Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

41
Vége

42
Úvod do cytologie 2009

43
Přednáška je přežitek středověku, kdy neexistoval tisk a knížku měl jedině profesor. Proto profesor sám přednášel text, četl ho. Dnes už máme tisk, ale pořád to jaksi nebereme na vědomí. Přednášky jsou potřebné jen ve třech případech. Za prvé je to úvodní přednáška, kdy je studentům zapotřebí říci, co mají číst; za druhé je to výuka jazyků (mezi jiným i logiky) a za třetí, jestliže profesor provádí nějaké zkoumání a referuje o něm. (Bochenski, J., (2001) Mezi logikou a vírou. Barrister & Principal, Brno, str. 31)
 a za třetí, jestliže profesor provádí nějaké zkoumání a referuje o něm. (Bochenski, J., (2001) Mezi logikou a vírou. Barrister & Principal, Brno, str. 31).")
44
Většina buněk má velikost 1μm- 100μm
1m = 1000 mm 1mm = 1000μm 1μm = 1000 nm 1nm = 10-3μm = 10-9 m

46
„His /Schrödinger/ starting point was to ask what he called an „odd, almost ludicrous question: Why are atom so small?“ No doubt every schoolchild, frustrated that matter is made of particles to small to be seen, has wondered the same thing. However, Schrödinger realized that the more important question was a little different: „Why must our bodies be so large compared with the atom?“ The answer he gave was that we are big relative to atom to insulate us from atomic-level events. Schrödinger reasoned that in order to work properly, organisms need to base their physiologies on predictable, physical laws. Imagine the chaos that would result if we could hear each atom that smashed against our eardrum, react to each water molecule touching our skin, see the very oxygen and nitrogen that we inhale with every breath.

47
Because we are so large relative to the atom, we can live in a world where our bodies only have to deal with events caused by hundreds of thousands of atoms. Because our parts are so large, we are able to live in an orderly, predictable world in which the random movements of molecules and the haphazard chaos of molecular movements disappear into the backround. That´s why atoms are so small.“ (Miller, K.R. (1999) Finding Darwin´s God. Cliff Street Books. HarperCollins Publishers, New York. p. 205)
 Finding Darwin´s God. Cliff Street Books. HarperCollins Publishers, New York. p. 205)")
48
Historie

49
Dějiny buňky musí existovat nepřerušená linie buněk vzniklých mitosami až k první buňce cca před 3,5 miliardami lety i každá buňka našeho těla má své dějiny – v nepřerušené linii mitos se lze dostat až do chvíle, kdy moje tělo tvořila jedna jediná buňka - zygota

50
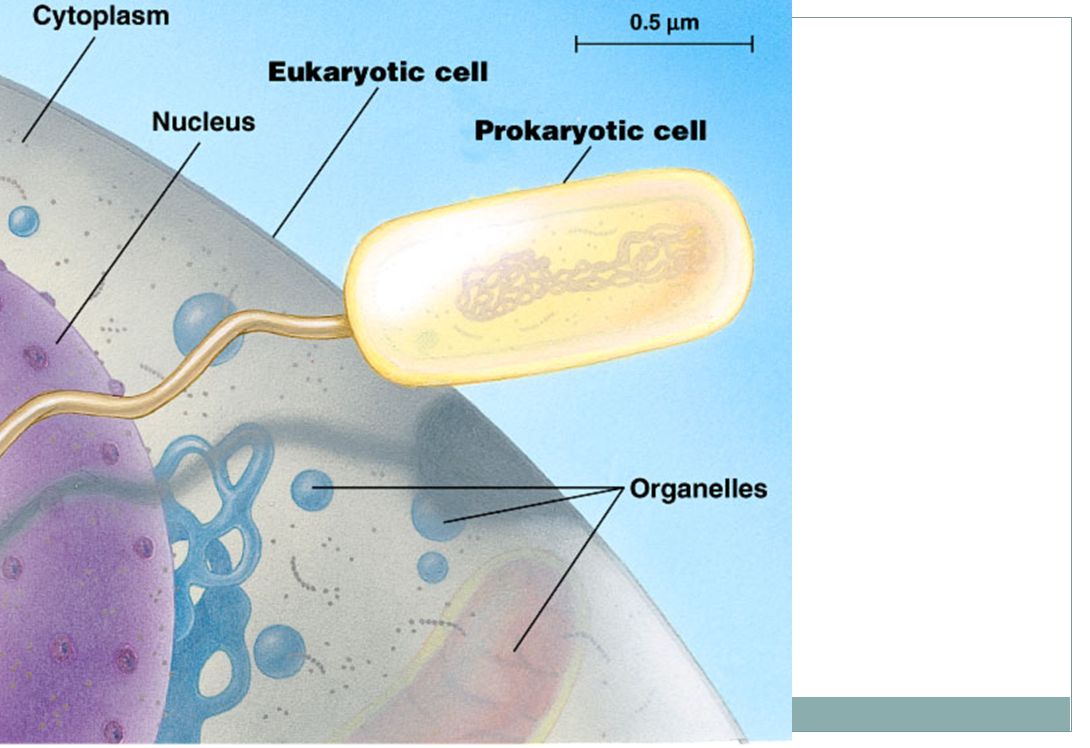
Rozdíly mezi prokaryotickou a eukaryotickou buňkou
Prokaryota (pro=před; karyon=jádro) DNA je v oblasti zvané nukleoid a není ohraničena membránou vůči buněčnému obsahu organely ohraničené membránami chybí buňka v obvykle v rozmezí 1μm - 10μm Eukaryota (eu=pravý; karyon=jádro) Chromosomy jsou v jádře, které je ohraničeno dvěma membránami v cytoplazmě jsou organely, ohraničené membránami buňka je obvykle mnohem větší (10μm - 100μm) než u prokaryot
 DNA je v oblasti zvané nukleoid a není ohraničena membránou vůči buněčnému obsahu. organely ohraničené membránami chybí. buňka v obvykle v rozmezí 1μm - 10μm. Eukaryota. (eu=pravý; karyon=jádro) Chromosomy jsou v jádře, které je ohraničeno dvěma membránami. v cytoplazmě jsou organely, ohraničené membránami. buňka je obvykle mnohem větší (10μm - 100μm) než u prokaryot.")
51
Rozdíly mezi prokaryotickou a eukaryotickou buňkou

52
Velikost eukaryotické buňky
oproti baktériím mají kompartmentalizaci, ketrá jim zřejmě umožňuje větší velikost buňky obvykle se ale důležité metabolické děje neodehrávají hlouběji než 15 µm – 25 µm pod povrchem, což omezuje průměr buňky na cca 50 µm obvyklá eukaryotická buňka má průměr cca 10µm – 30 µm některé neurony u člověka až 1 m! savčí oocyt 100 µm největší buňka = pštrosí vejce o průměru cca 20 cm (24 slepičích)!
!")
53
Prokaryontní buňka

55
Rozdíly mezi živočišnou a rostlinnou buňkou
Rostlinná buňka nemá: lyzosómy centrioly bičíky (s některými výjimkami) Živočišná buňka nemá: chloroplasty centrální vakuolu a tonoplast buněčnou stěnu plasmodesmata
 Živočišná buňka nemá: chloroplasty. centrální vakuolu a tonoplast. buněčnou stěnu. plasmodesmata.")
56
Živočišná buňka

57
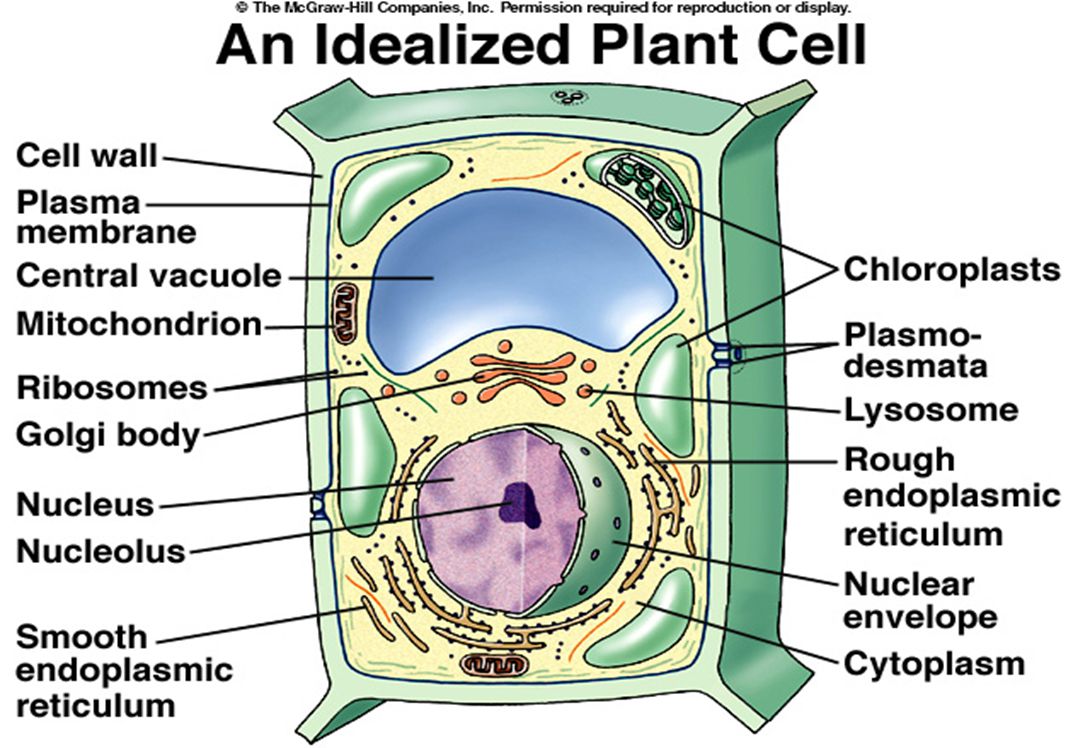
Rostlinná buňka

58
Rostlinná buňka na rozdíl od živočišné
Je na svém povrchu kryta buněčnou stěnou Obsahuje vakuolu či vakuoly ohraničené membránou zvanou tonoplast Obsahuje plastidy

59
Užitečné pojmy Protoplast = vše, co je ohraničeno buněčnou stěnou (=plazmatická membrána + vnitřek buňky) Kompartmentace = členění buňky na různé prostory (díky kompartmentaci mohou v různých místech, kompartmentech, buňky probíhat opačné chemické reakce, např. syntéza a rozklad téže látky) Pool = suma molekul jednoho typu v daném kompartmentu, sloužící jako zásoba pro biochemické reakce Symplast = kontinuum protoplastů (= protoplasty všech buněk rostliny spojené plazmodezmaty Apoplast = kontinuum buněčných stěn
 Pool = suma molekul jednoho typu v daném kompartmentu, sloužící jako zásoba pro biochemické reakce. Symplast = kontinuum protoplastů (= protoplasty všech buněk rostliny spojené plazmodezmaty. Apoplast = kontinuum buněčných stěn.")
61
V buňce je pořádná tlačenice…
Povrch buňky, endoplasmatické retikulum, Golgiho aparát mitochondrie jádro

62
Jádro (Nucleus) v průměru až 5μm
jaderná membrána je dvojitá, mezi membránami je prostor nm póry mají průměr okolo 100 nm Nukleární lamina na jaderné straně membrány udržuje tvar jádra. Tvoří ji intermediátní filamenta v prostoru jádra je zřejmě ještě jeden typ lešení, nukleární matrix

63
Nucleus

64
Jádro Jádro má na svém povrchu dvojitou membránu. Vnější membrána plynule přechází do endoplazmatického retikula. Jádro je tvořeno DNA a bílkovinami. Jadérko (nukleolus) je místem syntézy prekurzorů rRNA. Někdy jsou v buňce 2 i více jadérek; záleží na organismu a na fázi buněčného cyklu
 je místem syntézy prekurzorů rRNA. Někdy jsou v buňce 2 i více jadérek; záleží na organismu. a na fázi buněčného cyklu.")
65
Jádro na obrázku a na fotografii
Jaderné póry (na fotografii modře). Póry jsou vystlány komplikovanou proteinovou strukturou
. Póry jsou vystlány komplikovanou. proteinovou strukturou.")
66
Jádro Jaderná membrána (karyotéka) ohraničuje tzv. karyoplazmu, která obsahuje chromatin. Chromatin = DNA, histonové bílkoviny + bílkoviny nehistonové povahy. Přesto že jsou jaderné póry velké v průměru, je přísně kontrolováno, které látky proniknou dovnitř a které ven. Póry mohou tvořit až 20% povrchu jádra Perinukleární prostor

67
Stavba chromozómu Histonový oktamer 200 párů bazí 146 párů bází

68
Stavba chromosomu

69
jeden chromosom = jedna molekula DNA
na tomto obrázku je zobrazenou jen méně než 30 % DNA daného chromosomu

70
Typy chromosomů p = petite (malý); q = následující písmeno abecedy
na fotografii uprostřed chromosomy člověka během mitosy

71
Sesterské chromatidy Heterochromatin = kondenzované spiralizované části chromozómů Euchromatin = dekondenzované despiralizované části chromozómů

72
Ribozómy Vázané ribozómy jsou na povrchu ER
Volné ribozómy jsou v cytosolu Každý ribozóm se skládá ze dvou podjednotek. Buňky metabolicky aktivní mají velké množství ribozómů. Buňky pankreasu člověka např. až několik miliónů.

73
Ribozómy

74
Rozdíly mezi eukaryotickými a prokaryotickými ribozómy
sedimentační konstanta 80S velká podjednotka 60S 5S rRNA 5,8S rRNA 28S rRNA 40 proteinů malá podjednotka 40S 18S rRNA 30 proteinů Prokaryotické: sedimentační konstanta 70S velká podjednotka 50S 5S rRNA 23S rRNA 34 proteinů malá podjednotka 30S 16S rRNA 21 proteinů

75
Endoplazmatické retikulum

76
Endomembránový systém
Patří sem endoplazmatické retikulum (ER), Golgiho aparát, jaderný obal, lysozómy, tonoplast (u rostlin), plazmatická membrána sice není endomembrána, ale je spojena s ER a dalšími vnitřními membránami Nepatří sem plastidy a mitochondrie
, Golgiho aparát, jaderný obal, lysozómy, tonoplast (u rostlin), plazmatická membrána sice není endomembrána, ale je spojena s ER a dalšími vnitřními membránami. Nepatří sem plastidy a mitochondrie.")
77
Endoplazmatické retikulum
Membrány ER jsou nejhojnějšími membránami buňky (asi polovina všech membrán buněk) Velikost se může měnit (i během několika minut) v závislosti na vnitřních i vnějších faktorech ER s připojenými ribozómy = drsné endoplazmatické retikulum (tvorba proteinů) ER bez ribozómů = hladké endoplazmatické retikulum (tvorba lipidů) ER prochází plasmodesmaty (u rostlin) – možná existuje i kontinuum ER v celém symplastu ER slouží i jako zásobárna iontů vápníku
 Velikost se může měnit (i během několika minut) v závislosti na vnitřních i vnějších faktorech. ER s připojenými ribozómy = drsné endoplazmatické retikulum (tvorba proteinů) ER bez ribozómů = hladké endoplazmatické retikulum (tvorba lipidů) ER prochází plasmodesmaty (u rostlin) – možná existuje i kontinuum ER v celém symplastu. ER slouží i jako zásobárna iontů vápníku.")
78
Endoplazmantické retikulum
Hladké ER: syntéza lipidů (např. steroidní pohlavní hormony metabolismus cukrů (v játrech se účastní přeměny glykogenu na glukózu detoxifikace drog a jedů (zejména v játrech) ve svalových buňkách přes něj pronikají ionty Ca++ při stimulaci svalového stahu
 ve svalových buňkách přes něj pronikají ionty Ca++ při stimulaci svalového stahu.")
79
Drsné ER Proteiny „na export“ (např. inzulin v pankreatu). Rostoucí polypeptidový řetězec proniká z ribozómu pórem do ER. Na tyto polypeptidy je většinou v ER navěšen oligosacharid a vzniká tak glykoprotein. Tyto proteiny pak opouštějí ER ve speciálních „bublinách“ z membrán, transportních veziklech
. Rostoucí polypeptidový řetězec proniká z ribozómu pórem do ER. Na tyto polypeptidy je většinou v ER navěšen oligosacharid a vzniká tak glykoprotein. Tyto proteiny pak opouštějí ER ve speciálních „bublinách z membrán, transportních veziklech.")
80
Endoplazmatické retikulum
Drsné endoplazmatické retikulum (na fotografii ribozómy zeleně) Hladké endoplazmatické retikulum
 Hladké endoplazmatické retikulum.")
81
Golgiho aparát Strana obrácená k ER bývá označována jako „cis“
Strana obrácená do cytosolu jako „trans“

82
Golgiho aparát

83
Golgiho aparát GA je tvořen tzv. diktyozómy. Diktyozóm je sloupec 3 – 7 plochým membránových vaků, cisteren. Soubor diktyozómů = Golgiho aparát Vnitřní obsah cisteren není propojen Funkce Přebírá membránové lipidy a proteiny od ER a distribuuje je dál (do plazmalemy či tonoplastu) Sekrece bílkovin „na export“ Úprava těchto bílkovin, především jejich glykozylace (= navěšení sacharidové skupiny) Tvorba a exocytózy polysacharidů (lapací slizy masožravých rostlin, slizy kořenové čepičky, hemicelulózy buněčné stěny, tvorba buněčné stěny po mitóze)
 Sekrece bílkovin „na export Úprava těchto bílkovin, především jejich glykozylace (= navěšení sacharidové skupiny) Tvorba a exocytózy polysacharidů (lapací slizy masožravých rostlin, slizy kořenové čepičky, hemicelulózy buněčné stěny, tvorba buněčné stěny po mitóze)")
84
Golgiho aparát Při exocytóze splývají váčky Golgiho aparátu s plazmatickou membránou, čímž dochází k přibývání jejího množství; proto probíhá opačným směrem endocytóza v jedné buňce mohou být až stovky GA v GA dochází rovněž k syntéze sacharidů, zejména (u rostlin) pektinů a dalších necelulózových sacharidů dnes vnímáme GA jako velmi dynamický systém
 pektinů a dalších necelulózových sacharidů. dnes vnímáme GA jako velmi dynamický systém.")
85
Golgiho aparát jednotlivé cisterny stárnou, dozrávají a mění se i se svým proteinovým nákladem ve směru od cis k trans na proteiny jsou navěšeny různé fosfátové skupiny, které slouží jako PSČ – ukazují budoucí destinaci proteinu, místo, kam má být dopraven. Transportní molekuly umí takovéto zprávy „přečíst“

86
Golgiho aparát (vznik lysozómů)
Alespoň některé lyzosómy vznikají z GA

87
Golgiho aparát Strana trans (směrem od ER) Strana cis (u ER)
 Strana cis (u ER)")
88
Lysozómy Obsahují enzymy schopné štěpit proteiny, polysacharidy, tuky, nukleové kyseliny kyselé prostředí - uvnitř lyzosómu je pH 5. Toto nízké pH vzniká pumpováním H+ dovntiř lyzosómu alespoň nějaké lyzosómy vznikají z veziklů uvolněných z trans strany GA

89
U prvoků lyzosómy pomáhají v trávení potravy
Měňavky a další prvoci fagocytují částečky potravy za vzniku potravní vakuoly (jevu se říká fagocytóza). Tyto potravní vakuoly pak v těle prvoka splývají s lyzosómy, jejichž sada trávicích enzymů potravu naštěpí Potravní vakuola Lyzosóm
. Tyto potravní vakuoly pak v těle prvoka splývají s lyzosómy, jejichž sada trávicích enzymů potravu naštěpí. Potravní vakuola. Lyzosóm.")
90
Další funkce lyzosómů Autophagie - lyzosóm je schopen pohltit nefunkční organelu, a monomery uvolnit do cytosolu k dalšímu použití. Takto buňka obnovuje samu sebe programovaná buněčná smrt je širší pojem než apoptóza (i když se někdy oba termíny používají jako synonyma) Apoptóza - lyzosómy se účastní též programované buněčné smrti, např. v smrti buněk ocasu při přeměně pulce v žábu či buněk mezi prsty lidského embrya. Mnohé nemoci jsou spojeny se špatnou funkcí lyzosómů, např. Tay-Sachsova nemoc
 Apoptóza - lyzosómy se účastní též programované buněčné smrti, např. v smrti buněk ocasu při přeměně pulce v žábu či buněk mezi prsty lidského embrya. Mnohé nemoci jsou spojeny se špatnou funkcí lyzosómů, např. Tay-Sachsova nemoc.")
91
Programmed Cell Death developmentally defective cells (T-lymphocytes)
Excess cells (brain development) Cells that serve no function (tadpole tail) Cells whose cycle is perturbed Virus infected Cells Chemotherapeutic Killing of Cells
 Cells that serve no function (tadpole tail) Cells whose cycle is perturbed. Virus infected Cells. Chemotherapeutic Killing of Cells.")
92
Apoptóza

93
Příklady programované buněčné smrti v těle člověka

94
Apoptosis programmed cell death initiated by a burst of thyroid hormone i s responsible for resorption of the tadpole tail Caenorhabditis: Of the 1090 somatic cells that are produced during embryogenesis of the C.elegans hermaphrodite, 131 undergo programmed cell death at reproducible locations and times.

95
Apoptosis during craniofacial development, the hard palate develops from two lateral precursors, each covered in a protective layer of epithelial cells. As the two halves grow together at the midline of the nasopharynx, they remain separated by this covering of epithelium until, in response to a developmental cue, the epithelial cells at the midline undergo programmed cell death. Then the two halves of the palate can fuse. Failure of the epithelial cells to die at the appropriate time can interfere with the fusion of the bone, causing the left palate. in male embryos, progenitors of the Mullerian ducts develop, even though they have no function. Programmed cell death eliminates the constituent cells of these embryonic ducts.

96
Semiautonomní organely (=plastidy a mitochondrie)
Nejsou součástí endomembránového systému Podle endosymbiotické teorie byly původně samostatné prokaryontní organismy Nevznikají v buňce de novo, ale udržují se dělením stávajících organel Mají vlastní genetický aparát (DNA + ribozómy). DNA je kruhová, bez histonů, a ribozómy jsou prokaryotického typu (70S) Mají dvě membrány Jsou alespoň potenciálně schopny tvorby ATP
. DNA je kruhová, bez histonů, a ribozómy jsou prokaryotického typu (70S) Mají dvě membrány. Jsou alespoň potenciálně schopny tvorby ATP.")
97
Současná představa vzniku eukaryontní buňky z prokaryontní Endosymbiotická teorie

98
Evoluční počátky eukaryot

100
Plastidy Ohraničeny dvojitou membránou
Chloroplasty – místo, kde probíhá fotosyntéza. Obsahují chlorofyl Chromoplasty – obsahují karotenoidy (karoteny a xantofyly). Způsobují žluté, červené a oranžové zbarvení květů, listů a plodů – ekologická funkce (lákání opylovačů) Leukoplasty – neobsahují barevné pigmenty. V některých dochází k syntéze škrobu – ty nesou název amyloplasty
. Způsobují žluté, červené a oranžové zbarvení květů, listů a plodů – ekologická funkce (lákání opylovačů) Leukoplasty – neobsahují barevné pigmenty. V některých dochází k syntéze škrobu – ty nesou název amyloplasty.")
101
Chloroplasty

102
Chloroplasty V rostlinné buňce je řádově 20 – 50 chloroplastů. Vnitřek chloroplastů se nazývá stroma. Stroma je protkáno systémem membrán, který vytváří plošné měchýřky zvané tylakoidy. Každý tylakoid se strukturálně podobá plastidovému obalu, neboť je tvořen dvěma membránami. Štočky tylakoidů naskládaných na sebe tvoří grana.

103
Chloroplasty chloroplasty rostlin a řas obsahují cca 20 – 80 kopií DNA
např. chloroplast tabáku obsahuje bp DNA obsahující 113 genů rekombinace mezi těmito DNA je extrémně vzácná u většiny kvetoucích rostlin se chloroplasty dědí matroklinně avšak alespoň u některých gymnosperm se dědí po „otci“ – z pylového zrnka u některých kvetoucích rostlin byla rovněž pozorována biparentální dědičnost

104
Chloroplasty zřejmě nejhojnější enzym na planetě, RuBisCO, který katalyzuje fixaci CO2 v rámci Calvino-Bensonova cyklu je molekulární chimérou. Skládá se ze dvou podjednotek: gen pro malou podjednotku se nachází v jaderném genomu gen pro velkou podjednotku se nachází v cpDNA (chloroplastové DNA)
")
105
Mitochondrie

106
Mitochondrie Buňky obsahují 1 - tisíce mitochondrií. Mitochondrie měří 1μm - 10μm. Vnější membrána je hladká a vysoce propustná. Vnitřní membrána je naopak velmi málo propustná. Vchlipuje se dovnitř a tvoří tzv. kristy. Vnitřní prostor mitochondrie se nazývá matrix. Funkce – Krebsův cyklus, elektrontransportní řetězec a tvorba ATP.

107
Mitochondrie na obrázku i na fotografii

108
Mitochondrie počet mitochondrií na buňku
4 u některých jednobuněčných Fungi tisíce ve svalech obratlovců u obratlovců je v každé mitochondrii cca 5 – 10 kopií DNA

109
Genom jádra a mitochondrií u člověka

110
Mitochondrie nervová a svalová buňka obsahuje tisíce mitochondrií
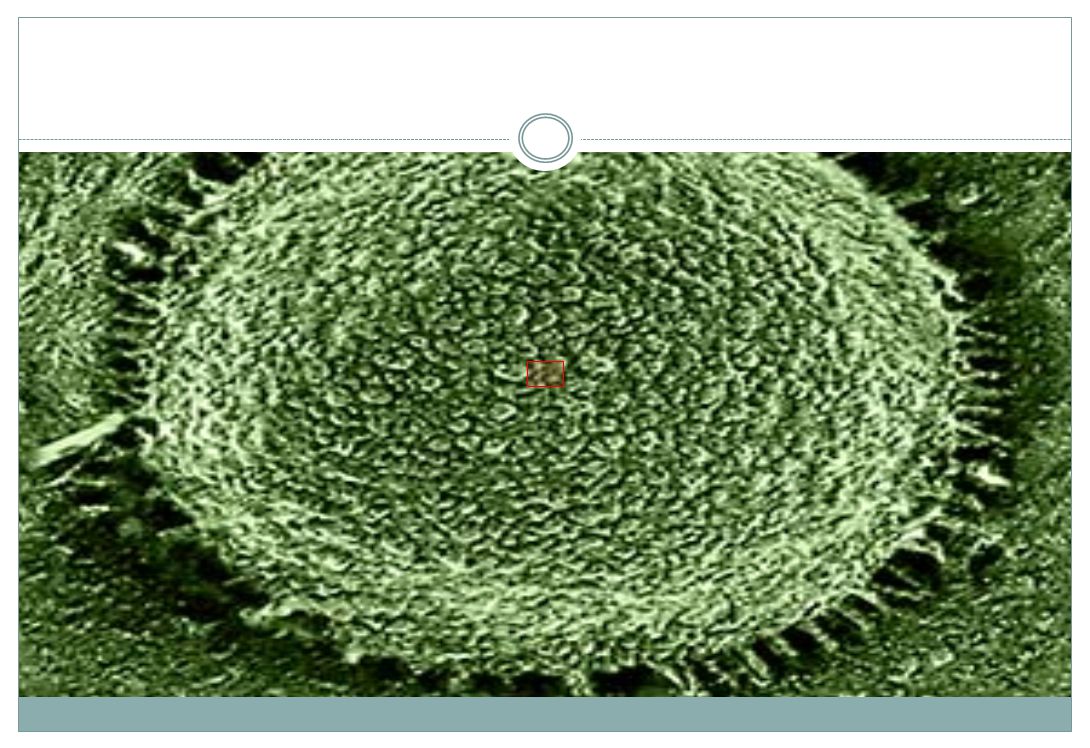
oocyt obsahuje asi mitochondrií spermie obsahuje 50 – 75 mitochondrií (do oocytu zpravidla proniknou, ale jsou zde nějak eliminovány)
")
111
Peroxizómy Jsou ohraničeny jedinou membránou. Získávají proteiny a lipidy v cytosolu, a když jsou velké, rozdělí se. z různých látek přenášejí vodík na kyslík, za vzniku peroxidu vodíku H2O2 peroxizómy umí peroxid vodíku vyrobit i rozložit některé peroxizómy užívají kyslík ke štěpení mastných kyselin na menší molekuly, které mohou být následně transportovány do mitochondrií jako palivo pro buněčnou respiraci

112
Peroxizómy peroxizómy v játrech pomáhají detoxifikovat alkohol
chybá funkce peroxizómů u člověka často ústí v mentální postižení glyoxyzómy jsou specializované peroxizómy v semenech rostlin. Přeměňují mastné kyseliny na cukry, dokud se u semenáčku nenastartuje fotosyntéza

113
Peroxizómy

114
Opakování

115
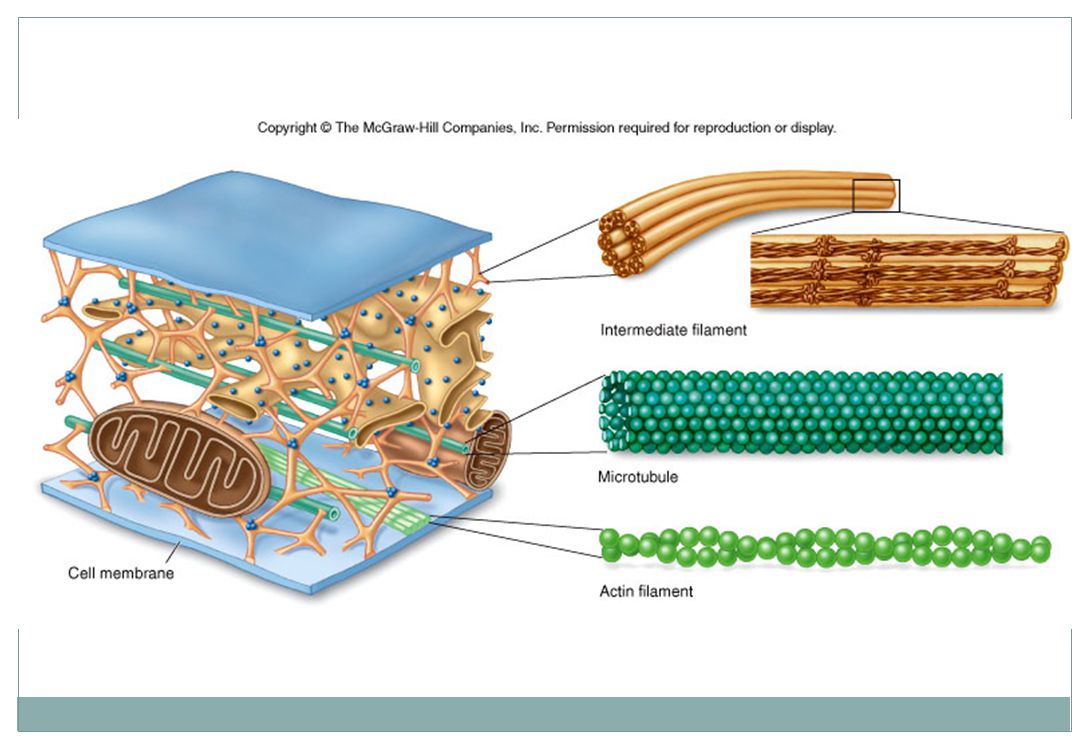
Cytoskelet Mikrotubuly, intermediární filamenta a mikrofilamenta
dávají buňce mechanickou podporu a udržují její tvar (=důležité zejména pro živočišné buňky) kotví mnohé organely a dokonce i některé enzymy podobně jako lešení může být rychle postaven či odbourán podle potřeby
 kotví mnohé organely a dokonce i některé enzymy. podobně jako lešení může být rychle postaven či odbourán podle potřeby.")
117
Cytoskelet tvoří mikrotubuly, mikrofilamenta a intermediální filamenta

118
Funkce cytoskeletu zajišťují buněčný pohyb: jak buňky samotné, tak i pohyb uvnitř buňky: pohyb řas a bičíků pomocí motorových molekul vezikly putují v buňce po „jednokolejkách“ cytoskeletu práce měchýřků naplněných neurotransmitery na koncích axonů pohyb potravní vakuoly při fagocytóze pohyb cytoplazmy rostlinných buněk regulují biochemickou aktivitu buňky přenášením mechanických impulsů až do jádra

119
Molekulární motory (a) molekulární motory pohání klouzání jednoho mikrotubulu po druhém (b) molekulární motory se mohou navázat k organele a „kráčet“ s ní podél mikrotubulu. Tímto způsobem se např. vezikly s neurotransmitery dostávají na konec axonů
 molekulární motory se mohou navázat k organele a „kráčet s ní podél mikrotubulu. Tímto způsobem se např. vezikly s neurotransmitery dostávají na konec axonů.")
120
Molekulární motory

121
Mikrotubuly U všech eukaryontních buněk
duté trubice o průměru 25 nm; dlouhé 200 nm - 25μm stěna mikrotubulu je tvořena globulárním proteinem zvaným tubulin. Každá tubulinová molekula je ovšem dimer, tvořený α-tubulinem a β-tubulinem. Mikrotubuly rostou přidáváním tubulinových dimerů k jednomu konci. Dimery ale mohou být i odbourány a použity jinde v buňce mikrotubuly vedou vezikly z GA směrem ven z buňky a jsou odpovědny za pohyb chromozómů při mitóze a meióze

122
Mikrotubuly: Centrosom a centrioly
Živočišná buňka má 2 centrioly v tzv. centrosomu. Centrioly jsou na sebe kolmě a jsou uspořádány v devíti řadách po třech. Modře jsou zančeny netubulinové proteiny, které drží mikrotubulové triplety pohromadě

123
Mikrotubuly: Centrosom a centrioly

124
Struktura mikrotubulu
(A) (C) kružnice mikrotubulu se sestává z 13 podjednotek tubulinových dimerů (D) boční pohled (E) schema jedné tubulinové molekuly αβ-dimerů. Díky tomuto uspořádání má mikrotubulus polaritu, odlišujeme + konec a - konec
 (C) kružnice mikrotubulu se sestává z 13 podjednotek tubulinových dimerů. (D) boční pohled. (E) schema jedné tubulinové molekuly αβ-dimerů. Díky tomuto uspořádání má mikrotubulus polaritu, odlišujeme + konec a - konec.")
125
Polymerace tubulinu na centrosomu
(A) Červeně jsou označeny tzv. nukleační místa, z nichž na centrosomu vyrůstají mikrotubuly (B) centrosom s napojenými mikrotubuly. Minus konec mikrotubulu je zanořen do centrosomu. Plus konec je volný a směřuje do cytoplazmy
 Červeně jsou označeny tzv. nukleační místa, z nichž na centrosomu vyrůstají mikrotubuly. (B) centrosom s napojenými mikrotubuly. Minus konec mikrotubulu je zanořen do centrosomu. Plus konec je volný a směřuje do cytoplazmy.")
126
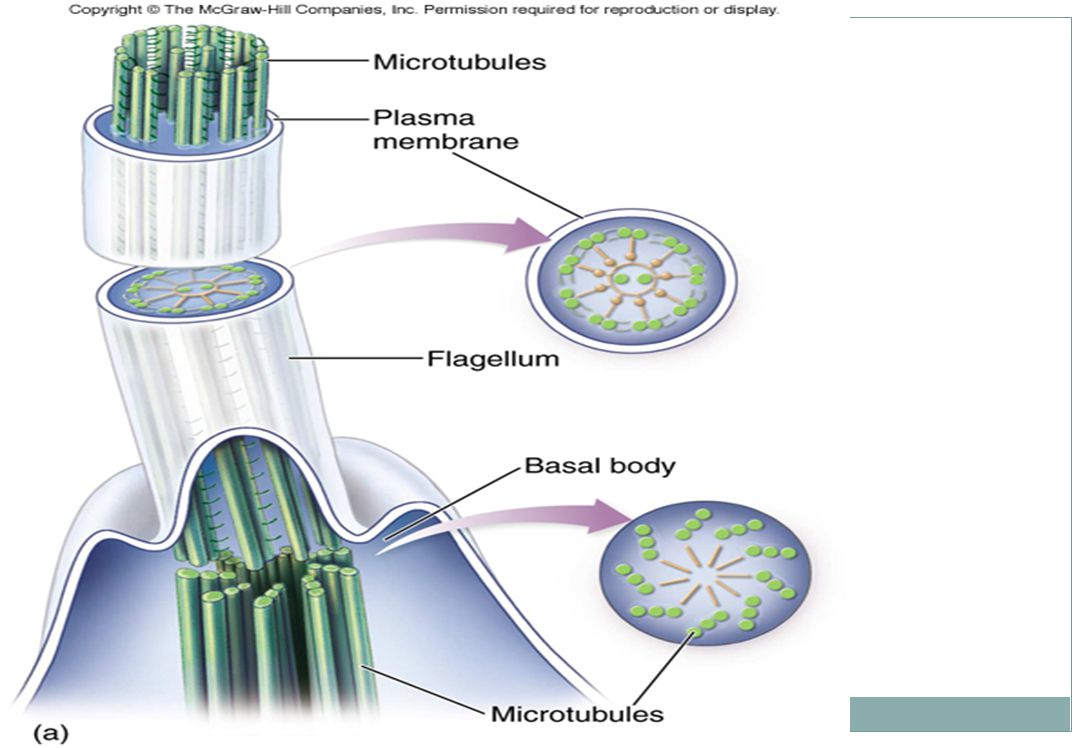
Cilia a flagella (řasy a bičíky)
Pohyb mnoha jednobuněčných organismů pohyb spermií živočichů, řas a některých rostlin výstelka části dýchací trubice, výstelka vejcovodů Stejným rysem brv i bičíků je vzorec „9+2“ = devět dvojic a uprostřed dva mikrotubuly volné. Rovněž jsou oba zakotveny do buňky tzv. bazálním tělískem, které má stejnou strukturu jako centriola

127
Cilia a flagella - srovnání
na povrchu buňky obvykle ve větším počtu průměr 0,25 μm, dlouhá μm pracují spíše jako vesla, pohyb buňky je kolmý k ose brvy Bičíky: jen jeden nebo jen několik na buňku průměr 0,25 μm, dlouhé μm pohyb buňky je v ose bičíku

128
Cilia a flagella - srovnání
Ačkoli se brvy a bičíky liší v délce, počtu na buňku, a způsobem práce, mají stejnou ultrastrukturu („9+2“).
.")
129
Cilia Struktura „9 + 2“ je zřejmě velmi starobylým vynálezem. Na horním obrázku vidíme brvy trepky, na dolním brvy v dýchací soustavě člověka.

130
(P)ohyb bičíků a brv Proteiny molekulárních motorů spojujících jednotlivé dvojice mikrotubulů se nazývá dynein. Tyto molekuly způsobují klouzání jednotlivých dvojic mikrotubulů mezi sebou. Jiné struktury drží celý bičík pohromadě, takže se neprodlužuje, ale ohýbá

133
Mikrofilamenta Plná vlákna 7nm v průměru
ve všech eukaryontních buňkách říká se jim rovněž aktinová filamenta, protože je tvoří molekuly aktinu jedno mikrofilamentum je tvořeno dvěma navzájem se ovíjejícími se aktinovými řetězci tvoří trojrozměrnou síť pod plazmatickou membránou a tak udržují tvar živočišné buňky. Proto má cytoplazma těsně pod membránou charakter gelu, zatímco uvnitř buňky se jedná spíše o sol nejznámější funkce: spolu s myosinem pohyb svalů. Myosin zde funguje jako molekulární motor, svými výběžky „kráčí“ podél aktinového vlákna

134
Mikrofilamenta 15 % proteinů eukaryotické buňky připadá na aktin

135
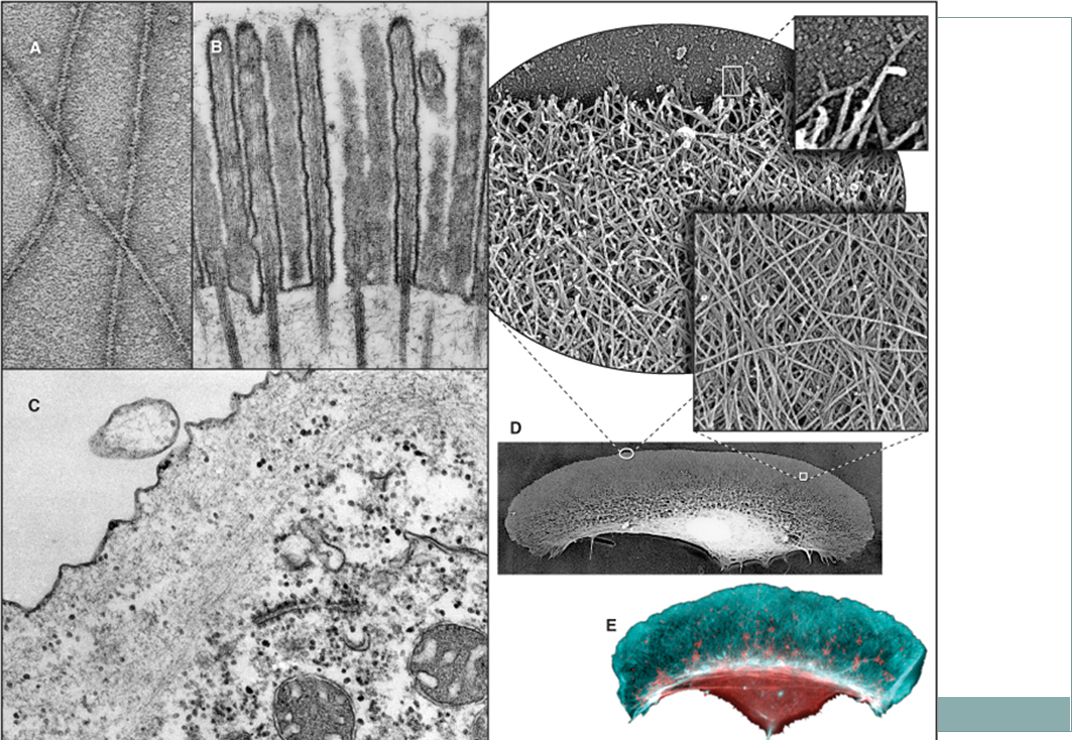
MICROGRAPHS OF CONTRACTILE BUNDLES OF ACTIN FILAMENTS
MICROGRAPHS OF CONTRACTILE BUNDLES OF ACTIN FILAMENTS. A, Fluorescence micrograph of a dividing normal rat kidney cell stained with fluorescein-phalloidin. Actin filaments are concentrated in the contractile ring in the constricting cleavage furrow. The drawing illustrates the filaments in the contractile ring. B, Fluorescence micrograph of a myofibril isolated from skeletal muscle and stained with fluorescein-phalloidin for actin filaments (green) and rhodamine-antibody to α;-actinin for Z disks (yellow). C, Electron micrograph of a thin section of skeletal muscle. (A, Micrograph courtesy of Y.-L. Wang, University of Massachusetts, Worcester. B, Courtesy of V. Fowler, Scripps Research Institute, La Jolla, California. C, Courtesy of H. E. Huxley, Brandeis University, Waltham, Massachusetts.)
 and rhodamine-antibody to α;-actinin for Z disks (yellow). C, Electron micrograph of a thin section of skeletal muscle. (A, Micrograph courtesy of Y.-L. Wang, University of Massachusetts, Worcester. B, Courtesy of V. Fowler, Scripps Research Institute, La Jolla, California. C, Courtesy of H. E. Huxley, Brandeis University, Waltham, Massachusetts.)")
136
ELECTRON MICROGRAPHS OF ACTIN FILAMENTS
ELECTRON MICROGRAPHS OF ACTIN FILAMENTS. A, Filaments of purified actin prepared by negative staining. B, A thin section of an intestinal epithelial cell illustrating finger-like microvilli with tightly packed bundles of actin filaments linked to the surrounding plasma membrane by myosin-I. The barbed ends of these filaments (see Fig. 33-9) are located at the tips of the microvilli. C, A thin section of Acanthamoeba showing the actin filament meshwork in the cortex beneath the plasma membrane. D-E, Cultured fish scale keratocytes, fixed while actively migrating toward the top of the figure. D, Electron micrograph of a whole mount of a cell illustrating the meshwork of branched filaments near the leading edge and longer, unbranched filaments deeper in the cytoplasm. Most filaments are oriented with their barbed ends forward. E, Fluorescence micrograph with phalloidin staining actin filaments (blue) and antibodies staining myosin II (red). (A, Courtesy of U. Aebi, University of Basel, Switzerland. B, Courtesy of L. Tilney, University of Pennsylvania, Philadelphia, and M. Mooseker, Yale University, New Haven, Connecticut. D-E, Courtesy of T. Svitkina and G. Borisy, University of Wisconsin, Madison.)
 are located at the tips of the microvilli. C, A thin section of Acanthamoeba showing the actin filament meshwork in the cortex beneath the plasma membrane. D-E, Cultured fish scale keratocytes, fixed while actively migrating toward the top of the figure. D, Electron micrograph of a whole mount of a cell illustrating the meshwork of branched filaments near the leading edge and longer, unbranched filaments deeper in the cytoplasm. Most filaments are oriented with their barbed ends forward. E, Fluorescence micrograph with phalloidin staining actin filaments (blue) and antibodies staining myosin II (red). (A, Courtesy of U. Aebi, University of Basel, Switzerland. B, Courtesy of L. Tilney, University of Pennsylvania, Philadelphia, and M. Mooseker, Yale University, New Haven, Connecticut. D-E, Courtesy of T. Svitkina and G. Borisy, University of Wisconsin, Madison.)")
138
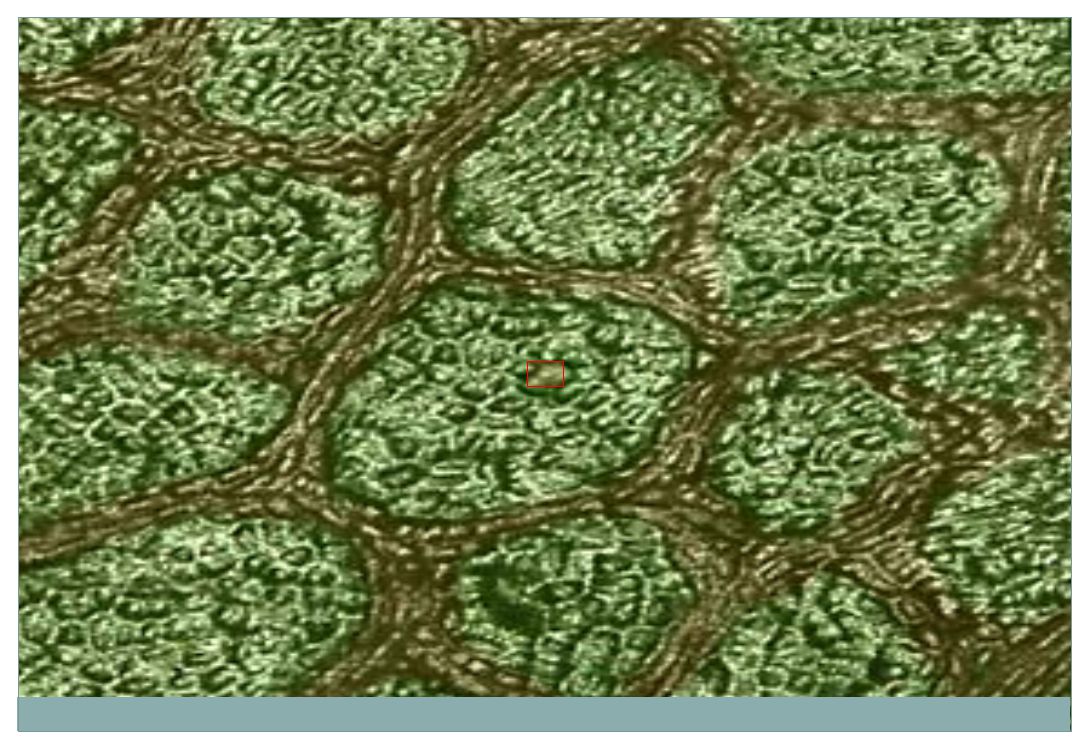
Mikrofilamenta Absorpční povrch střevní buňky je mnohonásobně zvětšen mikroklky, které jsou vyztuženy mikrofilamenty

139
Mikrofilamenta - funkce
důležitý při pohybu svalu výztuha mikroklků tvoří zaškrcení mateřské buňky při cytokinezi živočišné buňky aktinomyosinový pohyb také odpovídá za pohyb panožek (pseudopodií) např. měňavek a leukocytů proudění cytoplazmy; běžné zejména v rostlinných buňkách
 např. měňavek a leukocytů. proudění cytoplazmy; běžné zejména v rostlinných buňkách.")
140
Mikrofilamenta - příklady funkce

141
Intermediární filamenta (=střední filamenta)
Průměr nm (=mezi mikrotubuly a mikrofilamenty nemají jednotnou chemickou stavbu, podjednotky, které je tvoří patří mezi keratiny jsou mnohem stabilnější než mikrotubuly a mikrofilamenta, které se často staví a odbourávají udržují tvar buňky a pozici některých organel. Např. jádro je v buňce uloženo v „kleci“ z mikrofilament. Jiné intermediární filamenta tvoří nukleární laminu. Udržují také správný tvar axonového vlákna v nervové buňce

142
Struktura a funkce cytoskeletu

143
Cytoskelet Mikrotubuly bičíků a brv mají typické uspořádání „9+2“

144
Cytoskelet - přehled

145
Buněčná stěna = neprotoplazmatická část buňky, ukládaná vně od protoplastu. Od 0,1 μm po několik mikrometrů Složení: Střední lamela – tvořená pouze pektiny; je společná sousedním buňkám. Produkuje ji Golgiho aparát při buněčném dělení. Tvoří se od středu k okrajům Primární stěna – tvořena amorfní hmotou (hemicelulózy, pektiny, bílkoviny) a mikrofibrily celulózy – princip železobetonu. Primární stěna je schopna růst s buňkou Sekundární stěna – jen u některých buněk. Její vznik je spojen s ukončením růstu. Někdy výrazně zmenšuje lumen buňky. Ukládá se v ní lignin a suberin a kutin. Typická pro buňky mechanických a vodivých pletiv
 a mikrofibrily celulózy – princip železobetonu. Primární stěna je schopna růst s buňkou. Sekundární stěna – jen u některých buněk. Její vznik je spojen s ukončením růstu. Někdy výrazně zmenšuje lumen buňky. Ukládá se v ní lignin a suberin a kutin. Typická pro buňky mechanických a vodivých pletiv.")
146
Buněčná stěna Rostliny nejprve vytvoří primární stěnu, a u některých buněk po zastavení růstu i silnější sekundární stěnu. Lepkavá střední lamela drží buňky u sebe (pektiny se užívají i v džemech). Skrze plasmodesmata je cytoplasma obou buněk propojena.
. Skrze plasmodesmata je cytoplasma obou buněk propojena.")
147
Buněčná stěna Buňka Střední lamela Sekundární stěna Primární stěna

148
Buněčná stěna Funkce Mechanická pevnost buňky Příjem a transport látek v rostlině Zabraňuje prasknutí buňky v důsledku osmotického příjmu vody Tečky. V místě, kde prostupují plazmodesmata buněčnou stěnu, není stěna ztlustlá. Tato místa se označují jako tečky.

149
Plasmodesmata = kanálky procházející buněčnou stěnou, propojující sousední buňky. Nejsou v buňce rozptýleny rovnoměrně, ale shlukují se do tzv. primárních políček, kde je buněčná stěna značně ztenčená. V průměrné buňce jich je – Jsou vystlány plazmalemou.

150
Extracelulární matrix
U živočišných buněk tvořena zejména glykoproteiny vylučovanými buňkou. Nejhojnějším glykoproteinem je kolagen, který tvoří vlákna vně buňky (polovina bílkovin lidského těla je kolagen) dalšími látkami jsou proteoglykany, spojující kolagenová vlákna, a fibronektiny a integriny, které jsou zabudovány v plasmatické mebráně snad ovlivňuje i aktivitu genů
 dalšími látkami jsou proteoglykany, spojující kolagenová vlákna, a fibronektiny a integriny, které jsou zabudovány v plasmatické mebráně. snad ovlivňuje i aktivitu genů.")
151
Extracelulární matrix
Proteoglykany kolagen fibronektiny integriny

152
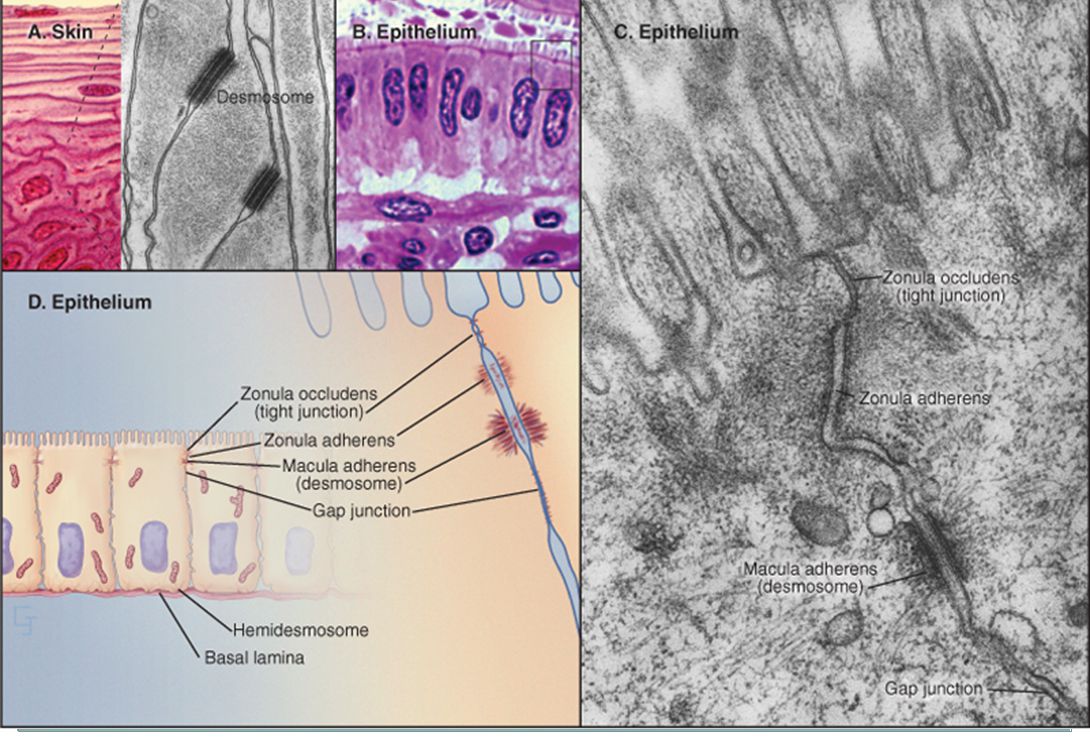
Buněčné spoje U rostlinných buněk plasmodesmata: může jimi pronikat voda a malé molekuly, ale také RNA a proteiny u živočišných buněk jsou: těsné spoje tvoří pásy např. v epitelu střeva, aby střevní obsah neprosakoval skrze buňky adhezní spoje desmosómy pevně spojují buňky k sobě mezerové spoje cytoplazmatické kanály mezi sousedními buňkami hemidesmosomy

153
Buněčné spoje živočišných buněk

155
Buněčné spoje živočišných buněk

156
Buněčné spoje živočišných buněk

157
Tight junctions EPITHELIAL TIGHT JUNCTIONS. A, Electron micrograph of a thin section of endothelial cells, showing a point of contact between the plasma membranes at a tight junction (arrow). B, Electron micrograph of a replica of a freeze-fractured cell. This method exposes proteins within the lipid bilayer and reveals strands aligned along the points of contact between the plasma membranes. C, Interpretive drawing, showing the strands at points of contact as rows of transmembrane proteins. (A, Courtesy of George Palade, University of California, San Diego. B, Courtesy of Don W. Fawcett, Harvard Medical School, Boston, Massachusetts.)
. B, Electron micrograph of a replica of a freeze-fractured cell. This method exposes proteins within the lipid bilayer and reveals strands aligned along the points of contact between the plasma membranes. C, Interpretive drawing, showing the strands at points of contact as rows of transmembrane proteins. (A, Courtesy of George Palade, University of California, San Diego. B, Courtesy of Don W. Fawcett, Harvard Medical School, Boston, Massachusetts.)")
158
Základní cytoplazma Tvořena převážně vodou a bílkovinami
Obsahuje rovněž kapénky tuků, krystaly …

159
krásný týden přeje Orko

Podobné prezentace

































.>")










>")





