Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Motorická kontrola

2
Sval - efektor motorického systému

3
A – srdeční – autonomní NS B – hladká – autonomní NS
C B Svalovina: A – srdeční – autonomní NS B – hladká – autonomní NS C – příčně pruhovaná Příčné pruhování - rozdíly v lomivém indexu , světlý proužek I (aktin tenčí) s čárou Z uprostřed, tmavý proužek A (myosin, tlustčí) s H úsekem uprostřed, Sarkomera – oblast mezi sousedními čarami Z, Různé kontraktilní vlastnosti, které předurčují odpověd svalu na podráždění
 s čárou Z uprostřed, tmavý proužek A (myosin, tlustčí) s H úsekem uprostřed, Sarkomera – oblast mezi sousedními čarami Z, Různé kontraktilní vlastnosti, které předurčují odpověd svalu na podráždění.")
4
Srdeční svalovina - morfologie
Příčné pruhování podobné jako u kosterní svaloviny Svalová vlákna se proplétají a větví V místech přechodu membrány vláken zvlněny Tvoří interkalární disky – oblasti s těsnými spoji - syncytium Snadný přenos Tahu Podráždění

5
Elektrické spoje

6
Srdeční svalovina – elektrické vlastnosti
Myogenní – stahuje se i v nepřítomnosti motorického vstupu Pacemakerový potenciál Nestabilní klidový membránový potenciál Pomalé snižování propustnosti pro K+ ionty Strmost určuje frekvenci rytmicity 2 specializovaná místa myogenní aktivity Síňokomorový (AV) a Síňový uzlík (SA) rytmicita SA>AV>svaloviny SA udává rytmus – vlastní pacemaker pro celé srdce
 a. Síňový uzlík (SA) rytmicita SA>AV>svaloviny. SA udává rytmus – vlastní pacemaker pro celé srdce.")
8
Srdeční svalovina – elektrické vlastnosti
AP Rychlá depolarizace - 2 ms, přestřelení, Na+ kanály Pomalá repolarizace – 300 msec, pomalé Na+ kanály, K+ kanály Dlouhá refrakterní perioda - kontrakce se nemohou sumovat Kontrakce Vše nebo nic 1,5 x delší než AP Regulace - autonomní Parasympatikus, vagus, Ach Sympatikus, adrenalin, b-adrenergní receptory

9
Ach receptory srdeční svaloviny
1. Snižuje velikost napěťově řízeného Ca2+ proudu prostřednictvím vnitrobuněčného mediátoru 2. Prostřednictvím G proteinu přímo otevírá K+ kanály Pomalejší depolarizace a Prodloužená hyperpolarizace

10
b adrenergní receptory srdeční svaloviny
Prostřednictvím různých G proteinů zvyšuje depolarizaci a zrychlení srdeční aktivity

11
Hladká svalovina - morfologie
Zřetelně odlišná od srdeční a kosterní svaloviny – dělení: Útrobní hladká Extenzivní elektrické propojení Velké oblasti svaloviny se kontrahují společně syncytium – střevo, děloha, močovody Vícejednotková hladká – Buňky nepropojeny – zornička, artérie, kůže savcůkaždý sval inervován několika autonomními motorickými neurony Málo elektrických spojů mezi buňkami

12
Elektrické spoje Ztluštěniny podél autonomního nervu

13
Hladká svalovina – elektrické vlastnosti
Nestabilní klidový membránový potenciál Na membránový potenciál nasedají vlny různých typů Některé přestřelují linii nulového potenckálu, některé ne Pacemakerový potenciál – ohniska se přesouvají Spřažení dráždivosti a staživosti pomalé Stálé nepravidelné kontrakce nezávislé na neuronální aktivitě Kontrakce i po předchozím protažení Natažení způsobuje pokles membránového potenciálu

14
Funkce inervace Dvojí autonomní inervace V jiných orgánech modifikace
Modifikuje aktivitu Antagonistické působení Ve střevě parasympatikus zvyšuje aktivitu, sympatikus tlumí V jiných orgánech modifikace

15
Nervosvalové spojení hladké svaloviny
Hladká svalovina – nemá typické synaptické spoje Ztluštěniny axonů (2mm s 4mm inter-úseky) se synaptickými váčky Difuse mediátoru ke svalovým buňkám Autonomní systém kontroluje spíše vrstvy hladké svaloviny než jednotlivé svaly Významná místní kontrola enterickým nervovým systémem s mnoha neuromediátory
 se synaptickými váčky. Difuse mediátoru ke svalovým buňkám. Autonomní systém kontroluje spíše vrstvy hladké svaloviny než jednotlivé svaly. Významná místní kontrola enterickým nervovým systémem s mnoha neuromediátory.")
16
Příčně pruhovaný sval Vlákno samostatná vícejaderná buňka
Vznik splynutím mnoha embryonálních buněk tvořeno myofibrilami obaleno sarkolemou a sarkoplasmatickým retikulem složeno z myofibril = filamenty kontraktilních bílkovin Motorický axon Žíhání Vlákno Jádra Svalová vlákna

17
Snopce svalových vláken – napnuty paralelně mezi šlachami
Svalová vlákna obalená vazivovou pochvou Svalové vlákno Myofibrila Pojivová tkáň kolem svalu

18
Sarkoméra Myosin Aktin Stah H Proužek A Proužek I
Šíře A proužku je stálá, I proužek se při zasouvání zkracuje Stah

19
Tropomyosin V klidu jsou místa vážící myosin blokována relaxační bílkovinou komplex troponinu a tropomyosinu brání interakci aktinu s myozinem tropomyosin - připojuje troponin k aktinu Depolarizace zvyšuje volnou koncentraci Ca2+ (Ca2+ svalu v klidu vázáno v sarkoplasmatickém retikulu) Ca2+ tlumí inhibiční vliv tropomyosinu To aktivuje ATPázovou aktivitu v hlavách myosinových molekul a Energie – hydrolýza ATP na ADP, adenosintrifosfatázová aktivita se nalézá v hlavách myosinových molekul, které jsou v kontaktu s aktinem. ATP také používáno pro zpětné čerpání Ca do sarkoplasmatického retikula. Jak kontrakce, tak i relaxace svalu vyžaduje energii
 Ca2+ tlumí inhibiční vliv tropomyosinu. To aktivuje ATPázovou aktivitu v hlavách myosinových molekul a. Energie – hydrolýza ATP na ADP, adenosintrifosfatázová aktivita se nalézá v hlavách myosinových molekul, které jsou v kontaktu s aktinem. ATP také používáno pro zpětné čerpání Ca do sarkoplasmatického retikula. Jak kontrakce, tak i relaxace svalu vyžaduje energii.")
20
AP Ca2+ Ca2+ Spřažení dráždivosti a stažlivosti Aktin
Myosinové hlavy se vrací do původní polohy Aktivace stažlivého mechanismu Hlavy myosinu se váží na aktin Ca2+ Myosin Pi ATP Hydrolýza ATP Navázání ATP uvolní aktin Příčné můstky mění svůj tvar a posouvají aktin o 12 nm do středu sarkomery

21
Kosterní svalovina Nemá elektrické spoje Nevětví se
Kontrola somatickým nervovým systémem Strukturální a funkční diverzita Tonické Twich

22
Tonické svaly Fázické svaly Stah Více myoglobinu Bohatě mitochondrií
Pomalý, relativně slabý, trvání - stovky ms Více myoglobinu Bohatě mitochondrií Pro dlohodobé kontrakce bez únavy svalů Posturální svalovina Fázické svaly Stah Rychlý a silný, 5-10ms Histologická a biochemická variabilita Okohybné svaly, svalovina ruky

23
Fázické svaly obratlovců
Kategorizace podle hlavních metabolických drah Pomalé – „slow oxidative“ SO Rychlé – „fast oxidative glycolytic, FOG“ Rychlé – „Fast glykolytic“, FG Svaly obvykle obsahují směs vláken různých typů Atribut rychlé či pomalé se vztahuje ke kontraktilním vlastnostem

24
Rychlá vlákna Pomalá vlákna Diverzita svalových vláken savců

25
Charakteristiky savčích fázických svalů
Vlastnost Rychlost kontrakce Trvání stahu Únavnost Obsah glykogenu Obsah myoglobinu Obsah mitochondrií Glykolitická aktivita Oxidativní kapacita Pomalé SO Pomalá Dlouhé Nízký Vysoký Nízká Vysoká Pomalé FOG Rychlá Krátké Nízká Střední Vysoký Vysoká Rychlé FG Rychlá Krátké Vysoká Vysoký Nízký Nízká

26
Funkční organizace svaloviny
Antagonistické svaly Synergistické svaly - Extenzor Flexor Kloub Exoskelet

27
Neurální kontrola kosterní svaloviny

28
Obecně Motorický systém je složitější než smyslový Hůř se studuje
Každý pohyb je kontrolován smyslovými orgány, které zpětně modifikují aktivitu motorických drah Každý neuron motorické dráhy odpovídá na stimulaci určitým setem interneuronů předchozí úrovně, který NEZNÁME a tudíž ho nemůžeme napodobit

29
Principy motorické kontroly
Chování jednodušších živočichů založeno na jednoduchých mechanismech probíhajících na periferní úrovni Evolučně vyšší živočichové vytvářejí vyšší koordinační mechanismy v závislosti na stupni tzv. encefalizace Jednodušší koordinační mechanismy na segmentální úrovni zůstávají Ale „očekávají“ více z centra Při přerušení toku info z centra – „šok“ – spinální šok (vymizeni funkce, později objevení se nové, primitivnější – „vyvázání“ se z kontrolního vlivu

30
Motorické jednotky 1 Motorický neuron – inervuje obvykle jenom část svalových vláken – motorická jednotka Fázické svaly obratlovců – jednotky se nepřekrývají Bezobratlí a tonické svaly obratlovců – přerkývání motorických jednotek Svalová membrána není dostatečně excitabilní Aktivace jedné synapse nevyvolá stah Proto mnohočetné synapse

31
Motorické axony Motorické jednotky obratlovčího svalu se nepřekrývají

32
Kontrola svalového napětí
Nábor motorických jednotek - recruitment – Ve svalech s více motorickými jednotkami Přesná hierarchie, které MJ a kdy budou aktivovány Kontrola frekvence AP Frekvence AP určuje, jestli se sval před dalším stahem uvolní Exkluzivně u bezobratlých

33
Pořadí aktivace motoneuronů dáno velikostí
Svaly složeny s FG, FOG a SO vláken FG inervovány většími motoneurony SO inervovány menšími motoneurony Vlákna ve svalech jsou specificky kombinována pro specifické funkce Svaly jedné funkce však mohou být používány různě v závislosti na typu a pořadí aktivovaných jednotek Malé neurony bývají aktivovány dříve Motoneurony kontrolovány prostřednictvím interneuronů - Mozek „vybírá“ specifické soubory interneuornů

34
A B Snímací elektroda Snímací elektroda Aktivace Aktivace
Iniciální zóna Iniciální zóna Princip velikosti - malé neurony excitovány první

35
Fyziologické vlastnosti svalů dány inervací
Různé typy svalů inervovány motoneurony různých fyziologických vlastností Fyziologické vlastnosti motoneuronů odpovídají kontraktilním vlastnostem svalu Jestliže se během vývoje experimentálně vymění inervace, svaly se typu inervace „přizpůsobí“ - princip shody

36
Fast axon Elektroda - EPSP Princip shody Rychlé vlákno FG FOG Pomalé vlákno SO Slow axon Velké pomalé EPSP

37
Motorika Volní x mimovolní Opěrná (postojová) x cílená (pohybová) Reflexní x s centrálním motorickým programem
 x cílená (pohybová) Reflexní x s centrálním motorickým programem.")
38
Motoneurony

39
Morfologie motoneuronů
Bezobratlí Obratlovci Dendrity Tělo Dendrity Axon Axon Tělo Monopolární, těla na periferii ganglií, smyslové vstupy motorické výstupy v nervech promíchány Multipolární, těla v šedé hmotě míchy, motorické výstupy odděleny

40
Motoneurony – místo konečného výstupu CNS info ke svalům
Gama-eferentní systém Intrafuzální vlákna svalových vřetének Pod kontrolou sestupných drah Alfa-eferentní systém Extrafuzální vlákna Obratlovci – největší neurony, velké dendritické větvení (až 10 cm), velký informační přenos, axony tvoří kolaterály Zpětnovazebná kontrola prostřednictvím svalových vřetének Inervace dvojího typu, Ia aferenty-největší a nejrychlejší z perif. Nervů, dynamické parametry II – pomalejší aferenty, statická míra natažení Soubory motoneuronů určitého svalu tvoří v míše shluky – motorická jádra Somatotopické uspořádání
, velký informační přenos, axony tvoří kolaterály. Zpětnovazebná kontrola prostřednictvím svalových vřetének. Inervace dvojího typu, Ia aferenty-největší a nejrychlejší z perif. Nervů, dynamické parametry. II – pomalejší aferenty, statická míra natažení. Soubory motoneuronů určitého svalu tvoří v míše shluky – motorická jádra. Somatotopické uspořádání.")
41
Konvergence a divergence aferentních signálů
Aferenty po vstupu do míchy zadními míšními kořeny tvoří kolaterály kaudální, rostrální, ventrální Každé vlákno tvoří cca synapsí (max. 6 / m) Každý motoneruon dostává info z cca synapsí EPSP u synergistů IPSP u antagonistů Poloha synapse důležitá Některé EPSP se z pravidla vymykají EPSP mají různé trvání, přicházejí v různý čas Časová a prostorová sumace
 Každý motoneruon dostává info z cca synapsí. EPSP u synergistů. IPSP u antagonistů. Poloha synapse důležitá. Některé EPSP se z pravidla vymykají. EPSP mají různé trvání, přicházejí v různý čas. Časová a prostorová sumace.")
42
Schemata jednoduchých neuronálních zapojení
B Feed-forward inhibice C D Feedback inhibice

43
Inhibiční mechanismy Feed forward – dopředná inhibice
Feed back – zpětnovazebná Postsynaptická, presynaptická) (rozdíl v latenci, ovlivněné vstupem nebo výstupem,..) Reciproční inhibice – kombinace obou Laterální inhibice Disinhibice a disfacilitace při konvergenci EPSP a IPSP
 (rozdíl v latenci, ovlivněné vstupem nebo výstupem,..) Reciproční inhibice – kombinace obou. Laterální inhibice. Disinhibice a disfacilitace při konvergenci EPSP a IPSP.")
44
Inhibiční okruhy kontrolující přenos informace z aferentních vláken
Vzdálená inhibice Feed-back inhibice Přepojovací neuron Motorické jádro Feed-forward inhibice Mícha Vstup

45
Vlastnosti monosynaptického reflexního oblouku
Specificita Funkční determinace Účinnost Zkušenost Reflex ze 2 neuronů je abstrakce

46
Monosynaptický reflexní oblouk
Ia aferenty končí na svalech homonymních a heteronymních (=synergisté na daném kloubu) Ia aferent (ze svalových vřetének téhož svalu) a motoneuron (homonymní sval)
 Ia aferent. (ze svalových. vřetének téhož. svalu) a motoneuron. (homonymní sval)")
47
Vřazení excitačního interneuronu
Ia vlákna se přepojují na motoneurony také přes excitační interneuron - zpoždění – základ pro facilitaci Ia aferent Feed-forward facilitace a motoneruon

48
Vřazení inhibičního neuronu
Flexor Ia aferent Extenzor Ia aferent Přenos info mezi Ia aferenty a příslušnými motoneurony je kontrolován presynaptickou inhibicí - feed forward inhibice po předcházejí stimulaci Extenzor Flexor

49
Ia aferent Renshawova bb. a motoneuron Přenos info mezi Ia
aferenty a příslušnými motoneurony je kontrolován feed back inhibicí Prostřednictvím Renshawových bb. Ia aferent Renshawova bb. Analogie laterální inhibice – posílení aktivity nejvíce aktivovaných motoneuronů na úkor méně aktivovaných a motoneuron

50
R R Extenzor I a aferent Reciproční inhibice Extenzor Flexor
Ia inhibiční dráha 2 synaptická Extenzor I a aferent R R Reciproční inhibice Extenzor Flexor

51
Zkřížený napínací reflex
Nociceptor Extenzor Flexor

52
Vřazení inhibičního interneuronu
Ib aferent Extenzoru (šlachová tělíska) Inervace motoneuronů Ib aferenty Extenzor Flexor
 Inervace. motoneuronů. Ib aferenty. Extenzor. Flexor.")
53
Mícha Zkřížený napínací reflex Uhýbací reflex Šedá hmota Bílá hmota Dráždění nociceptorů Inhibiční interneuron EXTENZE FLEXE Výstup k extenzoru Výstup k extenzoru Výstup k flexoru Výstup k flexoru Motoneurony Excitační Inhibiční Reciproční inhibice

54
Konkrétní příklad míry konvergence a divergence aferentních signálů
trojhlavý lýtkový sval kočky 150 vřetének Každé vřeténko tvoří 2-5 synapsí, 90% končí na homonymním svalu, 60% končí na heteronymních svalech, 750 tricepsových motoneuronů Míra divergence jednoho Ia vlákna je 1:500 Míra konvergence na jeden motoneuron je 1: 500

55
Výsledek difuzního šíření informace - flexibilita reflexů
Reflexy – nepředpokládaně flexibilní Různé smyslové vstupy reflexy různě modifikují Také el. stimulace sousedících míst míchy vyvolává různé reflexní modifikace Aktivace různých souborů interneuronů, které následně kontrolují aktivitu motoneuronů Tyto různé soubory interneuronů mohou být aktivovány mozkem (volní pohyb) nebo smyslovými vstupy - úprava postoje
 nebo. smyslovými vstupy - úprava postoje.")
56
Reflexy Základní funkční jednotka nervové soustavy
Odpověď na podráždění zprostředkovaná CNS 5 prvků: Receptor, aferetní dráha, centrum, eferentní dráha, efektor Dělení Mono- x polysynaptické Extero- x intero- x proprioceptivní Centrální x extracentrální Somatické x autonomní Podmíněné x nepodmíněné

57
Proprioreceptivní reflexy
Řízení svalového tonu Krátká reflexní doba Nepodléhají únavě Nepodléhají přímé kontrole z vyšších center Jsou segmentovány

58
Proprioreceptivní reflexy
Monosynaptické – (svalová vřeténka) Vzruchy ze svalových vřetének – působí Facilitaci a motoneuronů vlastního svalech, Inhibici antagonistů Napínací (patelární) reflex Polysynaptické – (Golgiho šlachová tělíska) Vzruchy se šlachových receptorů působí prostřednictvím interneuronů Inhibici a motoneuronů
 Vzruchy ze svalových vřetének – působí. Facilitaci a motoneuronů vlastního svalech, Inhibici antagonistů. Napínací (patelární) reflex. Polysynaptické – (Golgiho šlachová tělíska) Vzruchy se šlachových receptorů působí prostřednictvím interneuronů. Inhibici a motoneuronů.")
59
Polysynaptické reflexy
Exteroreceptivní reflexy Vřazení různých počtů interneuronů Delší reakční doba Řízené z vyšších Extenzorové – stimulace dotykových receptorů vyvolá extenzi – základ posturálních reflexů Zkřížený extenzorový reflex Flexorové – stimulace nociceptorů vyvolá flexi – základ obranných reflexů Spinální reflexy působící svalový spazmus – obranný reflex

60
Proprioreceptivní reflexy
Myotatické (napínací) reflexy – natažení svalu prostřednictvím intrafuzálních vláken vřetének vede ke kontrakci Obrácené myotatické (uhýbací) – přílišné natažení svalu prostřednictvím šlachových proprioreceptorů vede k relaxaci Reciproční inhibice
 reflexy – natažení svalu prostřednictvím intrafuzálních vláken vřetének vede ke kontrakci. Obrácené myotatické (uhýbací) – přílišné natažení svalu prostřednictvím šlachových proprioreceptorů vede k relaxaci. Reciproční inhibice.")
61
Postojové reflexy Změny prahů míšních napínacích reflexů
Lokální statická reakce spinální exteroreceptivní reflexy extenzorové odpovídá svaly podrážděné končetiny Segmentální statické reakce Součinnost více končetin Zkřížený extenzorový reflex Celkové statické reakce Nadřazeny základním postojovým reakcím Koordinace svalového tonu končetin i svalů trupu Mícha Retikulární formace Tonické šíjové reflexy – dráždění proprioreceptorů šíje Tonické labyrintové reflexy – dráždění vestibulárního aparátu Fázické labyrintové reflexy – udržování postoje v klidu - dráždění kinetického čidla rotačním pohybem hlavy – udržování postoje při pohybu Změny prahů míšních napínacích reflexů -změnou excitability mtoneuronů -změnou frekvence systému

62
Vzpřimovací reflexy Vyšší koordinace statických reakcí
Usměrňování těžiště v klidu i v pohybu Integrační činnost všech výše jmenovaných motorických CNS center podporovaný činností Vestibulárního aparátu Korového centra subjektivního vnímání rovnováhy Iniciovány drážděním taktilních exteroreceptorů

63
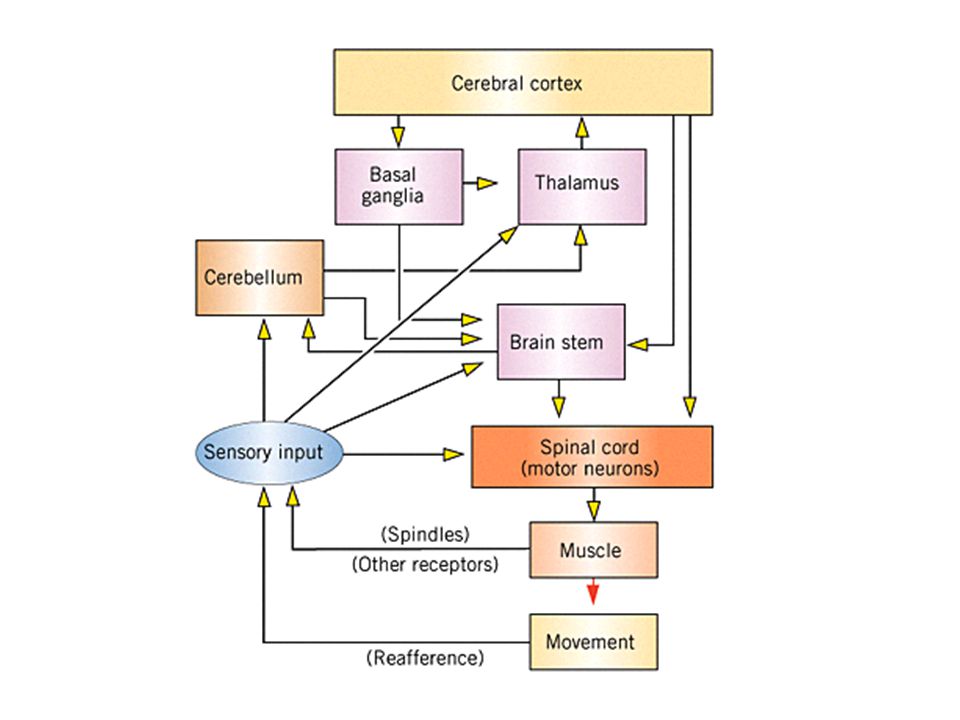
Přehled mechanizmů řízení hybnosti
Hlavní úrovně řízení motoriky Páteřní mícha Mozkový kmen Mozeček Bazální ganglia Talamus Kůra

64
Sestupné motorické dráhy
Retikulospinální – z retikulární formace Končí na míšních interneuronech, postojové reakce, reflexy trupu, nejstarší Vestibulospinální – vestibulární jádro Končí na míšních interneuronech, postoj a reakce v souvislosti s gravitací, kontrola flexorů, spojení s mozečkem Rubrospinální – červené jádro Stř mozek, přepojovací místo mozečku, končí na interneuronech Tektospinální – tektum Orientační reakce Kortikospinální – kůra, dráhy pro zahájení pohybu Pyramidové dráhy – kříží se v medulle v obl. Pyramid U člka a primátů tvoří monosynaptické spoje na míšních motoneuronech, manipulace Extrapyramidové dráhy – posturální reflexy

65
Sestupné motorické dráhy
Extrapyramidový systém Hrubé pohyby, postoj Víceneuronové spojení Začátek - premotorická oblast kůry Přepojení na Retikulospinální dráhu hlavní mimopyramidová dráha Proprioreceptivní reflexy Vestibulospinální dráhu Vzpřimovací reflexy Koordinace úmyslných a neúmyslných pohybů Tektospinální dráhu

66
Sestupné motorické dráhy
Pyramidový systém Jednoneuronové spojení kůry a motoneuronů Primární motorická kůra Tractus corticospinalis Tractus corticobulbaris Jemné, přesné a fázické cílené pohyby Úmyslné pohyby řízeny současně extra i pyramidovým systémem

67
Postranní kortikospinální*
Rubrospinální* Střední Postranní retikulospinální Vestibulospinální Tektospinální* Anteriorní Kortikospinální*

68
Přetětí Spinální preparát – všechny sestupné dráhy přerušeny
Ztráta volních pohybů Svalového tonu Po odeznění spinálního šoku se objevují míšní reflexy, zvířata můžeme postavit, při správné stimulaci mohou kráčet Decerebrovaný preparát (podíl prodloužené míchy) – tonická hyperaktivita extenzorů Stimulace napínacích reflexů retikulární formací a vestibulárními jádry Dekortikovaný preparát – spastitita daná excitačním vlivem retikulární formace, normálně inhibována kůrou
 – tonická hyperaktivita extenzorů. Stimulace napínacích reflexů retikulární formací a vestibulárními jádry. Dekortikovaný preparát – spastitita daná excitačním vlivem retikulární formace, normálně inhibována kůrou.")
70
Mícha VSTUPY: primární aferentní vlákna ze svalů, šlach, kloubů, kůže, podkoží VYSTUPY: a – a g motoneuronový systém Segmentované okruhy mezi Reflexní ústředí Místo CPG

71
Mozkový kmen Retikulární formace Vestibulární jádra Červené jádro

72
Střední mozek Červené jádro Most Mozeček Prodloužená mícha Retikulární
mostu Lokomoční oblast středního mozku Červené jádro Vestibulární jádro Most Mozeček Střední retikulární jádro Prodloužená mícha Dráha retikulospinální

73
Retikulární formace mozkového kmene
Multisynaptická síť malých neuronů Nespecifické ovlivnění Systém Vzestupný Sestupný Aktivační Inhibiční Specifická centra regulace dýchání, tlaku, trávení, endokrinní regulace,

74
Ascendentní aktivační systém Ascendentní inhibiční systém
Arousal – probouzecí reakce Ascendentní inhibiční systém Při poškození mozkového kmene – trvalé bdění Descendentní facilitační systém Zesílení míšních reflexů – řízení gama motoneuronů Aktivace ze statokinetického čidla, vestibulárních jader, jader mozečku, propriorecepce, exterorecepce, mozkové kůry Decerebrační rigidita Descendentní inhibiční systém Útlum míšních reflexů Aktivace z kůry, spinálního mozečku, bazálních ganglií

75
Mozkový kmen Jádra pro sestupné motorické info - začínají zde všechny sestupné mimopyramidové dráhy Jádra pro vzestupné sensorické info Prodloužená mícha, most Nepodmíněné obranné reflexy (kašel, kýcháí, zvracení) Regulace dýchání – pacemakerové inspirační a expirační jádra Regulace krevního oběhu a trávení Regulace mimických pohybů, fonace, řeč Regulace postojové motoriky Střední mozek Přepojování info sluchové a zrakové dráhy Centrum nepodmíněných reflexů (sluchové, zrakové, pohotovostní..) Mezencefalcká lokomoční oblast s generátory (např. chůze)
 Regulace dýchání – pacemakerové inspirační a expirační jádra. Regulace krevního oběhu a trávení. Regulace mimických pohybů, fonace, řeč. Regulace postojové motoriky. Střední mozek. Přepojování info sluchové a zrakové dráhy. Centrum nepodmíněných reflexů (sluchové, zrakové, pohotovostní..) Mezencefalcká lokomoční oblast s generátory (např. chůze)")
76
Motorická kůra Provádí pohyb
Výbírá svaly a svalové skupiny pro zamýšlený pohyb Somatotopické uspořádání Výstupy přímo do míchy (kortikospinální trakt) Nepřímo do mozkového kmene (kortikobulbální trakty) Vstupy z Premotorické, prefrontální a parietální kůry
 Nepřímo do mozkového kmene (kortikobulbální trakty) Vstupy z. Premotorické, prefrontální a parietální kůry.")
77
Primární motorická kůra Přídatná motorická kůra Centrální rýha Primární motorická kůra Premotorická kůra Centrální rýha

78
Motorická mapa šimpanze

79
Vlastnosti korových neuronů
Dynamické neurony – kódují změnu síly Statické neurony – kódují intenzitu síly Intermediální Neurony směrové preference kódují směr pohybu – pohyb veden ve směru, který je vektorem směroých preferencí aktvních neuronů

80
Premotorická kůra Výstupy premotorické kůry Programování
Mozkový kmen Mícha Kortikokortikální spoje s prefrontální a parietální kůrou Programování Rozhodování o pohybu Identifikace cílů

81
Výstupní informace motorické kůry pak zpracovávají:
Mozeček Bazální ganglia Retikulární formace Info pro pohyb veden k motoneuronům míchy Propojení ve složitých zpětnovazebných okruzích, které informují centrum o prováděném pohybu

82
– kůra limbická, front. lalok, podkorové limb. str.
Idea – kůra limbická, front. lalok, podkorové limb. str. Program asociační frontální limb. oblasti Mozeček rychlá, cílená motorika cerebrocerebellum Bazální ganglia pomalé, ustálené pohyby Premotorická a Motorická kůra Taktika a provedení Přes talamus Přes talamus Mozkový kmen Mícha

83
Mozeček Vestibulární mozeček – archicerebellum - vestibulocerebellum
Fylogeneticky nejstarší – vznik z vestibulárního jádra Smyslové vstupy z ucha a rovnovážného ústrojí Výstupy do vestibulárního járdra – oční pohyby, pohyby hlavy, posturální kontrola axiálních svalů těla Udržování vzpřímené polohy Spinální mozeček – paleocerebellum - spinocerebellum Fylogeneticky mladší Vstupy z periferie, výstupy různě Regulace svalového napětí Korový mozeček – neocerebellum - cerebrocerebellum Nejmladší Koordinace pohybů Přímé vstupy z motorické a premotorické kůry, nepřímé smyslové vstupy z míchy, mozkového kmene a kůry

84
Pontocerebellum Mozkový kmen Spinocerebellum Prodloužená
mícha Vestibulocerebellum Anterior lobe Posterior lobe Flocullonodullar lobe

85
Vestibulární nerv Vestibulární jádro Vestibulocerebellum

86
Retikulární formace Vestibulární jádro Spinocerebellum

87
Pontocerebellum Thalamus Červené jádro Most Spodní olivární jádro Nc. reticularis lateralis

88
Mozeček Kontrola pohybů a postoje
Nepřímo kontroluje segmentovaný motorický systém úpravou jiných motorických kontrolních signálů Složen z hlubokých jader a kůry Nc. Dentatus Nc. Fastigial Nc. interpositus

89
Propojení mozečku Vstupy do jader i kůry z
Periferních smyslových orgánů Mozkového kmene Mozkové kůry Výstupy do mozečkových jader Jádra mohou porovnávat aferentní signály se signály zpracovanými v mozečkové kůře

90
Výstupy Vstupy Trakt pontocelebellární Červené jádro Trakt
retikulocelebellární Trakt vestibulocelebellární Výstupy Retikulární jádro Vestibulární jádro Do talamu Nc.dentate Olivární jádro Červené jádro Trakt spinocelebellární Vstupy Retikulární jádro Fastigial a Interpositus ncc Vestibulární jádro

91
Mozeček 2 systémy excitačních vláken Mechová z Šplhavá z Míchy
Jader mozkového kmene Větví se do kůry a jader Končí na granulárních bb, tyto tvoří dlouhá paralelní vlákna, která jsou v kontaktu až s Purkyňovch bb. Speciální synapse – glomeruly – informace je sdílena s neurony- největší synapse mozku Šplhavá z Olivárního jádra Končí na Purkyňových bb – 1:1 Každé vlákno tvoří s P. bb mnohočetné kontakty

92
Buněčná organizace mozečku
Purkyňova buňka Paralelní vlákno Purkyňova buňka Granulární buňky Mechové vlákno Z různých zdrojů Z olivárního jádra Šplhavé vlákno Mozečkové jádro

93
ŠV MF PV P P GB Glomeruly Z olivárního Hluboká jádra mozečková
Z různých zdrojů

94
Základní koncept mozečkové sítě
Spočívá v existenci 2 mozečkových okruhů Primární – aferentní vlákna vedoucí do jader Sekundární – okruh kůry Sekundární moduluje okruh primární Změnou aktivity Purkyňových buněk

95
PV P P P GB GB MV CV CN Výsledkem operací v mozečkové kůře je
inhibiční výstup CN – mozečkové jádro CN MV CV

96
Purkyňovy buňky Mají extrémní dendritickou plochu, Spiny na dendritech Schopnost generovat denritické AP Ca kanály v aktivních zónách Axony PB tvoří kolaterály, které se větví kolem sousedních P. bb – výstupní neurony jsou propojeny – neobvyklé Synapse jenom axo-dendritické a axo-somatické, žádné dendro-dendritické PB dpovídají Komplexními spiky na podráždění ze šplhavých vláken Jednoduchými spiky na podráždění z mechových vláken

97
Komplexní spiky P. bb

98
Elektrofyziologická aktivita mozečkové kůry při neočekávaném
Během normálního pohybu PB pálí normálních spiků/s komplexní spiky se objevují 1/s Při neočekávaném se frekvence komplexních spiků zvyšuje Aktivita mozečkových jader je utlumena Info do kůry, korekce, vymizení útlumu

99
Uspořádání mozečkových okruhů je pravidelné
Pravidelnost je podkladem pro kódování časové posloupnosti při provádění pohybů aktivita jedné PB se šíří k sousedním, ty jsou aktivovány v intervalu daném svou vzdáleností

100
Long term deprese - LTD Soudí se, že je významným mechanismem motorického učení Vzniká, jestliže šplhavá a mechová vlákna pálí synchronně, Inhibuje přenos mezi paralelními vlákny a PB Snížuje citlivost GLU receptorů na subsynaptické membráně PB Šplhavá vlákna stimulují vtok Ca na dendritech PB Ca desensitizuje Glu receptory

101
Současné představy o LTD
Mozečková kůra tvoří modifikovatelnou postranní dráhu k Reflexnímu oblouku a Korovým informacím pro určitý pohyb Dráha vede přes mechové vstupy, paralelní vlákna a PB Přenos na dráze může být modifikován aktivitou šplhavých vláken Ta představují feed-back kontrolu chyb LTD inhibuje mozečkové okruhy zodpovědné za chyby

102
C D E C D E Šplhavá vlákna – vědomé pokyny Smyslový feedback
paralelním vláknům Paralelní vlákna – smyslový kontext Napoprvé Učení pohybu v mozečku Zahrnuje přestavbu Synapsí po opakované stimulaci C D E Následně C D E

103
Bazální ganglia Nc. Caudatus Putamen Globus pallidus
Velká jádra v kmeni Spolu se subtalamickým jádrem a subst. nigra Žádné přímé smyslové vstupy Žádné přímé výstupy Projekce zpět do kůry přes talamus Topografické uspořádání Fce neobjasněna Poškození vyvolává typické příznaky Parkinsonismus – poškození DOPAminergních nn. Subst. nigra

104
Thalamus Hlavní přepojovací centrum Jádra dělena - podle projekce
Specifická – aferentní přepojování do projekčních oblastí Nespecifická – 0 přímé spojení s dostředivými drahami Asociační – aktivována z projekčních jader talamu, hypotalamu a vedou do asociačních obl. k-ůry, hypotalamu a limbického systému Podle funkce Somatosensorická Převodní motorická přepojovací z mezimozku do limbického systému Z mezimozku do asociační kůry Širokých difuzních projekcí

105
Motorická jádra thalamu
Thalamická jádra se vstupy z globus pallidus a substantia nigra Úloha thalamu při kontrole motoriky je nesporná, ne zcela jasná

106
Bazální ganglia a thalamus tvoří několik složitých okruhů
Motorická kůra-putamen-globus pallidum-ventrolaterální jádro thalamu- přídatná motorická kůra Putamen-substantia nigra-thalamus Globus pallidum-subthalamické jádro Putamen-substantia nigra

107
Thalamus a bazální ganglia
Kritické pro normální motoriku Kontrolují postoj a koordinují pohyb Zahajují pohyb v souvislosti s taktilními a vizuálními podněty Zapojeny do vřazování pohybů do širšího behaviorálního a emočního rámce Postižení vede k narušení balance neurotransmiterů Cholinergního (striatum) a dopaminergního (substantia nigra) systému
 a dopaminergního (substantia nigra) systému.")
108
Lentiformní jádro Caudate nucleus - Putamen a Globus pallidus Thalamus Subthalamické jádro Substantia nigra (midbrain)
")
110
Motorika volní a mimovolní
Cílená (volní) motorika = motorický systém pohybu má složky: Kinetická volní (kůra – pyramidové dráhy) Kinetická mimovolní (neostriatum, neocerebellum) Statická mimovolní) (paleostriatum, archicerebellum, retikulární formace, spinální mícha) svalový tonus opěrná motorika - systém postojových a vzpřimovacích reflexů
 motorika = motorický systém pohybu má složky: Kinetická volní. (kůra – pyramidové dráhy) Kinetická mimovolní. (neostriatum, neocerebellum) Statická mimovolní) (paleostriatum, archicerebellum, retikulární formace, spinální mícha) svalový tonus. opěrná motorika - systém postojových a vzpřimovacích reflexů.")
111
Reflexy a chování Reflexy – na poč. 20. stol. – uvažovány jako základní moduly komplexního chování Věřilo se, že jeden reflex vyvolá smyslový vstup, který zase spouští další reflex Složité chování by podle této teorie bylo složeno ze série reflexů V současnosti se však soudí, že reflexy spíše chování modulují a „tvarují“

112
Motorika s centrálním programem - rytmické typy chování
Dříve se soudilo, že rytmické typy chování jsou sérií reflexů, kdy sensory feedback proběhlého reflexu spouští další Existuje však evidence, že základem mnoha rytmických typů chování jsou oscilátory resp. centrální aktivační generátory (central pattern generators) = sítě neuronů schopné produkovat přesně načasované série motorických impulzů i při absenci všech sensorických vstupů Smyslová informace není nezbytná, ale je významná
 = sítě neuronů schopné produkovat přesně načasované série motorických impulzů i při absenci všech sensorických vstupů. Smyslová informace není nezbytná, ale je významná.")
113
Deaferentace Zadní křídlo Přední křídlo ZK PK Donald Wilson 1961
Úhel křídla Čas Depresor ZK Elevator PK Depresor Elevator Donald Wilson 1961

114
Modely generátorů aktivity
Vstup, že se poletí Vstup A a B neurony – rozdílné prahy, reciproční inhibice Excitační synapse Inhibiční synapse K elevátoru K depresoru Wilsonův model

115
Modely tvorby rytmického výstupu při absenci rytmického vstupu
Inhibiční synapse Vstup

116
Předcházející model není stabilní
Aby fungoval, vlastnosti neuronů musí být nastaveny v úzkých limitech Stabilizace modelu možná zavedením jevu „postinhibiční excitace“ Tj. schopnosti neuronů produkovat krátké salvy AP po vyvázání z inhibice i při absenci excitačního vstupu

117
Postinhibiční excitace
U některých neuronů změna membránového potenciálu způsobí změnu prahu dráždivosti Hyperpolarizace – sníží, depolarizace zvýší práh dráždivosti Změna prahu dráždivosti dána změnami počtu Na+ nebo jiných napěťově řízených kanálů, které jsou v inaktivovaném stavu Hyperpolarizace snižuje počet inaktivovaných kanálů a protože snižuje práh, spiky je snadnější vyvolat

118
A B C Vliv hyperpolarizačního stimulu na práh stimulované buňky Práh
Normální neuron Em Neuron s postinhibičním reboundem B Práh Em Spikům se dá zabránit při pomalém odstraňování inhibice Práh C Em

119
Generátory skutečně existující
Neuronální sítě - komplexnější Principy reciproční inhibice Různé generátory různé dílčí detaily Málo známo

120
2 generátory - pylorický - CPG1
gastrický - CPG2 Humr Stomatogastrický nerv Stomatogastrické ganglion CPG1 CPG2 Střevo 1 3 2 Pylorický (vrátníkový) vak Žaludeční drtič Jícen Srdeční vak Esophageální ganglion Ústní otvor
 vak. Žaludeční. drtič. Jícen. Srdeční vak. Esophageální ganglion. Ústní otvor.")
121
Pylorický generátor humra
1 4 5 6 2 P 3 1 2 3 5 6 1 s Excitační synapse Elektrické spoje Inhibiční synapse

122
Gastrický generátor humra
B C E D A B P D F E C F Skupina neuronů 10 s

123
Inhibiční neurony Laterální neurony Excitační interneurony Motoneurony CPG mihule - plavání

124
Mechanismy pohybové koordinace
Každý segment s párem přívěsků obsahuje pár generátorů aktivity navzájem spřažených Langusta CPG aktivován z mozku Ovládá svaly jedné swimmerety CPG článků spojeny – koordinačními neurony - synchronizace (při přetětí – narušení koordinace) Swimmerety
 Swimmerety.")
125
Generátor dýchání Neuronová síť obsahuje neurony
Preinspiračními (Pre I) – jejich aktivita předchází výbojům respiračních motoneuronů Inspiračními (Insp I-III) – aktivní během inspirační fáze Expiračními (Exp) – hyperpolarizovány během inspirace Excitační výboje - Non-NMDA-receptory Inhibiční spoje - GABA Neuronální síť odlišná po narození a v dospělosti
 – jejich aktivita předchází výbojům respiračních motoneuronů. Inspiračními (Insp I-III) – aktivní během inspirační fáze. Expiračními (Exp) – hyperpolarizovány během inspirace. Excitační výboje - Non-NMDA-receptory. Inhibiční spoje - GABA. Neuronální síť odlišná po narození a v dospělosti.")
126
Dýchání savci Kmen – respirační skupiny jader:
Pontinní respirační skupina Klíčová struktura ukončení inspirace - vypínač Centrum v prodloužené míše: Dorzální skupina Přepojovací centrum respiračních reflexů z viscerálních orgánů Ventrální skupina Klíčové pro vznik rytmické aktivity - střídání vdechu a výdechu

127
Generátor inspiračního
Pontinní respirační skupina + - Generátor Respiračního Rytmu (Pre-I) Generátor inspiračního Rytmu (Insp) + Excitační Synaptická spojení + Inspirační motorický Výstup C4 Exp
 Generátor inspiračního. Rytmu. (Insp) + Excitační. Synaptická. spojení. + Inspirační motorický. Výstup. C4. Exp.")
128
CaL CaN KCa CaP H CaT NaP KIR Záznam akčního potenciálu neuronů tvořících respirační rytmickou aktivitu

129
CPG jsou subjektem regulace
CPGs – neuronové sítě kontrolují rytmické chování prostřednictvím fixního propojení jednotlivých neuronů Aktivita CPGs není fixní Může být modulována prostřednictvím změny synaptické síly synapsí umístěných na jednotlivých neuronech Tím dochází k „výběru“ určitých neuronů, které ovlivňují aktivitu systému za daných podmínek

130
Ovlivnění motorických výstupů smyslovými vstupy
Smyslové vstupy mají klíčový význam při motorických akcích V obecné rovině se podílí na Přizpůsobení plánovaného motorického výstupu podmínkám okolí Stabilizace prováděného pohybu Kompenzace nepředpokládaných událostí

131
Stabilizace prováděného pohybu
Svalová vřeténka a vestibulární systém -hlavní systémy, které se u obratlovců na stabilizaci pohybu podílejí Kompenzace nepředpokládaného Různé systémy a principy – Přímá regulace aktivity motoneuronů svalů Vliv na aktivitu CPG

132
a motoneuron g motoneuron Ia, II aferenty g motoneuron Váha odhadnutá správně Lehčí než předpokládaná Těžší Střední aktivita Vysoká aktivita Nízká aktivita

133
Vliv smyslových informací na CPG
Smyslové informace zpětně vedeny do motorických generátorů a koordinují CPG Plavání malých žraloků (Máčka skvrnitá) Deaferentace míchy – neovlivnila schopnost plavání – evidence o centrálně vytvářených motorických programech Decerebrace, kurarizace, oddělení ocasní míchy, mechanické ohýbání těla, elektrofyziologické snímání Citlivost ke smyslové modulaci
 Deaferentace míchy – neovlivnila schopnost plavání – evidence o centrálně vytvářených motorických programech. Decerebrace, kurarizace, oddělení ocasní míchy, mechanické ohýbání těla, elektrofyziologické snímání. Citlivost ke smyslové modulaci.")
134
Kurarizace – blokáda Ach receptorů – blokáda nervosvalového spojení
Manipulace ocasem Přetětí míchy Obnažení míchy - elektrody na motorických výstupech Míšní nervy Přetětí mozkového kmene Máčka skvrnitá CPG produkuje cykly 3,5 - 5 s Smyslové info generované mechanickým ohýbáním ocasu dominuje a mění frekvenci CPG

135
Nezávislý pohyb kol 1- měnitelný odpor 2- amputované prostřední končetiny Smyslový feedback a časování Stejný odpor pohybu P L 15x tíž vlevo P L Hmyz - přední a zadní noha na jedné straně a prostřední noha na protější straně se pohybují současně

136
Smyslový feedback a časování
Smyslové informace ze strany se zátěží ovlivňuje pohyb končetin na straně bez zátěže, pohyb je pomalejší, ale koordinace je zachována Po amputaci prostředních noh se přední a zadní nohy se pohybují alternativně - smyslové informace mění časování

137
Pacemakerová aktivita
Kir kanály obrácené usměrňovače směřující proud K do buňky Aktivovány SA, NA, A, DOPA, prostaglandiny, Cl, GABA, GLY, opiáty, somatostatin, Ach, TRH, cAMP, tachykininy Neselektivní H+kanály proud kationtů do b. – pomalá depolarizace Na kanály Ca++ kanály (3 typy) - depolarizace KCa kanály - repolarizace
 - depolarizace. KCa kanály - repolarizace.")
138
Centrální generátory aktivity - shrnutí
Řeč byla o alternující aktivitě antagonistických svalů jednoho tělního přívěsku nebo segmentu Jak vidno, skutečné generátory jsou velmi komplexní a rozmanité Využívají reciproční inhibici mezi interneurony Jsou součástí hierarchického systému motorické kontroly

Podobné prezentace