Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
PLAZMATICKÉ LIPIDY A TRANSPORT LIPIDŮ
Podpora zdraví FTK UP Olomouc

2
VOLNÉ MASTNÉ KYSELINY (VMK, FFA)
se vážou na albumin. CHOLESTEROL (CH), TRIGLYCERIDY (TG) A FOSFOLIPIDY jsou transportovány ve formě lipoproteinových komplexů.
, TRIGLYCERIDY (TG) A FOSFOLIPIDY. jsou transportovány ve formě lipoproteinových komplexů.")
3
HLAVNÍ LIPOPROTEINY chylomikrony chylomikronové zbytky
lipoproteiny o velmi nízké hustotě (VLDL) lipoproteiny a stření hustotě (IDL) lipoproteiny o nízké hustotě (LDL) lipoproteiny o vysoké hustotě (HDL)
 lipoproteiny a stření hustotě (IDL) lipoproteiny o nízké hustotě (LDL) lipoproteiny o vysoké hustotě (HDL)")
4
VELIKOST Chylomikrony 75 - 1000 Chylomikronové zbytky 30 - 80
VLDL IDL LDL HDL ,5 - 10

5
PROTEIN Chylomikrony 2 Chylomikronové zbytky .. VLDL 8 IDL 10 LDL 20
HDL

6
VOLNÝ CH Chylomikrony 2 Chylomikronové zbytky .. VLDL 4 IDL 5 LDL 7
HDL

7
ESTERY CH Chylomikrony 3 Chylomikronové zbytky .. VLDL 16 IDL 25
HDL

8
TG Chylomikrony 90 Chylomikronové zbytky VLDL IDL LDL HDL

9
FOSFOLIPIDY Chylomikrony 3 Chylomikronové zbytky .. VLDL 17 IDL 20
HDL

10
PŮVOD Chylomikrony střevo Chylomikronové zbytky kapiláry
VLDL játra a střevo IDL VLDL LDL IDL HDL játra a střevo

11
Hustota lipoproteinů (rychlost s jakou sedimentují při ultracentrifugaci) má obrácený poměr k obsahu lipidů VLDL CH = 4, estery CH = 16, TG = 55 HDL CH = 4, estery CH = 16, TG = 5 Čím větší hustota, tím menší obsah lipidů

12
SLOŽENÍ LIPOPROTEINŮ TG + estery CH proteiny fosfolipidy
jádro TG + estery CH proteiny fosfolipidy

13
PROTEINY= APOPROTEINY (Apo)
Apo E (chylomikrony, chylomikronové zbytky, VLDL, IDL, HDL) Apo C (chylomikrony, VLDL) - umožňuje, aby zůstaly v kapilárách Apo B -48 (chylomikrony, chylomikronové zbytky) = exogenní transportní systém Apo B-100 (VLDL, IDL, LDL) = endogenní transportní systém
 Apo C (chylomikrony, VLDL) - umožňuje, aby zůstaly v kapilárách. Apo B -48 (chylomikrony, chylomikronové zbytky) = exogenní transportní systém. Apo B-100 (VLDL, IDL, LDL) = endogenní transportní systém.")
14
CHYLOMIKRONY Obrovské lipoproteinové komplexy
Do krve lymfatickými cestami Tvoří se ve střevní sliznici během resorpce produktů štěpení tuků (po jídle plazma mléčný vzhled) Chylomikrony jsou odstraňovány z krve působením lipoproteinové lipázy (LPL), která je umístěna na povrchu endotelu kapilár LPL štěpí TG na FFA a glycerol FFA a glycerol vstupují do adipocytů a reesterifikují se zpět na tuky
 Chylomikrony jsou odstraňovány z krve působením lipoproteinové lipázy (LPL), která je umístěna na povrchu endotelu kapilár. LPL štěpí TG na FFA a glycerol. FFA a glycerol vstupují do adipocytů a reesterifikují se zpět na tuky.")
15
LPL Odstraňuje TG i z VLDL Je aktivována apo C-II (jeden z apo C) Čili
Slupka chylomikronů obsahuje apo C-II Když se chylomikrony dostanou k endotelu cév, aktivuje apo C-II LPL LPL rozloží TG v jádru chylomikronů na FFA a glycerol

16
CHYLOMIKRONOVÉ ZBYTKY
Chylomikrony - TG = lipoproteiny bohaté na CH Do jater, kde se váží na receptory pro chylomikronové zbytky a na LDL receptory Odbourávají se v hepatocytech. CHYLOMIKRONOVÉ ZBYTKY A CHYLOMIKRONY VYTVÁŘEJÍ TRANSPORTNÍ SYSTÉM PRO STRÁVENÉ EXOGENNÍ LIPIDY

17
EXOGENNÍ DRÁHA Žlučové kyselin CH z potravy + CH STŘEVO JÁTRA C B-48
Chylomikrony Chylomikronové zbytky LPL KAPILÁRA

18
VLDL IDL LDL HDL CHYLOMIKRONOVÉ ZBYTKY A CHYLOMIKRONY
VYTVÁŘEJÍ EXOGENNÍ DRÁHU ENDOGENNÍ DRÁHA? VLDL IDL LDL HDL TRANSPORTUJÍ TG a CH PO CELÉM TĚLE

19
VLDL vznikají v játrech
transportují TG vytvořené z mastných kyselin a sacharidů z jater do mimojaterních tkání jsou-li TG velkou měrou z VLDL odstraněny pomocí LPL, stávají se z nich IDL VLDL s menším obsahem TG Co to jsou IDL?

20
ENDOGENNÍ DRÁHA JÁTRA VLDL IDL E C B-100 E B-100 LPL KAPILÁRA

21
IDL poskytují fosfolipidy
působením lecitincholesterolacyltransferázy (LCAT) vychytávají estery CH, které vznikají z CH v HDL.
 vychytávají estery CH, které vznikají z CH v HDL.")
22
ENDOGENNÍ DRÁHA JÁTRA B-100 VLDL IDL HDL E C B-100 E LPL KAPILÁRA
PLASMA LCAT

23
HDL2 HDL IDL HDL3 LCAT estery CH Variabilní množství Poměrně
konstantní množství

24
ENDOGENNÍ DRÁHA JÁTRA B-100 VLDL IDL HDL E C B-100 E LPL KAPILÁRA
PLASMA LCAT

25
V játrech se z IDL odštěpují další TG a proteiny a vznikají
LDL ztrácejí apo E B-100 v nich zůstává

26
ENDOGENNÍ DRÁHA B-100 LDL JÁTRA B-100 VLDL IDL HDL E C B-100 E LPL
KAPILÁRA PLASMA LCAT

27
LDL poskytuje tkáním CH
CH je základní součástí buněčných membrán z CH vznikají steroidní hormony LDL jsou v játrech a mimojaterních tkáních vychytávány receptory, které rozpoznají apo B-100 složky LDL LDL-receptory vytvářejí také vazbu s apo E, ne s apo B-48

28
ENDOGENNÍ DRÁHA Receptor LDL B-100 LDL JÁTRA MIMOJATERNÍ TKÁŇ B-100
VLDL IDL HDL E C B-100 E LPL KAPILÁRA PLASMA LCAT

29
Lidský receptor pro LDL
Je mozaikový protein, skládající se z pěti oblastí. Receptory zprostředkovávají endocytózu v povlečených jamkách buněk.

30
Endocytóza zprostředkovaná receptory
Každá z povlečených jamek se odškrcuje a vytváří potažený vezikul. Tím se uvolní receptor pro LDL a zabudovává se znovu do buněčné membrány.

31
LDL receptor endozom Vazba LDL Internalizace

32
Endocytóza zprostředkovaná receptory
Každá z povlečených jamek se odškrcuje a vytváří potažený vezikul. Tím se uvolní receptor pro LDL a zabudovává se znovu do buněčné membrány. Endozom splyne s lysozomem a cholesterol, který vzniká působením kyselé lipázy, může doplnit potřeby buňky.

33
LDL receptor CH lipáza Vazba LDL Internalizace Lysozomální hydrolýza

34
Cholesterol zpětnou vazbou inhibuje svou vlastní syntézu.
Je-li příjem CH vysoký, sníží se syntéza CH v játrech a naopak Stimuluje esterifikaci přebytečného CH, který se uvolňuje a inhibuje syntézu nových LDL receptorů.

35
LDL receptor Vazba LDL

36
LDL receptor CH Vazba LDL

37
VŠECHNY REAKCE ZAJIŠŤUJÍ ZPĚTNOU KONTROLOU MNOŽSTVÍ CH V BUŇCE.

38
LDL receptor CH CH

39
LDL receptor CH CH

40
LDL receptor HDL

41
Syntetizovaný v játrech a ve střevě
LDL receptor Syntetizovaný v játrech a ve střevě HDL

42
Některé HDL obsahují apo E a navazují se na LDL receptory v buňkách a tak přenášejí CH z jedné buňky na druhou. Navíc přes LCAT poskytují HDL estery CH IDL a poté buď zpět do jater nebo na LDL.

43
ENDOGENNÍ DRÁHA Receptor LDL B-100 LDL JÁTRA MIMOJATERNÍ TKÁŇ B-100
VLDL IDL HDL E C B-100 E LPL KAPILÁRA PLASMA LCAT

44
Apo E syntetizují jaterní buňky, astrocyty v mozku, buňky sleziny, nadledvin, vaječníku a ledvin.
Koncentrace apo E stoupá při nervovém poškození a tak vyrovnává koncentraci CH a tím se zúčastní regenerace nervů.

45
Metabolismus volných mastných kyselin.
Volné mastné kyseliny (FFA) pomocí chylomikronů a VLDL jdou do tukových buněk a buněk ostatních tkání. Mastné kyseliny (FA) se syntetizují v tukových zásobárnách, kde se současně skladují. Cirkulují v krvi vázané na albumin a jsou významným zdrojem energie pro všechny tkáně (včetně mozku). Oxidují se na CO2 a H2O.
 pomocí chylomikronů a VLDL jdou do tukových buněk a buněk ostatních tkání. Mastné kyseliny (FA) se syntetizují v tukových zásobárnách, kde se současně skladují. Cirkulují v krvi vázané na albumin a jsou významným zdrojem energie pro všechny tkáně (včetně mozku). Oxidují se na CO2 a H2O.")
46
Lipázy LPL na povrchu kapilárního endotelu hydrolyzuje TG v chylomikronech a VLDL na FFA a glycerol, které se v adipocytech slučují znovu na nové TG. Intracelulární lipáza senzitivní vůči účinkům hormonů (TG-lipáza, hormon senzitivní lipáza) katalyzuje odbourávání zásobních TG na glycerol a FA a FA putují do oběhu (stávají se FFA).
 katalyzuje odbourávání zásobních TG na glycerol a FA a FA putují do oběhu (stávají se FFA).")
47
TG-lipáza se působením cyklického AMP, který vzniká z ATP působením adenylátcyklázy, přes proteinokinázu mění z inaktivní formy na formu aktivní. Adenylátcyklázu v adipocytech aktivují katecholaminy před beta1-adrenergní receptory a snad i přes beta3-receptory.

48
katecholaminy beta1 receptor adenylátcykláza ATP cyklický AMP proteinkináza A aktivní lipáza neaktivní lipáza

49
Adenylátcyklázu aktivují ještě
glukagon GH glukokortikoidy T3 a T4 ACTH TSH LH serotonin vazopresin Pomalejší proces, vyžaduje tvorbu nového proteinu Fyziologická úloha neurčitá

50
Inzulín a prostaglandin E snižují aktivitu TG-lipázy
inhibicí tvorby cAMP snižují aktivitu TG-lipázy

51
katecholaminy beta1 receptor adenylátcykláza ATP cyklický AMP proteinkináza A aktivní lipáza neaktivní lipáza TG FFA + glycerol

52
Aktivitu TG-lipázy ještě ovlivňují
inzulín nasycenost hladovění stres - +

53
AKTIVITU TG-LIPÁZY ZVYŠUJÍ katecholaminy glukagon GH ACTH kortizol TSH
T3, T4 LH serotonin vazopresin hladovění stres SNIŽUJÍ inzulín nasycení

54
AKTIVITU TG-LIPÁZY ZVYŠUJÍ katecholaminy glukagon GH ACTH kortizol TSH
T3, T4 LH serotonin vazopresin hladovění stres SNIŽUJÍ inzulín nasycení Působí katabolicky Působí anabolicky

55
Zvyšují riziko aterosklerózy Zvyšují riziko obezity
AKTIVITU TG-LIPÁZY ZVYŠUJÍ katecholaminy glukagon GH ACTH kortizol TSH T3, T4 LH serotonin vazopresin hladovění stres SNIŽUJÍ inzulín nasycení Zvyšují riziko aterosklerózy Zvyšují riziko obezity

56
Metabolismus CH Základní prekrusor steroidních hormonů a žlučových kyselin a základní složka buněčných membrán. Vyskytuje se pouze u živočichů (vaječný žloutek a živočišný tuk). CH se vstřebává ze střeva a inkorporuje se do chylomikronů, které vznikají ve sliznici. Po odstranění TG v tukové tkáni chylomikronové zbytky transportují CH do jater.
. CH se vstřebává ze střeva a inkorporuje se do chylomikronů, které vznikají ve sliznici. Po odstranění TG v tukové tkáni chylomikronové zbytky transportují CH do jater.")
57
Metabolismus CH Játra a jiné orgány CH syntetizují.
Část CH je játry vylučována do žluče (volný CH nebo žlučové kyseliny). Část žlučového CH se resorbuje ve střevě zpět. Většina CH se v játrech inkorporuje do VLDL. CH zpětnou vazbou inhibuje svou vlastní syntézu (viz HMG-CoA)
. Část žlučového CH se resorbuje ve střevě zpět. Většina CH se v játrech inkorporuje do VLDL. CH zpětnou vazbou inhibuje svou vlastní syntézu (viz HMG-CoA)")
58
Hormony štítné žlázy, které zvyšují produkci receptorů pro LDL, snižují plazmatickou hladinu CH.
Estrogeny zvyšují HDL a snižují LDL (rovněž zvýšením počtu LDL receptorů v játrech = zrychlené odstranění LDL). CH stoupá při obstrukci toku žluči a při neléčeném DM.
. CH stoupá při obstrukci toku žluči a při neléčeném DM.")
59
Esenciální mastné kyseliny.
Některé nenasycené tuky představují nezbytnou součást potravy, zvláště u dětí kyselina linolenová kyselina linolová kyselina arachidonová Jejich nedostatek se může projevit zpomalením růstu (děti), kožními a ledvinovými poruchami a neplodností.
, kožními a ledvinovými poruchami a neplodností.")
60
Eikosanoidy eikosanoidy Esenciální mastné kyseliny jsou prekrusory
prostaglandinů prostacyklinů tromboxanů lipoxinů leukotrienů eikosanoidy Název odvozen od 20-uhlíkaté (eikosa-) mnohonásobně nenasycené FA (k. arachidonová) a od 20-uhlíkatých derivátů k. linolové a linolenové
 mnohonásobně nenasycené FA (k. arachidonová) a od 20-uhlíkatých derivátů k. linolové a linolenové.")
61
jsou tvořeny řadou nenasycených FA s 20-uhlíkatým řetězcem.
Prostaglandiny jsou tvořeny řadou nenasycených FA s 20-uhlíkatým řetězcem. Nejdříve byly izolovány ze spermií, ale syntetizují se ve všech tělesných orgánech. Prostaglandiny se dělí (např. na PGE a PGF) podle sestavení jejich cyklopentanového kruhu. Počet dvojných vazeb a jejich umístění v řetězci se vyznačuje číslicí v indexu.
 podle sestavení jejich cyklopentanového kruhu. Počet dvojných vazeb a jejich umístění v řetězci se vyznačuje číslicí v indexu.")
62
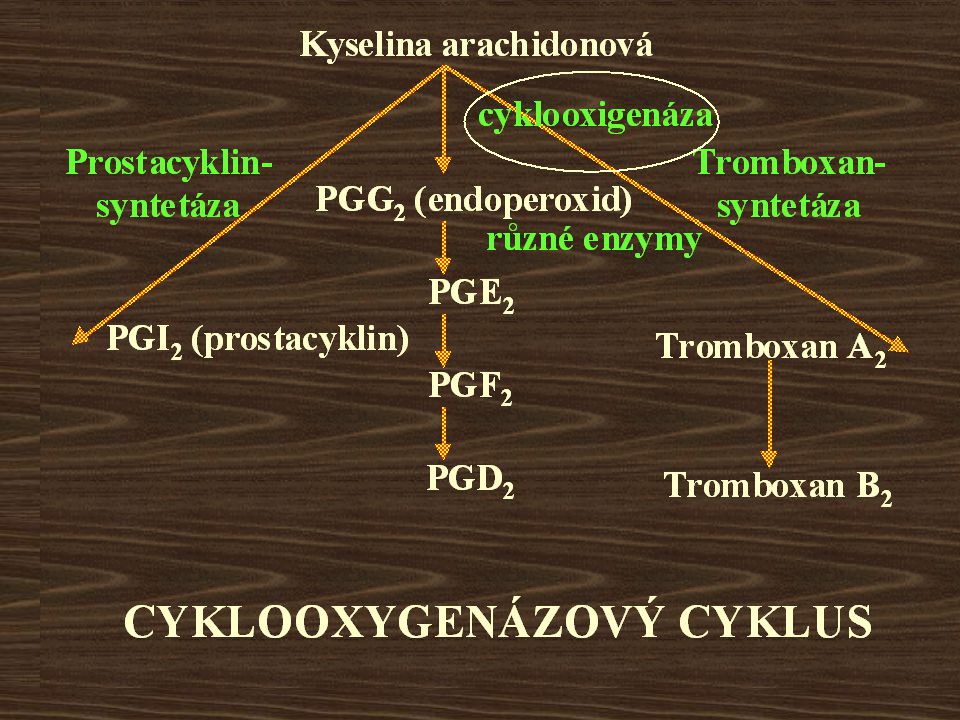
Kyselina arachidonová CYKLOOXYGENÁZOVÝ CYKLUS
cyklooxigenáza Prostacyklin- syntetáza PGG2 (endoperoxid) Tromboxan- syntetáza různé enzymy PGE2 PGI2 (prostacyklin) Tromboxan A2 PGF2 PGD2 Tromboxan B2 CYKLOOXYGENÁZOVÝ CYKLUS
 Tromboxan- syntetáza. různé enzymy. PGE2. PGI2 (prostacyklin) Tromboxan A2. PGF2. PGD2. Tromboxan B2. CYKLOOXYGENÁZOVÝ CYKLUS.")
63
Prostacykliny a tromboxany
Tromboxan A2 je tvořen trombocyty a agreguje destičky a způsobuje vázokonstrikci. Prostacyklin je tvořen endoteliálními buňkami a působí srážlivě jen v místě, kde je céva poraněná, ale lumen udržuje bez sraženiny - inhibuje agregaci destiček a podporuje vázodilataci.

64
Prostacykliny a tromboxany
Rovnováha mezi tromboxanem A2 a prostacyklinem udržuje místní agregaci destiček a následnou tvorbu sraženiny. Brání nadměrnému rozsahu sraženiny a udržuje kolem ní průtok.

65
Aspirin - protisrážlivý mechanismus?
Podávání nízkých dávek kyseliny acetylsalicylové (Aspirin) posunuje rovnováhu směrem k prostacyklinu. Aspirin vyvolává ireverzibilní inhibici cyklooxigenázy a tím sníží produkci tromboxanu A2 i prostacyklinu. Endoteliální buňky však dokáží vyprodukovat novou cyklooxygenázu v průběhu několika hodin, trombocyty ne.
 posunuje rovnováhu směrem k prostacyklinu. Aspirin vyvolává ireverzibilní inhibici cyklooxigenázy a tím sníží produkci tromboxanu A2 i prostacyklinu. Endoteliální buňky však dokáží vyprodukovat novou cyklooxygenázu v průběhu několika hodin, trombocyty ne.")
67
Prostacykliny a tromboxany
Hladina tromboxanu se může postupně zvyšovat až se vytvoří nové trombocyty (pomalý proces, poločas asi 4 dny). Proto podávání malých dávek Aspirinu po delší dobu snižuje tvorbu sraženiny a má preventivní význam u IM, nestabilní AP, přechodných ischemických příhod a centrální mozkové příhody.
. Proto podávání malých dávek Aspirinu po delší dobu snižuje tvorbu sraženiny a má preventivní význam u IM, nestabilní AP, přechodných ischemických příhod a centrální mozkové příhody.")
68
Leukotrieny Kyselina arachidonová se také přeměňuje na kyselinu 5-hydroxyeikosatetraenovou (5-HPETE), ze které vznikají leukotrieny.
, ze které vznikají leukotrieny.")
69
Kyselina arachidonová
5-lipoxygenáza 5-HPETE Leukotrien A4 + glutation Leukotrien B4 Leukotrien C4 Leukotrien F4 + k. glutamová - k. glutamová Leukotrien E4 Leukotrien D4 - glycin

71
Kyselina arachidonová
15-lipoxygenáza Lipoxin A Lipoxin B

72
Mají krátký poločas života a působí v místech, kde vznikají.
Leukotrieny, prostacykliny, tromboxany, lipoxiny a prostaglandiny jsou tzv. „MÍSTNÍ HORMONY“. Mají krátký poločas života a působí v místech, kde vznikají.

73
Leukotrieny jsou mediátory alergických reakcí při zánětu.
Způsobují bronchokonstrikci, vázokonstrikci, zvyšují vaskulární permeabilitu a přitahují leukocyty do místa zánětu.

Podobné prezentace


















.>")

