Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Přednášky z lékařské biofyziky Biofyzikální ústav Lékařské fakulty Masarykovy univerzity, Brno

2
Přístrojové metody molekulární biofyziky
Přednášky z lékařské biofyziky Biofyzikální ústav Lékařské fakulty Masarykovy univerzity, Brno Přístrojové metody molekulární biofyziky

3
Obsah přednášky Biomolekulární vědy mají klíčový význam pro molekulární medicínu. Budeme se zabývat zařízeními pro studium struktury, měření koncentrace (in-vitro i in-vivo), a pro studium vlastností membrán. Nejdůležitější zařízení založená na interakci elektromagnetického záření s makromolekulami VIS, UV a IR spektrofotometry Ramanovy spektrometry Zařízení pro měření cirkulárního dichroismu Zařízení pro rentgenstrukturní analýzu Zařízení založená na jiných vlastnostech biomolekul (např. mechanických a elektrických) Elektroforéza Zařízení pro měření membránových potenciálů a koncentrace iontů v buňkách
, a pro studium vlastností membrán. Nejdůležitější zařízení založená na interakci elektromagnetického záření s makromolekulami. VIS, UV a IR spektrofotometry. Ramanovy spektrometry. Zařízení pro měření cirkulárního dichroismu. Zařízení pro rentgenstrukturní analýzu. Zařízení založená na jiných vlastnostech biomolekul (např. mechanických a elektrických) Elektroforéza. Zařízení pro měření membránových potenciálů a koncentrace iontů v buňkách.")
4
Nebudeme se zabývat…. Zařízeními pro měření
Osmolární koncentrace (měření je založeno na kryoskopii), Rychlosti difuze Viskozity (praktická cvičení) Přístroji pro stanovení sekundární a terciární struktury bílkovin a nukleových kyselin pracujícími na elektrochemickém základě (je studována interakce makromolekul s elektrodami) Nukleární magnetickou rezonancí (umožňuje např. zjistit, jak je v molekule chemicky vázaný vodík – zmíněno v přednášce o MRI) Elektronovou spinovou rezonancí, Centrifugami (jiná přednáška) atd.
, Rychlosti difuze. Viskozity (praktická cvičení) Přístroji pro stanovení sekundární a terciární struktury bílkovin a nukleových kyselin pracujícími na elektrochemickém základě (je studována interakce makromolekul s elektrodami) Nukleární magnetickou rezonancí (umožňuje např. zjistit, jak je v molekule chemicky vázaný vodík – zmíněno v přednášce o MRI) Elektronovou spinovou rezonancí, Centrifugami (jiná přednáška) atd.")
5
Biofyzika a biomolekulární výzkum
Tento výzkum je orientován zejména na strukturální studie, které umožňují porozumět např.: Specifičnosti enzymatických a imunologických reakcí Účinkům některých léků (např. cytostatik) na molekulární úrovni. Mechanismům pasivního i aktivního transportu Buněčnému pohybu ……………..
 na molekulární úrovni. Mechanismům pasivního i aktivního transportu. Buněčnému pohybu. ……………..")
6
Zařízení založená na interakci elektromagnetického záření s makromolekulami

7
Druhy spektrofotometrů
Spektrofotometry jsou laboratorní přístroje používané studium látek absorbujících nebo emitujících infračervené, viditelné nebo ultrafialové světlo, včetně studia jejich chemické struktury. Absorpční spektrofotometry: založeny na spektrální závislosti absorpce světla. Emisní spektrofotometry: Zdrojem světla je sama analyzovaná látka, jež je injektována nebo rozprašována do bezbarvého plamene. Emitované světlo prochází optickým hranolem nebo mřížkou, takže můžeme získat celé emisní spektrum. Frekvence přítomné ve spektru umožňují identifikovat např. přítomné ionty. Spektrofluorimetry: emise světla je vyvolána světlem o vlnové délce kratší než je vlnová délka světla emitovaného.

8
Absorpční spektrofotometry: Lambertův-Beerův zákon
Absorpční spektrofotometrie je založena na absorpci světla při průchodu vrstvou roztoku látky pohlcující světlo. Jeho koncentrace může být zjištěna pomocí Lambertova-Beerova zákona: I = I0.10-ecx c je koncentrace rozpuštěné látky, x tloušťka vrstvy, I0 původní intenzita světla, I je intenzita světla po průchodu vrstvou. Konstanta e (epsilon, absorpční nebo extinkční koeficient) závisí na vlnové délce světla, na rozpuštěné látce i rozpouštědle. Její hodnoty pro běžné chemické sloučeniny lze nalézt v tabulkách. Tyto hodnoty jsou vždy udávány pro určitou vlnovou délku (obvykle absorpční maximum). Číselné hodnoty tohoto koeficientu závisejí na tom, jak je vyjadřována koncentrace rozpuštěné látky. Když použijeme mol.l-1, hovoříme o molárním absorpčním koeficientu.
 závisí na vlnové délce světla, na rozpuštěné látce i rozpouštědle. Její hodnoty pro běžné chemické sloučeniny lze nalézt v tabulkách. Tyto hodnoty jsou vždy udávány pro určitou vlnovou délku (obvykle absorpční maximum). Číselné hodnoty tohoto koeficientu závisejí na tom, jak je vyjadřována koncentrace rozpuštěné látky. Když použijeme mol.l-1, hovoříme o molárním absorpčním koeficientu.")
9
Poměr intenzit světla prošlého a dopadajícího se nazývá transmitance (dříve transparence). Dekadický logaritmus převrácené hodnoty transmitance se nazývá absorbance A. S ohledem na L.-B. zákon je tedy absorbance přímo úměrná koncentraci rozpuštěné látky a tloušťce absorbující vrstvy roztoku. A = e.c.x

10
Druhy absorpčních spektrofotometrů
Podle konstrukce rozdělujeme spektrofotometry na jednopaprskové a dvoupaprskové. U jednopaprskových spektrofotometrů jeden svazek světla prochází nejdříve srovnávacím a pak měřeným vzorkem (kyvety obsahující roztoky musí být pohyblivé). U dvoupaprskových spektrofotometrů jeden svazek světla prochází měřeným vzorkem a druhý srovnávacím vzorkem (blankem). Dvoupaprskové přístroje umožňují podstatně rychlejší měření, avšak jsou dražší. U jednoduchých přístrojů je nastavování vlnové délky světla ruční. U pokročilejších přístrojů se toto nastavování děje automaticky, což umožňuje přímo získávat absorpční křivky, tj. grafy závislostí absorbance na vlnové délce světla.
. U dvoupaprskových spektrofotometrů jeden svazek světla prochází měřeným vzorkem a druhý srovnávacím vzorkem (blankem). Dvoupaprskové přístroje umožňují podstatně rychlejší měření, avšak jsou dražší. U jednoduchých přístrojů je nastavování vlnové délky světla ruční. U pokročilejších přístrojů se toto nastavování děje automaticky, což umožňuje přímo získávat absorpční křivky, tj. grafy závislostí absorbance na vlnové délce světla.")
11
Jednopaprskový spektrofotometr
Zdrojem světla (1) je žárovka s wolframovým vláknem. Její polychromatické světlo prochází kondenzorem (2) a odráží se od zrcadla (3) na vstupní štěrbinu (4) monochromátoru (části 4 až 8, plus 12). Světlo je soustřeďováno čočkou (5) na odrazovou optickou mřížku (6), která tvoří barevné spektrum. Téměř monochromatické světlo je promítáno objektivem (7) na výstupní štěrbinu (8) monochromátoru.
 je žárovka s wolframovým vláknem. Její polychromatické světlo prochází kondenzorem (2) a odráží se od zrcadla (3) na vstupní štěrbinu (4) monochromátoru (části 4 až 8, plus 12). Světlo je soustřeďováno čočkou (5) na odrazovou optickou mřížku (6), která tvoří barevné spektrum. Téměř monochromatické světlo je promítáno objektivem (7) na výstupní štěrbinu (8) monochromátoru.")
12
Jednopaprskový spektrofotometr
S mřížkou lze otáčet pomocí ovladače vlnových délek (12), čímž se zaměřuje světlo o určité vlnové délce na výstupní štěrbinu. Svazek světla pak prochází kyvetou (9) se vzorkem. Intenzita prošlého světla je měřena fotodetektorem (10, 11). Jeho signál je zesilován zesilovačem (13). Hodnota absorbance je zobrazena na displeji (14). Intenzita světla prošlého srovnávacím roztokem je vždy srovnávána s intenzitou téhož svazku světla prošlého měřeným vzorkem.
, čímž se zaměřuje světlo o určité vlnové délce na výstupní štěrbinu. Svazek světla pak prochází kyvetou (9) se vzorkem. Intenzita prošlého světla je měřena fotodetektorem (10, 11). Jeho signál je zesilován zesilovačem (13). Hodnota absorbance je zobrazena na displeji (14). Intenzita světla prošlého srovnávacím roztokem je vždy srovnávána s intenzitou téhož svazku světla prošlého měřeným vzorkem.")
13
Moderní UV/VIS/NIR spektrofotometr
NIR = near infrared = blízká infračervená oblast Světlo jedné vybrané vlnové délky nebo celé prošlé spektrum může být měřeno

14
Absorpční UV spektrofotometrie
Ultrafialové (UV) světlo je absorbováno různými sloučeninami, zejména těmi, které mají konjugované dvojné vazby. Jak bílkoviny, tak nukleové kyseliny silně absorbují UV světlo, což lze využít pro jejich zkoumání. Aminokyseliny tryptofan a tyrosin mají absorpční maxima při přibližně 280 nm. Fenylalanin při 255 nm. Nukleotidy (dusíkaté báze) mají absorpční maxima v oblasti nm. Chromofory – jejich absorpční vlastnosti se mění podle chemického složení prostředí.
 světlo je absorbováno různými sloučeninami, zejména těmi, které mají konjugované dvojné vazby. Jak bílkoviny, tak nukleové kyseliny silně absorbují UV světlo, což lze využít pro jejich zkoumání. Aminokyseliny tryptofan a tyrosin mají absorpční maxima při přibližně 280 nm. Fenylalanin při 255 nm. Nukleotidy (dusíkaté báze) mají absorpční maxima v oblasti nm. Chromofory – jejich absorpční vlastnosti se mění podle chemického složení prostředí.")
15
Absorpční spektra aminokyselin
According:

16
Hypochromní efekt (HE)
Absorpce světla je ovlivňována dipólovými momenty chemických vazeb, které interagují s fotony. Stochasticky (náhodně) orientované dipólové momenty (denaturovaná bílkovina) absorbují světlo lépe než ve stavu uspořádaném (šroubovice). U bílkovin je HE způsoben peptidovými vazbami, které mají UV absorpční maximum kolem 190 nm. Dvoušroubovice DNA absorbuje UV světlo lépe než DNA denaturovaná (neuspořádaná). Helicita – relativní zastoupení uspořádaných částí makromolekuly
 orientované dipólové momenty (denaturovaná bílkovina) absorbují světlo lépe než ve stavu uspořádaném (šroubovice). U bílkovin je HE způsoben peptidovými vazbami, které mají UV absorpční maximum kolem 190 nm. Dvoušroubovice DNA absorbuje UV světlo lépe než DNA denaturovaná (neuspořádaná). Helicita – relativní zastoupení uspořádaných částí makromolekuly.")
17
Hypochromní efekt u kys. polyglutamové
Hypochromní efekt u kys. polyglutamové. Při pH 7 tento polypeptid tvoří stochastické (neuspořádané) klubko (1), při pH 4 získává šroubovicovou strukturu (2). Absorpční maximum peptidových vazeb je snížené vlivem jejich prostorového uspořádání. e je molární absorpční koeficient a l je vlnová délka UV světla. [dle: Kalous a Pavlíček, 1980]
 klubko (1), při pH 4 získává šroubovicovou strukturu (2). Absorpční maximum peptidových vazeb je snížené vlivem jejich prostorového uspořádání. e je molární absorpční koeficient a l je vlnová délka UV světla. [dle: Kalous a Pavlíček, 1980]")
18
IR spektrofotometrie Infračervené záření (IR) působí na rotační a vibrační stavy molekul. Složité molekuly mohou vibrovat nebo rotovat mnoha různými způsoby (módy). Různé chemické skupiny (-CH3, -OH, -COOH, -NH2 atd.) mají specifické vibrační a rotační frekvence, a proto absorbují IR světlo o specifických vlnových délkách. Z tohoto důvodu mají infračervená absorpční spektra mnoho maxim. Změna chemické struktury se projevuje jako změna polohy těchto maxim ve spektru.
 působí na rotační a vibrační stavy molekul. Složité molekuly mohou vibrovat nebo rotovat mnoha různými způsoby (módy). Různé chemické skupiny (-CH3, -OH, -COOH, -NH2 atd.) mají specifické vibrační a rotační frekvence, a proto absorbují IR světlo o specifických vlnových délkách. Z tohoto důvodu mají infračervená absorpční spektra mnoho maxim. Změna chemické struktury se projevuje jako změna polohy těchto maxim ve spektru.")
19
Infračervené spektrum hexanu vyjádřené jako závislost transmitance na vlnočtu

20
Ramanova spektrometrie
Rayleighův rozptyl světla. Nastává interakce fotonů s molekulami, jež se projevuje jen velmi malou nebo žádnou změnou vlnové délky. Intenzita rozptýleného světla závisí na molekulové hmotnosti a také na úhlu rozptylu, což lze využít pro odhad tvaru makromolekul. Ramanova spektrometrie. Při rozptylu fotonů nastává malá změna (posun) vlnové délky, způsobená malým poklesem nebo zvýšením energie rozptýlených fotonů během přechodu z původního do změněného vibračního nebo rotačního stavu interagující molekuly. Tyto stavy se mohou měnit v důsledku strukturálních změn molekul. Proto změny v Ramanových spektrech (intenzita signálu v závislosti na posunu vlnové délky nebo odpovídajícímu vlnočtu) odrážejí konformační změny molekul.
 vlnové délky, způsobená malým poklesem nebo zvýšením energie rozptýlených fotonů během přechodu z původního do změněného vibračního nebo rotačního stavu interagující molekuly. Tyto stavy se mohou měnit v důsledku strukturálních změn molekul. Proto změny v Ramanových spektrech (intenzita signálu v závislosti na posunu vlnové délky nebo odpovídajícímu vlnočtu) odrážejí konformační změny molekul.")
21
Ramanova spektrometrie
Ramanovo spektrum polytenního chromosomu pakomára Chironomus. Při zvolených vlnočtech lze uskutečnit ramanovskou mikroskopii. Vybuzeno laserovým světlem o vlnové délce nm. According to:

22
Mikrofotografie v normálním bílém světle
(chromozom Chironomus Thummi Thummi) Konfokální ramanovská mikrofotografie zobrazující páteř DNA (vibrace při 1094 cm-1) Konfokální ramanovská mikrofotografie zobrazující alifatické řetězce v bílkovinách při 1449 cm-1 podle: volume2/edition3/section4.htm
 Konfokální ramanovská mikrofotografie zobrazující páteř DNA (vibrace při 1094 cm-1) Konfokální ramanovská mikrofotografie zobrazující alifatické řetězce v bílkovinách při 1449 cm-1. podle: volume2/edition3/section4.htm.")
23
Optická rotační disperze
Metodou optické rotační disperze (ORD) měříme závislost optické aktivity na vlnové délce světla. Tato metoda však byla nahrazena citlivější metodou cirkulárního dichroismu (CD), která poskytuje podobné informace.
 měříme závislost optické aktivity na vlnové délce světla. Tato metoda však byla nahrazena citlivější metodou cirkulárního dichroismu (CD), která poskytuje podobné informace.")
24
Cirkulární dichroismus (CD) - nepovinné
Měření optické aktivity (schopnosti stáčet rovinu polarizovaného světla). Konformační změny molekul mohou být sledovány jako změny optické aktivity při použití speciálního polarimetru. U metody CD srovnáváme absorbance levotočivě a pravotočivě cirkulárně polarizovaného světla, jehož vlnová délka je blízká absorpčnímu maximu bílkoviny. CD lze využít též pro studium struktury nukleových kyselin. Obrázek ukazuje změny elipticity syntetického polypeptidu, obsahujícího dlouhé sekvence poly-glu, po přídavku trifluoroethanolu (TFE), který zvyšuje podíl a-šroubovice.
. Konformační změny molekul mohou být sledovány jako změny optické aktivity při použití speciálního polarimetru. U metody CD srovnáváme absorbance levotočivě a pravotočivě cirkulárně polarizovaného světla, jehož vlnová délka je blízká absorpčnímu maximu bílkoviny. CD lze využít též pro studium struktury nukleových kyselin. Obrázek ukazuje změny elipticity syntetického polypeptidu, obsahujícího dlouhé sekvence poly-glu, po přídavku trifluoroethanolu (TFE), který zvyšuje podíl a-šroubovice.")
25
Rentgenstrukturní analýza
Krystalová mřížka působí na rentgenové záření jako optická mřížka na viditelné světlo. Nastávají ohybové jevy a na stínítku se objevuje difrakční obrazec. Tyto obrazce mohou být matematicky analyzovány, aby se získala informace o rozložení elektronů v molekulách tvořících krystal.

26
Mapa elektronové hustoty organické sloučeniny vypočtená z rentgenového krystalogramu

27
Krystalogram B-DNA získaný v r. 1952 Rosalindou E
Krystalogram B-DNA získaný v r Rosalindou E. Franklinovou, na jehož základě Watson a Crick navrhli dvoušroubovicový model struktury DNA. F C W

28
Metody založené na měření mechanických a elektrických vlastností makromolekul
Velikost a tvar makromolekul můžeme studovat na základě měření: Osmotického tlaku (velikost, přednáška ″Termodynamika a život″) Difuzního koeficientu (velikost, přednáška ″Termodynamika a život″) Viskozity (tvar, praktická cvičení) Sedimentace (velikost, přednáška ″Zařízení pro elektrochemickou analýzu. Pomocné laboratorní přístroje ″ Dále můžeme použít: Elektronovou mikroskopii a metody příbuzné, včetně AFM (velikost a tvar, přednáška ″Mikroskopie″) Chromatografii – molekulárně síťový efekt u gelové permeační chromatografie (chemie) Elektroforézu (konec této části přednášky)
 Difuzního koeficientu (velikost, přednáška ″Termodynamika a život″) Viskozity (tvar, praktická cvičení) Sedimentace (velikost, přednáška ″Zařízení pro elektrochemickou analýzu. Pomocné laboratorní přístroje ″ Dále můžeme použít: Elektronovou mikroskopii a metody příbuzné, včetně AFM (velikost a tvar, přednáška ″Mikroskopie″) Chromatografii – molekulárně síťový efekt u gelové permeační chromatografie (chemie) Elektroforézu (konec této části přednášky)")
29
Zařízení pro elektroforézu
Zdroj napětí Jamky v gelu pro vzorky Gelová plotna Látkový knot Roztok elektrolytu

30
Elektrochemické vlastnosti koloidů
Koloidy jsou roztoky, které obsahují částice o velikosti 10 – 1000 nm. Některé molekulární i micelární koloidy jsou polyelektrolyty s amfoterními vlastnostmi. Tyto amfolyty se chovají buď jako zásady nebo kyseliny v závislosti na pH prostředí. U bílkovin se mění počet skupin –NH3+ a –COO-.

31
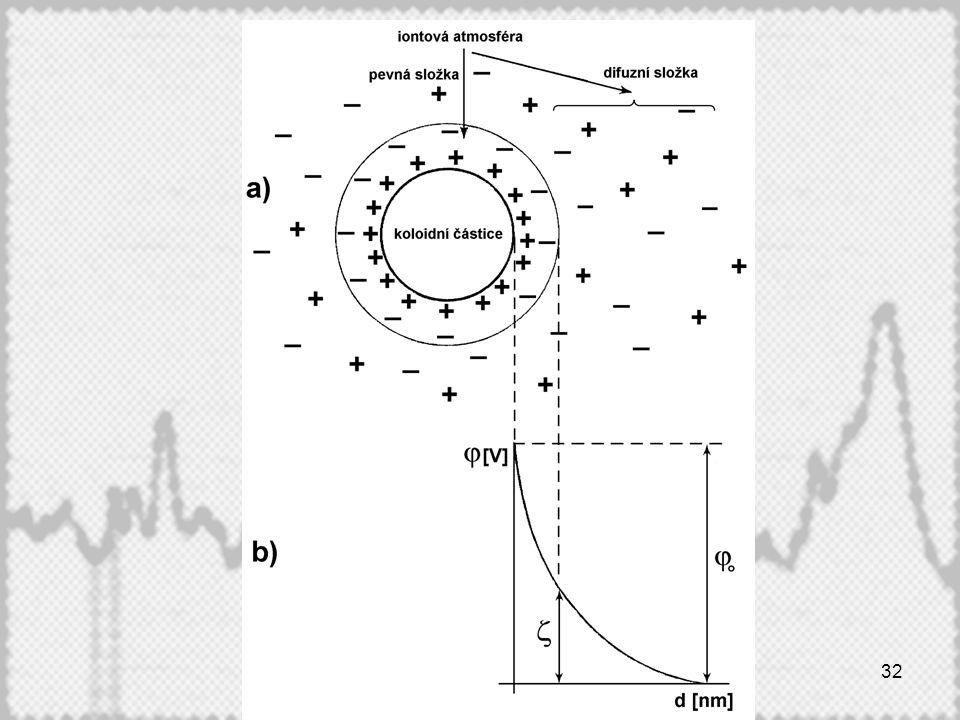
Vznik elektrické dvojvrstvy na povrchu koloidní částice
Dva mechanismy: Adsorpce iontů (i u hydrofobních koloidů) Elektrolytická disociace (převažuje u hydrofilních koloidů) Dvojvrstva na povrchu částice se liší v koncentrovaných a zředěných elektrolytech. U zředěných elektrolytů můžeme v celé iontové atmosféře částice rozlišit stabilní, difuzní a elektroneutrální oblast. Elektrokinetický potenciál – z (zeta)-potenciál
 Elektrolytická disociace (převažuje u hydrofilních koloidů) Dvojvrstva na povrchu částice se liší v koncentrovaných a zředěných elektrolytech. U zředěných elektrolytů můžeme v celé iontové atmosféře částice rozlišit stabilní, difuzní a elektroneutrální oblast. Elektrokinetický potenciál – z (zeta)-potenciál.")
33
Elektroforéza F = 6.p.r.h.v F = z.e.E
Elektroforéza – pohyb nabitých molekul v elektrickém poli. Při rovnoměrném přímočarém pohybu sférické částice o poloměru r, je elektrostatická síla působící na částici v rovnováze se silou tření, jež je dána viskozitou. Sílu tření lze vypočítat dle Stokesova vzorce: F = 6.p.r.h.v kde v je rychlost částice a h je dynamická viskozita prostředí. Elektrické pole působí na částici silou: F = z.e.E kde z je počet elementárních nábojů nesených částicí, e je elementární náboj (1, C) a E [V.m-1] je intenzita elektrického pole v daném místě. Rychlost částice je pak v důsledku rovnosti obou sil:
 a E [V.m-1] je intenzita elektrického pole v daném místě. Rychlost částice je pak v důsledku rovnosti obou sil:")
34
Elektroforetická pohyblivost
Elektroforetická pohyblivost u nezávisí na intenzitě elektrického pole. Je definována jako podíl rychlosti částice a intenzity elektrického pole. Platí: Poznámka. Elektroforéza s dodecylsulfátem sodným. Tato sloučenina, která nese jeden negativní elementární náboj, se váže definovaným způsobem k bílkovinám a eliminuje jejich vlastní elektrický náboj. Molekuly bílkovin se pak pohybují s různou rychlostí jen proto, že mají různou velikost (poloměr).
.")
35
Měření membránových potenciálů
Membránové potenciály se měří s pomocí skleněných mikroelektrod, tj. skleněných kapilár s velmi jemnou úzkou špičkou. Průměr otvoru na konci špičky musí být menší než 1 mm, aby nedošlo při zavádění do buňky k jejímu významnému poškození. Vnitřní prostor špičky kapiláry je naplněn roztokem KCl o koncentraci 3 mol.l-1. Jako elektroda srovnávací se používá elektroda stříbrochloridová umístěná do mimobuněčného prostoru. Pro skleněné mikroelektrody je charakteristický vysoký vnitřní odpor (kolem 10 MW), takže potřebujeme pro měření vysoce kvalitní zesilovače, abychom zamezili zkreslení měřeného napětí.
, takže potřebujeme pro měření vysoce kvalitní zesilovače, abychom zamezili zkreslení měřeného napětí.")
36
Experimentální uspořádání pro měření membránových potenciálů kapilárními mikroelektrodami
Pomocí skleněných mikroelektrod lze také měřit jiné elektrochemické parametry buněk a membrán, např. koncentraci některých iontů. Mohou být připraveny jako elektrody iontově selektivní pro Na+, K+, Ca2+, H+ …

37
Metoda patch-clamp („terčíkový zámek“)
Tupá skleněná mikroelektroda se přiloží k povrchu buňky nebo k části biologické či umělé membrány. Otvor na konci mikroelektrody je zcela uzavřen „terčíkem“ membrány a měřená elektrická napětí nebo proudy se proto týkají jen malého okrsku membrány, v němž se nalézá jen malý počet iontových kanálů. Některé iontové kanály mohou být předem uzavřeny nebo otevřeny, náplň mikroelektrody může obsahovat ligandy, schopné interagovat s iontovými kanály, a všeobecně jakékoliv látky, jež mohou ovlivňovat funkci membrány. Tato metoda umožňuje studium aktivity jednotlivých iontových kanálů nebo jejich malých skupin.

38
Autor: Vojtěch Mornstein Obsahová spolupráce: Viktor Brabec, Carmel J
Autor: Vojtěch Mornstein Obsahová spolupráce: Viktor Brabec, Carmel J. Caruana Grafika: Lucie Mornsteinová Poslední revize: březen 2012

Podobné prezentace






 generátor>")






>")
 REZONANCE>")




