Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Zuzana KAUEROVÁ 4.r. biochemie 2004/2005
PROTEAZOMY Zuzana KAUEROVÁ 4.r. biochemie 2004/2005

2
1. Když se řekne proteazom
Velké membránové multipodjednotkové komplexy, obsahující proteázy Hlavní úkol: degradace intracelulárních proteinů Výskyt: jádro a cytoplazma nejen u eukaryotních buněk, ale i u archebakterií (Thermoplasma acidofillum)
")
3
1.1. Historie Poprvé objeveny americkými vědci Sherwinem Wilkem a Marianem Orlowskim, členy farmakologické fakulty Mount Sinai School of Medicine v roce 1980, jako komplexy endopeptidáz, izolované ze srdce pacienta postiženého hypertrofií srdce

4
V roce 1988 vědec Arrigo prokázal, že tento systém proteáz, nazývaný dosud MPC (multikatalytický proteázový komplex) je identický s již objevenou částicí – prozomem (objeven německou skupinou Klause Scherrera) Prozom přejmenován na oficiální název proteazom

5
1.2. Hlavní funkce Odstraňují abnormální proteiny z buňky
Podílejí se na reakcích na stresové situace degradací proteinů, označených ubikvitinem (Ub) Jako součást Ub systému se zapojují do regulace buněčných procesů Kontrolují diferenciaci buněk (degradují transkripční faktory a metabolické enzymy) Velkou roli hrají v imunitním systému, kde vytváří peptidy MHC glykoproteinů I. třídy
 Jako součást Ub systému se zapojují do regulace buněčných procesů. Kontrolují diferenciaci buněk (degradují transkripční faktory a metabolické enzymy) Velkou roli hrají v imunitním systému, kde vytváří peptidy MHC glykoproteinů I. třídy.")
6
2. Proteinová degradace Nově objevený mechanizmus buněčné regulace
Hlavní výhoda proteolýzy – rychlost, umožňující buňce rapidní redukci hladiny určitých proteinů Nevratný děj – kompletní ztráta funkcí Základní požadavek: vysoká specifita

7
2.1. Ubikvitinace Nejobecnější způsob označování proteinů k degradaci v proteazomech Připojení řetězce ubikvitinu o 76 aminokyselinách Katalýza: - E1 aktivující enzymy - E2 konjugační enzymy - E3 vazebné enzymy (pouze v některých případech)
")
8
Proces degradace s ubikvitinem:
Ubikvitin je aktivován vytvořením thiosterové vazby s aktivačním enzymem E1. Ubikvitin je pak přenesen do aktivní cysteinové skupiny konjugačního enzymu E2, který ubikvitin naváže na aminoskupinu zbytku lyzinu substrátového proteinu. Pro tento proces je v některých případech nutná kooperace s enzymem E3. Opakovaná konjugace ubikvitinu s lyzinovými zbytky vede ke vzniku multiubikvitinových řetězců, které, navázané na substrátový protein, jsou rozeznány a degradovány v proteazomech. Multiubikvitinový řetězec se uvolní z komplexu a ubikvitin je recyklován.

11
2.2. Další způsoby označení proteinů k degradaci
„PEST sekvence“ = degradační signály (obsah v prolinu, kys. aspartové a glutamové, serinu a threoninu) Vymazání PEST sekvence => stabilita Neoznačené proteiny obsahují „destrukční boxy“ – silně zakonzervované aminokyseliny, navozující signál k degradaci
 Vymazání PEST sekvence => stabilita. Neoznačené proteiny obsahují „destrukční boxy – silně zakonzervované aminokyseliny, navozující signál k degradaci.")
12
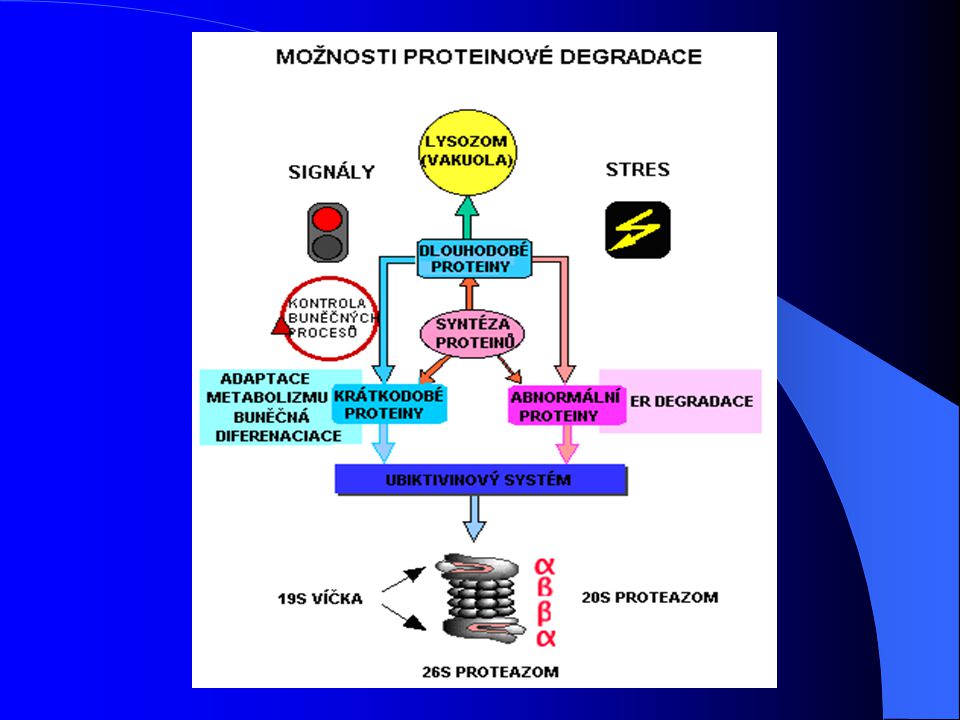
3. Struktura proteazomů 2 části:
20S proteazom jádrem větší částice 26S proteazomu Oba komplexy existují v dynamické rovnováze Hlavní rozdíl: 26S proteazom degraduje ubikvitinované proteiny, 20S proteazom ne

13
3.1. 20S proteazom 3.1.A Struktura
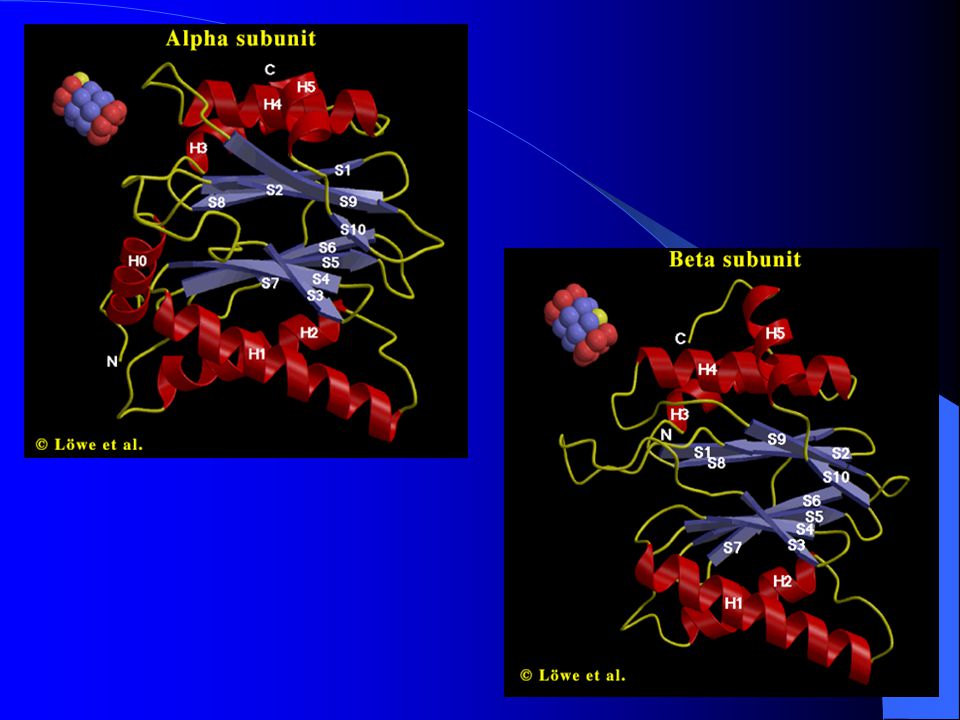
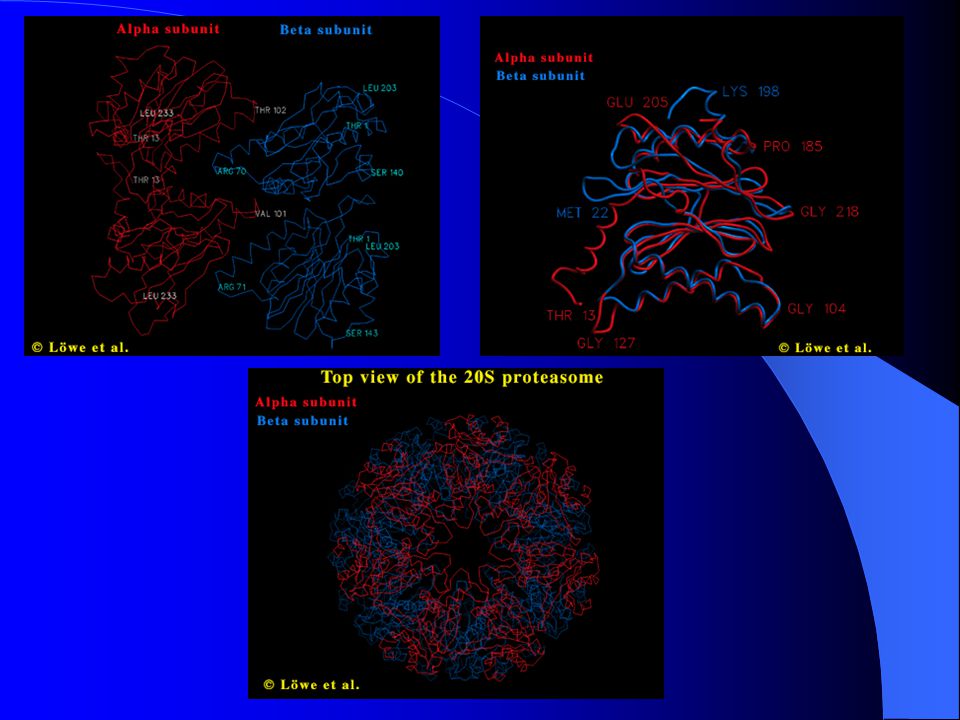
Často demonstrován na 26S proteazomu archebakterie Thermoplasma acidofillum – téměř identické s 20S proteazomy všech eukaryot Dutý cylindr, molekulová hmotnost 700 kDa Velké množství nízkomolekulárních podjednotek (25 – 35 kDa), 2 typy: α a β Uspořádány do 4 prstenců, každý o 7 podjednotkách
, 2 typy: α a β. Uspořádány do 4 prstenců, každý o 7 podjednotkách.")
15
Délka cylindru – 148 podjednotek (15 nm)
Výška cylindru – 113 podjednotek (11 nm) Vytváří kanál se 3 velkými výdutěmi (šířka 0,13 nm): 1. Dvě vnější výdutě na povrchu formovány α a β prstenci 2. Centrální výduť formována beta prstenci = centrum proteolytické aktivity Přístup do tohoto centra kontrolován 4 úzkými bránami (vnější brána, která je vytvářena α podjednotkami, propouští pouze otevírající se 113ti jednotkový průměr)
 Vytváří kanál se 3 velkými výdutěmi (šířka 0,13 nm): 1. Dvě vnější výdutě na povrchu formovány α a β prstenci. 2. Centrální výduť formována beta prstenci = centrum proteolytické aktivity. Přístup do tohoto centra kontrolován 4 úzkými bránami (vnější brána, která je vytvářena α podjednotkami, propouští pouze otevírající se 113ti jednotkový průměr)")
16
RRozdíly mezi α a β podjednotkami minimální
PPouze: α podjednotka vytváří strukturu α helixu napříč vrchní částí = součást výdutě, napomáhá transportu substrátových proteinů do centrálního kanálu NNezralá β podjednotka obsahuje N-terminální konec (Thr), který ji inaktivuje (funguje jako samostatná jednotka) => odstřihnutím => proteolyticky aktivní (ochrana před neadekvátní proteolýzou v cytoplazmě)
, který ji inaktivuje (funguje jako samostatná jednotka) => odstřihnutím => proteolyticky aktivní (ochrana před neadekvátní proteolýzou v cytoplazmě)")
18
Obr.: Proteazom na gelu a v elektronovém mikroskopu

20
3.1.B Funkce 20S proteazom je jádrem proteinové degradace
Reprezentuje unikátní typ proteáz – threonin proteázu Působí při degradaci řetězce A a B v inzulínu Oxiduje proteiny in vitro Veškeré procesy katalýzy probíhají uvnitř komplexu beta podjednotek

21
3.2. 26S proteazom 3.2.A Struktura
Na rozdíl od 20S proteazomů, které in vitro degradovaly pouze určité proteiny denaturací či oxidací za nepřítomnosti ATP, 26S proteazomy degradují ubikvitinové konjugáty v reakci závislé na ATP 26 proteazom = velký proteázový komplex, cca 1700kDa Struktura typická jako pro 20S proteazom, ale se dvěma přídavnými podstrukturami navázánými na oba konce = 19S části , tzv. „víčka“

22
3.2.B 19S struktura Asociována s 20S proteazomy na ATP závislým způsobem 19S víčka jsou prostředníkem mezi ubikvitinovým systémem a 26S proteazomem Složena z 20 různých podjednotek (25 – 110 kDa) Dosud přesně určeno pouze 6 podjednotek 19S struktury s ATPázovou aktivitou, které vytváří prstenec, sousedící s výdutí, ohraničenou α podjednotkami
 Dosud přesně určeno pouze 6 podjednotek 19S struktury s ATPázovou aktivitou, které vytváří prstenec, sousedící s výdutí, ohraničenou α podjednotkami.")
23
3.2.C Funkce 19S víček Slouží k zachycení a přitáhnutí označeného proteinu do proteazomu Hydrolýza ATP je spojena se vstupem označeného proteinu do proteolytického centra, přesný mechanizmus však není znám 19S struktura se nevyskytuje u archebakterií, typický pouze pro eukaryota

24
3.2.D Funkce 26S proteazomu Plní životně důležité funkce:
kontrola buněčného cyklu buněčná diferenciace regulace metabolických drah role ve stresových situacích odstraňování abnormálních proteinů

25
Kontrola buněčného cyklu
Cyklus dělení eukaryotických buněk kontrolován cyklin-dependentními protein kinázami (CDK) Vznik a zánik dílčích aktivovaných kinázových komplexů během různých fází buněčného cyklu regulován syntézou a proteolytickou degradací specifických cyklinů v proteazomech Inhibitory CDK jsou také kontrolovány proteolytickou destrukcí: cyklinová degradace potřebná pro zastavení aktivace CDK, proteolýza CDK inhibitorů => přepnutí CDK aktivity
 Vznik a zánik dílčích aktivovaných kinázových komplexů během různých fází buněčného cyklu regulován syntézou a proteolytickou degradací specifických cyklinů v proteazomech. Inhibitory CDK jsou také kontrolovány proteolytickou destrukcí: cyklinová degradace potřebná pro zastavení aktivace CDK, proteolýza CDK inhibitorů => přepnutí CDK aktivity.")
26
Stresová odpověď imunitního systému
Prostřednictvím NF-B = regulátor transkripce velkého množství genů, zapojujících se do imunitních a zánětlivých reakcí Tyto geny kódují zánětlivé a chemotaktické cytokiny, hematopoetické růstové faktory, adhezivní molekuly, protilátky, MHC glykoproteiny I. třídy a receptory pro cytokiny NF-B silně regulovaný a aktivovaný širokou směsicí většinou patogenních stimulů (např. viry, bakteriemi, energeticky bohatou radiací, oxidanty a zánětlivými cytokiny TNF- a IL-1)
")
27
p50 podjednotky (NF-B1) a p65 podjednotky (Re1A)
V aktivní formě je NF-B heterodimerní komplex, 2 podjednotky: p50 podjednotky (NF-B1) a p65 podjednotky (Re1A) => inaktivace dvěma různými mechanizmy: asociace proteinového inhibitoru I-B s p50-p65 komplexem syntéza NF-B1 - prekurzorového 105ti kilodaltonového proteinu, který se sice může kompletovat s p65 podjednotkou, ale nevytváří aktivní komplex.
 a p65 podjednotky (Re1A) => inaktivace dvěma různými mechanizmy: asociace proteinového inhibitoru I-B s p50-p65 komplexem. syntéza NF-B1 - prekurzorového 105ti kilodaltonového proteinu, který se sice může kompletovat s p65 podjednotkou, ale nevytváří aktivní komplex.")
28
Proteazomy jsou zodpovědné za aktivaci NF-B degradací proteinu I-B a převedením 105ti kilodaltonového prekurzoru NF-B na p50 aktivní formu Proteazomy tedy nejsou pouze zodpovědné za kompletní degradaci proteinů, ale také jsou nezbytné pro aktivaci proteinů při procesu aktivace prekurzorů.

29
Antigenní prezentace Proteazomy mohou syntetizovat krátké peptidové fragmenty => prezentovány na povrchu lymfocytů v MHC komplexu glykoproteinů Hrají důležitou roli ve schopnosti buněk reagovat na stresové situace V buňkách imunitního systému jsou některé β podjednotky nahrazeny gamma-interferon indikačními homology, které zvyšují efektivnost degradace v proteazomech (struktura téměř stejná)
")
31
4. ZÁVĚR Proteazomy mohou být považovány za funkčně propracované doplňky ribozomů. Regulace hladiny proteinů destrukcí prostřednictvím proteazomů je nezbytný nástroj buňky. Je zřejmé, že několik dodnes objevených substrátů v proteazomech reprezentují pouze špičku ledovce. Přesto jsou však velkou výzvou k objevu všech buněčných procesů, které se v proteazomech odehrávají, stejně tak jako detailní mechanizmy, zdůrazňující tyto selektivní procesy.

Podobné prezentace



>")










>")
>")






