskop, elektronkový zesilovač vlastní Gasserovy konstrukce, elektrofyziologicky rozlišili různé typy nervových vláken (elektroneurogram; oba za to dostali NC) (1924) 1929 – Hans Berger – použil strunný galvanometr k objevu EEG, pracoval nejdříve na obnažených mozcích, pak stříbrné drátky pod kůží; jako první popsal alfa (Bergerovy) vlny – Gibbs, Davis a Lennox – objevili epileptické rytmy v EEG (grand mal lze detekovat na EEG ještě předtím, než se plně rozvine) 1936 – William Grey Walter – lokalizace nádorů v mozku pomocí EEG. První záznam EEG Bergerem z roku")
15
Anglická škola Keith Lucas ( ) a Edgard Douglas Adrian ( ) „The Conduction of the Nervous Impulse“, 1917 První potvrzeni regenerace akčního potenciálu během jeho cesty vláknem První potvrzení charakteristiky „všechno nebo nic“ Farmakologická blokáda přenosu vzruchu alkoholovými výpary E.D.A.

16
1939 – Alan Lloyd Hodgkin a Andrew Fielding Huxley – měření akčního potenciálu pomocí longitudinálních mikroelektrod zasunutých v obřích vláknech hlavonožců, NC 1963 1939 až začátek 50. let – John Carew Eccles (NC dtto) a O´Connor; Schaefer [šefr] a Haas (druhá skupina) – změřili synaptické potenciály a projevy působení neuropřenašečů, zejména acetylcholinu (spolu s Bernardem Katzem) 1949 – Gilbert Ling a Ralph Waldo Gerard – skleněné mikroelektrody k měření transmembránových intracelulárních záznamů, zjistili, že při akčním potenciálu dochází ke zvratu polarity (vyvrátili Bersnteinovu představu o dominanci draselných iontů – draslík sám by nikdy MP „nedotáhl ke kladným hodnotám) 1952 – Hodgkin, Huxley a Bernard Katz – nahrazení Bernsteinovy představy iontovou membránovou teorií Sir Hodgkin, 1963 Sir Huxley na Trinity College, Cambridge, 2005 Sir Eccles
 a O´Connor; Schaefer [šefr] a Haas (druhá skupina) – změřili synaptické potenciály a projevy působení neuropřenašečů, zejména acetylcholinu (spolu s Bernardem Katzem) 1949 – Gilbert Ling a Ralph Waldo Gerard – skleněné mikroelektrody k měření transmembránových intracelulárních záznamů, zjistili, že při akčním potenciálu dochází ke zvratu polarity (vyvrátili Bersnteinovu představu o dominanci draselných iontů – draslík sám by nikdy MP „nedotáhl ke kladným hodnotám) 1952 – Hodgkin, Huxley a Bernard Katz – nahrazení Bernsteinovy představy iontovou membránovou teorií. Sir Hodgkin, Sir Huxley na Trinity College, Cambridge, Sir Eccles.")
17
První opublikovaný záznam akčního ponenciálu:
A.L. Hodgkin, A.F. Huxley Action potentials recorded from inside a nerve fibre Nature, 14 (1939), pp. 710–711
, pp. 710–711.")
18
1981 – Erwin Neher a Bert Sackmann – technika patch clamp
70. léta a ta okolo – prof. Vyskočil, prof. Bureš – iontově selektivní mikroelektrody, šířící se kaliová deprese v mozkové kůře; objev nekvantového výlevu, sovětská elektrofyziologická škola 1981 – Erwin Neher a Bert Sackmann – technika patch clamp - techniku patch clamp (terčíkového zámku) vyvíjeli v letech , NC 1991 - slouží k registraci proudů 0,5 nA – 5 pA Erwin Neher Bert Sackmann příklad záznamu aktivity jednoho acetylcholinového receptoru technikou patch clamp. přelom milénia – ohromná exploze charakterizování subtypů jednotlivých iontových kanálů, tvorba chimér a cílených mutací, molekulární vizitky nesmírného množství struktur s bioelektrickými jevy spojenými, aplikace – buněčné sensory aj. (BERA –bioelectric recognition assay), ale taky….
 vyvíjeli v letech , NC slouží k registraci proudů 0,5 nA – 5 pA. Erwin Neher. Bert Sackmann. příklad záznamu aktivity jednoho acetylcholinového receptoru technikou patch clamp. přelom milénia – ohromná exploze charakterizování subtypů jednotlivých iontových kanálů, tvorba chimér a cílených mutací, molekulární vizitky nesmírného množství struktur s bioelektrickými jevy spojenými, aplikace – buněčné sensory aj. (BERA –bioelectric recognition assay), ale taky….")
19
??? Are men like waffles – are women like spaghetti ???
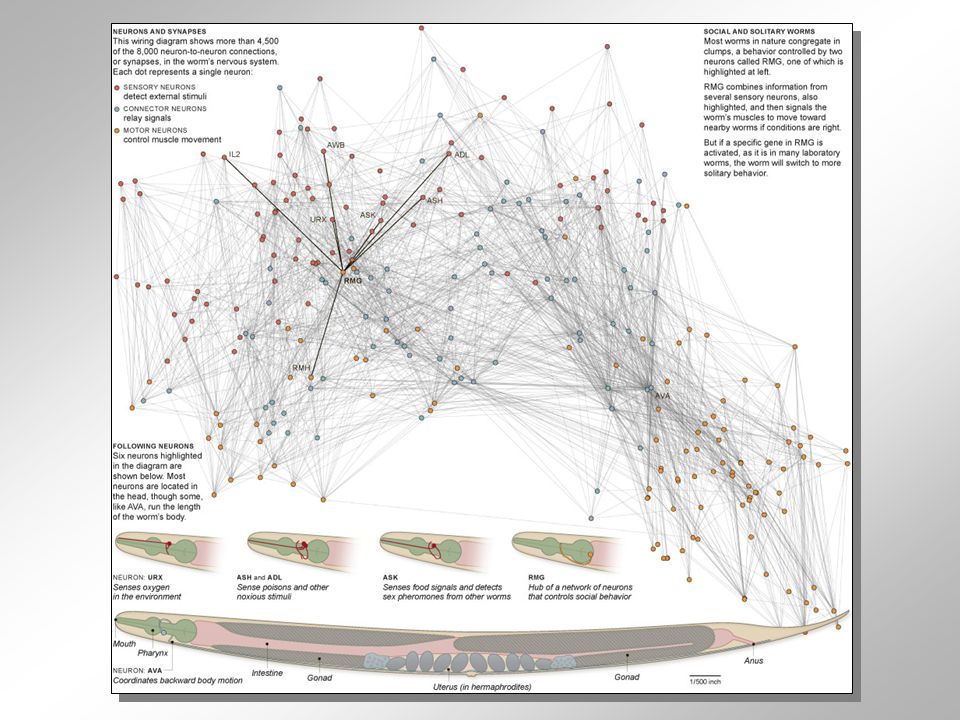
KONEKTOMIKA: decoding the essence of being – understanding the brain and all its connections Konektom = mapa všech neuronálních spojení /v mozku, sítnici…./ : Dr. Olaf Sporns (Indiana Universit, Dr. Patric Hagmann (Lausanne) C. elegans: 302 neuronů, 8000 synapsí White et. al., Philosophical Transactions of the Royal Society B: Biological Sciences 1986, 314 (1165): 1–340. ??? Are men like waffles – are women like spaghetti ??? 3D rekonstukce řezů MRI, fMRI funkční konektivita difúzní traktografie (DTI): kortikospinální trakt – sledování cesty motorické informace z motorickho kortexu přes páteřní míchu k periferním nervům CITIZEN (NEURO)SCIENCE:
 C. elegans: 302 neuronů, 8000 synapsí White et. al., Philosophical Transactions of the Royal Society B: Biological Sciences 1986, 314 (1165): 1–340. Are men like waffles – are women like spaghetti 3D rekonstukce řezů. MRI, fMRI. funkční konektivita. difúzní traktografie (DTI): kortikospinální trakt – sledování cesty. motorické informace z motorickho kortexu přes páteřní míchu k periferním nervům. CITIZEN (NEURO)SCIENCE:")
20
??? Are men like waffles – are women like spaghetti ???
KONEKTOMIKA: decoding the essence of being – understanding the brain and all its connections Konektom = mapa všech neuronálních spojení /v mozku, sítnici…./ : Dr. Olaf Sporns (Indiana Universit), Dr. Patric Hagmann (Lausanne) C. elegans: 302 neuronů, 8000 synapsí White et. al., Philosophical Transactions of the Royal Society B: Biological Sciences 1986, 314 (1165): 1–340. ??? Are men like waffles – are women like spaghetti ??? 3D rekonstukce řezů MRI, fMRI funkční konektivita difúzní traktografie (DTI): kortikospinální trakt – sledování cesty motorické informace z motorickho kortexu přes páteřní míchu k periferním nervům CITIZEN (NEURO)SCIENCE:
, Dr. Patric Hagmann (Lausanne) C. elegans: 302 neuronů, 8000 synapsí White et. al., Philosophical Transactions of the Royal Society B: Biological Sciences 1986, 314 (1165): 1–340. Are men like waffles – are women like spaghetti 3D rekonstukce řezů. MRI, fMRI. funkční konektivita. difúzní traktografie (DTI): kortikospinální trakt – sledování cesty. motorické informace z motorickho kortexu přes páteřní míchu k periferním nervům. CITIZEN (NEURO)SCIENCE:")
22
10 out of 950 neurons reconstructed in a block of mouse retina, imaged using serial block-face electron microscopy (gray images). Spheres indicate cell bodies (red, ganglion cells, green, amacrine cells, blue, bipolar cells).© Fabian Isensee, Julia Kuhl; Helmstaedter et al., 2013; © Max Planck Institute for Medical Research, Heidelberg, Germany
.© Fabian Isensee, Julia Kuhl; Helmstaedter et al., 2013; © Max Planck Institute for Medical Research, Heidelberg, Germany.")
23
Credit: http://www. datanami

24
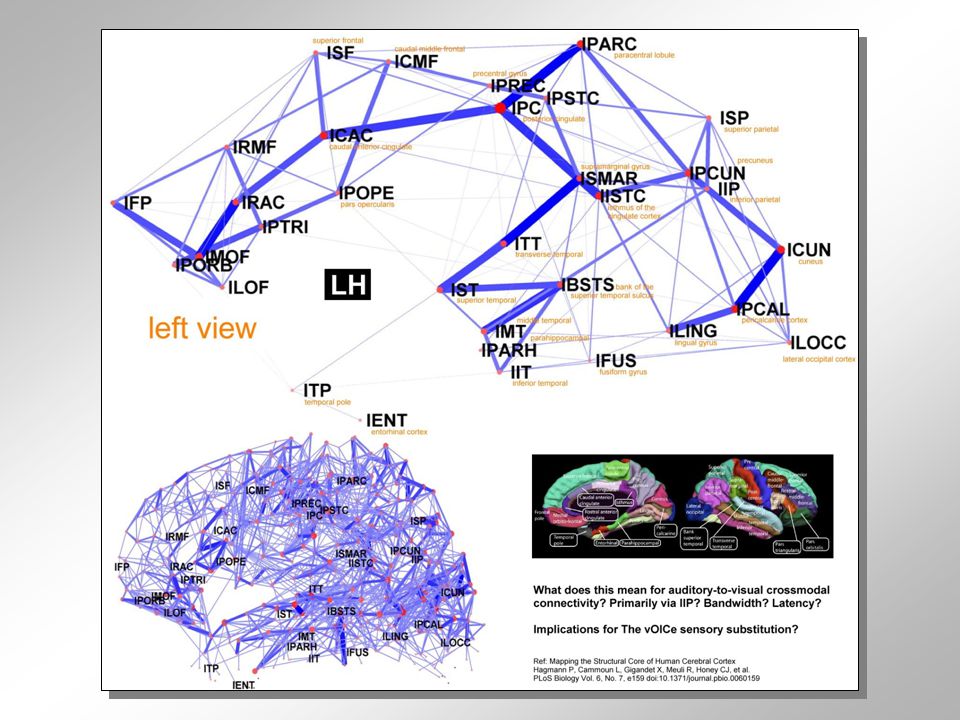
Figure 1: The human connectome
Figure 1: The human connectome. Images show the fiber architecture of the human brain as revealed by diffusion imaging (left), a reconstructed structural brain network (middle) and the location of the brain's core, its most highly and densely interconnected hub (right). Image on the left courtesy of Patric Hagmann, University of Lausanne; middle and right panels are from Hagmann et al. (2008).
, a reconstructed structural brain network (middle) and the location of the brain s core, its most highly and densely interconnected hub (right). Image on the left courtesy of Patric Hagmann, University of Lausanne; middle and right panels are from Hagmann et al. (2008).")
25
"DTI-sagittal-fibers" by Thomas Schultz - Own work
"DTI-sagittal-fibers" by Thomas Schultz - Own work. Licensed under CC BY-SA 3.0 via Wikimedia Commons -

26
Progression of seed-induced Aβ deposition within the limbic connectome.
Brain Pathol Feb 9; Ye L, Hamaguchi T, Fritschi SK, Eisele YS, Obermüller U, Jucker M, Walker LC Abstract An important early event in the pathogenesis of Alzheimer's disease (AD) is the aberrant polymerization and extracellular accumulation of amyloid-β peptide (Aβ). In young transgenic mice expressing the human Aβ-precursor protein (APP), deposits of Aβ can be induced by the inoculation of minute amounts of brain extract containing Aβ aggregates ("Aβ seeds"), indicative of a prion-like seeding phenomenon. Moreover, focal intracerebral injection of Aβ seeds can induce deposits not only in the immediate vicinity of the injection site, but, with time, also in distal regions of the brain. However, it remains uncertain whether the spatial progression of Aβ deposits occurs via nonsystematic diffusion from the injection site to proximal regions or via directed transit along neuroanatomical pathways. To address this question, we analyzed the spatiotemporal emergence of Aβ deposits in two different APP-transgenic mouse models that had been previously inoculated with Aβ seeds into the hippocampal formation. The results revealed a specific, neuroanatomically constrained pattern of induced Aβ deposits in structures corresponding to the limbic connectome, supporting the hypothesis that neuronal pathways act as conduits for the movement of proteopathic agents among brain regions, thereby facilitating the progression of disease.
 is the aberrant polymerization and extracellular accumulation of amyloid-β peptide (Aβ). In young transgenic mice expressing the human Aβ-precursor protein (APP), deposits of Aβ can be induced by the inoculation of minute amounts of brain extract containing Aβ aggregates ( Aβ seeds ), indicative of a prion-like seeding phenomenon. Moreover, focal intracerebral injection of Aβ seeds can induce deposits not only in the immediate vicinity of the injection site, but, with time, also in distal regions of the brain. However, it remains uncertain whether the spatial progression of Aβ deposits occurs via nonsystematic diffusion from the injection site to proximal regions or via directed transit along neuroanatomical pathways. To address this question, we analyzed the spatiotemporal emergence of Aβ deposits in two different APP-transgenic mouse models that had been previously inoculated with Aβ seeds into the hippocampal formation. The results revealed a specific, neuroanatomically constrained pattern of induced Aβ deposits in structures corresponding to the limbic connectome, supporting the hypothesis that neuronal pathways act as conduits for the movement of proteopathic agents among brain regions, thereby facilitating the progression of disease.")
28
Opakování za domácí úkol

29
Elektrostatické pole Název elektřina , resp. elektrikum, zavedl Wiiliam Gilbert, fyzik královny Alžběty, roku 1600, podle řeckého lektron, tedy jantar. Elektrika = předměty, které třeny kožešinou přitahovaly malé kousky papíru nebo bezové duše. Sklo produkuje kladný typ fluida, záporné fluidum produkuje např. jantar (fluidum pryskyřičné; Benjamin Franklin, 1747). ,, Elektrické jevy mají původ v elektrickém náboji. „Co je to elektrický náboj nebo elektřina, neumíme říci, je to jednoduše jedna ze základních věcí vesmíru.“ Z. Vodrážka, Fyzikální chemie pro biologické vědy, s. 20. Mezi základními elementárními částicemi hmoty jsou dvě s diskrétním, stejným, opačným elementárním nábojem. Jsou to proton a elektron. symbol náboj hmotnost v g proton elektron p e 1 pozitivní element. náboj 1, C 1 negativní element. náboj 1, C 1, g 9, g Jednotkou náboje je coulomb (C); 1 C = 1 A . s El. náboje (Q) kolem sebe vytvářejí elektrostatické pole a působí na sebe elektrostatickými silami. El. náboj (Q) se definuje pomocí síly (F), kterou působí na jiné náboje stejné velikosti, nebo pomocí jejich vzájemné potenciální energie (Ep).
. ,, Elektrické jevy mají původ v elektrickém náboji. „Co je to elektrický náboj nebo elektřina, neumíme říci, je to jednoduše jedna ze základních věcí vesmíru. Z. Vodrážka, Fyzikální chemie pro biologické vědy, s. 20. Mezi základními elementárními částicemi hmoty jsou dvě s diskrétním, stejným, opačným elementárním nábojem. Jsou to proton a elektron. symbol. náboj. hmotnost v g. proton. elektron. p. e. 1 pozitivní element. náboj 1, C. 1 negativní element. náboj 1, C. 1, g. 9, g. Jednotkou náboje je coulomb (C); 1 C = 1 A . s. El. náboje (Q) kolem sebe vytvářejí elektrostatické pole a působí na sebe elektrostatickými silami. El. náboj (Q) se definuje pomocí síly (F), kterou působí na jiné náboje stejné velikosti, nebo pomocí jejich vzájemné potenciální energie (Ep).")
30
Elektrostatické pole lze popsat intenzitou el. pole E a
+ El. náboj (Q) se definuje pomocí síly (F), kterou působí na jiné náboje stejné velikosti, nebo pomocí jejich vzájemné potenciální energie (Ep). Stejné náboje se odpuzují, opačné přitahují. QA QB r2 F = k Coulombův zákon k = 1 / 4pe - Pokud jsou náboje QA a QB, působící na sebe silou F, od sebe v určité vzdálenosti r, bude potenciální energie náboje QA definována jako změna potenciální energie spojená s přenosem tohoto náboje QA z nekonečna (kde Ep = 0) do vzdálenosti r. QA QB r Ep = k Elektrické pole izolovaných bodových nábojů. Plné čáry = siločáry; přerušované čáry= ekvipotenciály. Při přibližování se opačně nabitých částic se Ep uvolňuje a systém má negativní potenciální energii. Při přibližování stejně nabitých částic musí systém energii přijímat, jeho celková Ep je tedy kladná. potenciální energie Ep + x2 x1 E2 E1 Elektrostatické pole lze popsat intenzitou el. pole E a elektrickým potenciálem f.
 se definuje pomocí síly (F), kterou působí na jiné náboje stejné velikosti, nebo pomocí jejich vzájemné potenciální energie (Ep). Stejné náboje se odpuzují, opačné přitahují. QA QB. r2. F = k. Coulombův zákon. k = 1 / 4pe. - Pokud jsou náboje QA a QB, působící na sebe silou F, od sebe v určité vzdálenosti r, bude potenciální energie náboje QA definována jako změna potenciální energie spojená s přenosem tohoto náboje QA z nekonečna (kde Ep = 0) do vzdálenosti r. QA QB. r. Ep = k. Elektrické pole izolovaných bodových nábojů. Plné čáry = siločáry; přerušované čáry= ekvipotenciály. Při přibližování se opačně nabitých částic se Ep uvolňuje a systém má negativní potenciální energii. Při přibližování stejně nabitých částic musí systém energii přijímat, jeho celková Ep je tedy kladná. potenciální energie Ep. + x2. x1. E2. E1. Elektrostatické pole lze popsat. intenzitou el. pole E a. elektrickým potenciálem f.")
31
Intenzita el. pole E je dána silou, kterou by v daném bodě pole působilo na jednotkový náboj Q´ (náboj 1 C): E = Q´ F [E] = N / C Při pohybu náboje v el. poli je nutné překonávat určitou sílu F = E . Q´. Při tom se vykoná určitá práce W a náboj získá určitou potenciální energii Ep. Elektrický potenciál f je definován jako podíl elektrické potenciální energie Ep kladného náboje a velikosti tohoto náboje: f = Q´ Ep [f] = J / C = V Při přenosu náboje se vykoná práce, resp. el. potenciál se číselně rovná práci, kterou je třeba vykonat při přenesení jednotkového kladného náboje po libovolné dráze z místa s nulovým el. potenciálem do daného místa. Velikost této práce je dána rozdílem potenciálních energií daného náboje v počátečním (1) a konečném místě (2): f = Q´ W Náboj o velikosti 1 C tekoucí z plus k mínus pólu vykoná práci 1 joule. W = EP2 – EP1 = Ep = Q´ (f2 – f1) Všechny body v el. poli, které mají stejný el. potenciál, tvoří plochu = ekvipotenciálu. Rozdíl dvou potenciálů (f2 – f1) označujeme jako elektrické napětí U : U = Q´ W W = EP2 – EP1 = Ep = Q´ (f2 – f1) Q´. U Ep = Q´. U
 a konečném místě (2): f = Q´ W. Náboj o velikosti 1 C tekoucí z plus k mínus pólu vykoná práci 1 joule. W = EP2 – EP1 = Ep = Q´ (f2 – f1) Všechny body v el. poli, které mají stejný el. potenciál, tvoří plochu = ekvipotenciálu. Rozdíl dvou potenciálů (f2 – f1) označujeme jako elektrické napětí U : U = Q´ W. W = EP2 – EP1 = Ep = Q´ (f2 – f1) Q´. U Ep = Q´. U.")
32
či pomocí vektorového zápisu
+ - + f2 f1 E U U = Q´ W U = f2 – f1 Ep = Q´. U Při pohybu náboje po ekvipotenciální ploše je napětí U = 0. Nemění se tedy ani potenciální energie náboje a elektrické síly nekonají práci. Vektor intenzity el. pole je vždy kolmý na ekvipotenciální plochu. Elektrické pole izolovaných bodových nábojů. Plné čáry = siločáry; přerušované čáry= ekvipotenciály. Homogenní elektrické pole. Plné čáry = siločáry; přerušované čáry= ekvipotenciály. V každém bodě prostoru je intenzita el. pole vyjádřena jediným vektorem. Propojením všech na sebe navazujících vektorů intenzity el. pole bychom dostali obraz siločar daného pole. Tam, kde jsou siločáry hustě u sebe, je intenzita pole větší než tam, kde jsou řídce. El. pole má tedy v různých místech různou intenzitu. Tu můžeme vyjádřit také jako gradient potenciálu f: Ex = x Ey = y Ez = z či pomocí vektorového zápisu E = - grad f Záporné znaménko naznačuje, že náboje se budou pohybovat od míst s vyšším potenciálem do míst s nižším potenciálem.

33
Ze změny potenciálu lze vyčíst, kolik iontů se kam přesune.
Absolutní potenciál bodového náboje ve vzdálenosti r od náboje: V = 4peor Q permitivita vakua eo = 8, F.m-1 Příklad: V jaké vzdálenosti od náboje 1 mC naměříme hodnotu potenciálu 1 V? [cca 9 km] Ze změny potenciálu lze vyčíst, kolik iontů se kam přesune. 1 C představuje asi 6, jednotkových, elementárních nábojů (1/e). Fradayova konstanta je náboj 1 gramionu elementárních nábojů: je to součin Avogadrovy konstanty a elementárního náboje: F = NA . e = 6, x 1, = C.mol-1 V elektrickém poli o intenzitě E se kladné částice dají do pohybu v kladném směru intenzity pole, záporné v opačném směru. Uspořádaný pohyb náboje se nazývá elektrický proud. El. proud značíme I; je to základní veličina soustavy SI, jeho jednotkou je ampér . 1 A představuje množství náboje 1 C, který projde vodičem za 1 s: I = t Q [I] = A Mezi konci tohoto vodiče můžeme naměřit napětí U. Vztah mezi proudem a napětím pak popisuje Ohmův zákon: U = I . R R je odpor vodiče. Čím větší je odpor vodiče, tím větší napětí musí být na jeho koncích, aby jím protékal proud I – a naopak.
. Fradayova konstanta je náboj 1 gramionu elementárních nábojů: je to součin Avogadrovy konstanty a elementárního náboje: F = NA . e = 6, x 1, = C.mol-1. V elektrickém poli o intenzitě E se kladné částice dají do pohybu v kladném směru intenzity pole, záporné v opačném směru. Uspořádaný pohyb náboje se nazývá elektrický proud. El. proud značíme I; je to základní veličina soustavy SI, jeho jednotkou je ampér . 1 A představuje množství náboje 1 C, který projde vodičem za 1 s: I = t. Q. [I] = A. Mezi konci tohoto vodiče můžeme naměřit napětí U. Vztah mezi proudem a napětím pak popisuje Ohmův zákon: U = I . R. R je odpor vodiče. Čím větší je odpor vodiče, tím větší napětí musí být na jeho koncích, aby jím protékal proud I – a naopak.")
34
A) Kanály v membráně a membrána samotná jako vodiče
U Pro velikost odporu vodič plyne z Ohmova zákona: [R] = = V . A-1 Jednotkou vodivosti G (g z guide, vést) je siemens S: (vodivost je převrácená hodnota odporu) G = R 1 [G] = -1 = S Nervové vlákno je vodič a lze jej tedy jako vodič / elektrický obvod schematizovat. I1 I2 I U + - I1 = R1 U G1 G2 g1 g2 U Zapojení vodičů vedle sebe = paralelně; paralelně zapojené vodiče se chovají jako jeden vodič. Tímto způsobem lze vyjádřit i elektrické chování membrány samotné. I2 = R2 U I = I1 + I2 = R1 U + R2 A) Kanály v membráně a membrána samotná jako vodiče R2 1 I = U ( ) R1 R2 1 R1 R = + 1. Kirchhoffův zákon (součet proudů vstupujících do uzlu = součtu proudů vystupujících z uzlu) G = G1 + G2
 je siemens S: (vodivost je převrácená hodnota odporu) G = R. 1. [G] = -1 = S. Nervové vlákno je vodič a lze jej tedy jako vodič / elektrický obvod schematizovat. I1. I2. I. U. + - I1 = R1. U. G1. G2. g1. g2. U. Zapojení vodičů vedle sebe = paralelně; paralelně zapojené vodiče se chovají jako jeden vodič. Tímto způsobem lze vyjádřit i elektrické chování membrány samotné. I2 = R2. U. I = I1 + I2. = R1. U. + R2. A) Kanály v membráně a membrána samotná jako vodiče. R2. 1. I = U ( + ) R1. R2. 1. R1. R. = + 1. Kirchhoffův zákon (součet proudů vstupujících do uzlu = součtu proudů vystupujících z uzlu) G = G1 + G2.")
35
B) Axoplasma (cytoplasma nervového vlákna) jako vodič U = I . R
+ - U1 U2 U3 U1 = I . R1 U2 = I . R2 U3 = I . R3 Zapojení vodičů za sebou = sériově. Proud se nikde nemůže hromadit, je stejný při vstupu i výstupu do/z každého vodiče. Napětí na jednotlivých vodičích je tedy úměrné jejich odporu. U = I . (R1 + R2 + R3) Odpory sériově zapojených vodičů se sčítají. R = R1 + R2 + R3 C) Celkové schéma nervového vlákna jako vodiče rm ri U Ekvivalentní elektrický obvod představující nervové vlákno (zatím ignorujeme tzv. kapacitu membrány). rm... odpor membrány (1.103 cm2) ri... podélný odpor axoplasmy (30 cm) Membrána a všechny struktury v ní lokalizované se pokrývají nábojem. Pokud jsou různé vodiče (membrána, kanály, přenašeče...) pokryty stejným nábojem, není jejich elektrický potenciál obecně stejný. Potenciál vodiče se bude měnit podle jeho tvaru, podle jiných těles, které jsou v jeho přítomnosti, a pod. Všechny tyto vlivy shrnuje veličina nazvaná kapacita vodiče.
 Odpory sériově zapojených vodičů se sčítají. R = R1 + R2 + R3. C) Celkové schéma nervového vlákna jako vodiče. rm. ri. U. Ekvivalentní elektrický obvod představující nervové vlákno (zatím ignorujeme tzv. kapacitu membrány). rm... odpor membrány (1.103 cm2) ri... podélný odpor axoplasmy (30 cm) Membrána a všechny struktury v ní lokalizované se pokrývají nábojem. Pokud jsou různé vodiče (membrána, kanály, přenašeče...) pokryty stejným nábojem, není jejich elektrický potenciál obecně stejný. Potenciál vodiče se bude měnit podle jeho tvaru, podle jiných těles, které jsou v jeho přítomnosti, a pod. Všechny tyto vlivy shrnuje veličina nazvaná kapacita vodiče.")
36
Membrána je velmi tenké a velmi kvalitní dielektrikum.
Kapacita membrány Membrána buňky odděluje od sebe nevodivě dvě prostředí s různou koncentrací iontů. Část iontů – náboje z okolního prostředí se rozprostře i po jejím povrchu: membrána se nabíjí. Nevodivé látky obklopené z obou stran vodiči slouží jako kondenzátory. V tomto případě jsou vodiči vodné roztoky z obou stran membrány a izolačním materiálem, dielektrikem, je sama membrána. Vodiče se nabíjejí stejně velkými, ale opačnými náboji +Q a –Q. Tím mezi nimi vznikne napětí U. Kapacita membrány se definuje jako podíl kladného náboje a tohoto napětí: C = U Q [C] = C . V-1 = F (farad) Farad je dost velká jednotka, v praxi mikrofarady – pikofarady. Typická kapacita membrány neuronu je 1 mF na cm2. Modelová buňka o průměru 25 mm a povrchu cm2 (8 000 mm2) má tedy celkovou kapacitu membrány F, tj. 80 pF. Při membránovém potenciálu -70 mV tedy naaukumuluje na svém povrchu C náboje na cm2. Při daném povrchu je to C náboje, což odpovídá 3,5.107 kusů elementárního náboje (např. záporných iontů na vnitřní straně membrány). Membrána je velmi tenké a velmi kvalitní dielektrikum. + =
 Farad je dost velká jednotka, v praxi mikrofarady – pikofarady. Typická kapacita membrány neuronu je 1 mF na cm2. Modelová buňka o průměru 25 mm a povrchu cm2 (8 000 mm2) má tedy celkovou kapacitu membrány F, tj. 80 pF. Při membránovém potenciálu -70 mV tedy naaukumuluje na svém povrchu C náboje na cm2. Při daném povrchu je to C náboje, což odpovídá 3,5.107 kusů elementárního náboje (např. záporných iontů na vnitřní straně membrány). Membrána je velmi tenké a velmi kvalitní dielektrikum. + =")
37
Membrána je velmi tenké a –přinejmenším její lipidická složka- velmi
Membrána je velmi tenké a –přinejmenším její lipidická složka- velmi kvalitní dielektrikum. Jde vlastně o soustavu paralelně zapojených kondenzátorů, které dohromady považujeme za jeden kondenzátor. Při spojení kondenzátorů s kapacitou C1, C2 ... Cn se jejich kapacity sčítají. + = C1 C2 R1 R2 + = Membrána je ale také baterie paralelních odporů. Celkově tedy náhradní schéma membrány vypadá takto: RC člen rm ri U cm Ekvivalentní elektrický obvod představující nervové vlákno se zahrnutou kapacitou membrány. cm... kapacita membrány (1 mF na cm2) rm... odpor membrány (1.103 cm2) ri... podélný odpor axoplasmy (30 cm) Zatímco u obvodů ovlivněných čistě Ohmovým zákonem je napětí úměrné proudu (U = I.R), u obvodů s kapacitou se musí nejdříve nabít kondenzátor, pak jde proud i k odporu. Postupně s nabíjením kondenzátoru roste i napětí – napětí je v těchto obvodech funkcí času. Na konečnou hodnotu napětí roste exponenciálně s časovou konstantou t = R.C.
 rm... odpor membrány (1.103 cm2) ri... podélný odpor axoplasmy (30 cm) Zatímco u obvodů ovlivněných čistě Ohmovým zákonem je napětí úměrné proudu (U = I.R), u obvodů s kapacitou se musí nejdříve nabít kondenzátor, pak jde proud i k odporu. Postupně s nabíjením kondenzátoru roste i napětí – napětí je v těchto obvodech funkcí času. Na konečnou hodnotu napětí roste exponenciálně s časovou konstantou t = R.C.")
38
Postupně s nabíjením kondenzátoru roste i napětí – napětí je v těchto obvodech funkcí času. Na konečnou hodnotu napětí roste exponenciálně s časovou konstantou t = R.C. U (t) = I. R (1 – e –t/t) U Ir Ic U = Q / C dU / dt = I / C U = I. R Časová konstanta t udává dobu, za kterou potenciál dosáhne 63% své výsledné hodnoty (1 – 1/e). t = R.C (1) I(t) = Ir + Ic I(t) = Um Cm dt 1 rm dUm . Ir = 1 rm Um (2) (5) Ic = = dQ dt d(CmUm) (3) V = I(t) . Rm (1 – e ) -t / (6) Ic = Cm . dUm dt (4) (7) = rm. Cm
 = I. R (1 – e –t/t) U. Ir. Ic. U = Q / C. dU / dt = I / C. U = I. R. Časová konstanta t udává dobu, za kterou potenciál dosáhne 63% své výsledné hodnoty (1 – 1/e). t = R.C. (1) I(t) = Ir + Ic. I(t) = Um. Cm. dt. 1. rm. dUm. . Ir = 1. rm. Um. (2) (5) Ic = = dQ. dt. d(CmUm) (3) V = I(t) . Rm (1 – e ) -t / (6) Ic = Cm. . dUm. dt. (4) (7) = rm. Cm.")
39
Co si pamatovat z dnešní přednášky
co je základem bioelektrických jevů elementární náboj Faradayova konstanta Coulombův a Ohmův zákon vztahy pro napětí, proud, kapacitu chování membrány jako odporu chování membrány jako kondenzátoru časová konstanta

Podobné prezentace