Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
MUTAGENEZE in vitro postupy kterými se mění primární struktura DNA, především za účelem fenotypové změny

2
Mutace gen mutace mutovaný gen transkripce translace normalní protein
normální fenotyp mutovaný gen abnormální protein částečně funkční nefunkční žádný změněný fenotyp

3
Typy mutací BODOVÉ DELECE INZERCE INVERZE TRANSLOKACE

4
Mutace na genové úrovni
MUTACE SE ZTRÁTOU SMYSLU 5’ ATG GGA GCT CTA TTA ACC TAA 3’ met gly ala leu leu thr stop 5’ ATG GGA GCT CTA TGA ACC TAA 3’ met gly ala leu stop MUTACE BEZ PROJEVU 5’ ATG GGA GCT CTA TTA ACC TAA 3’ met gly ala leu leu thr stop 5’ ATG GGA GCT CTA TTG ACC TAA 3’ ZMĚNA JEDNÉ AMINOKYSELINY 5’ ATG GGA GCT CTA TTA ACC TAA 3’ met gly ala leu leu thr stop 5’ ATG GGA GCT CTA TTT ACC TAA 3’ met gly ala leu phe thr stop MUTACE S POSUNEM ORF 5’ ATG GGA GCT CTA TTA ACC TAA 3’ met gly ala leu leu thr stop 5’ ATG GGG AGC TCT ATT AAC CTA A 3’ met gly ser ser ile asn leu

5
Mutace na úrovni organismu
Letální mutace – organismus umírá v důsledku změněného nebo chybějícího produktu genu. Může být letální na základě okolních podmínek Mutace způsobí pouze částečnou změnu funkce genu (např. snížení aktivity) Mutace genu jehož funkce není pro buňku esenciální nebo může být nahrazená genem podobným (genová rodina) Skrytá mutace (bez efektu)
 Mutace genu jehož funkce není pro buňku esenciální nebo může být nahrazená genem podobným (genová rodina) Skrytá mutace (bez efektu)")
6
PSEUDOGENY 1) Procesované (nebo retrotransposované) pseudogeny
vznikly reverzní transkripcí z RNA a zabudováním do genomu 2) Neprocesované (nebo duplikované) pseudogeny vznikly duplikací a následnýma spontánníma mutacema 3) Jedinečné pseudogeny jedinečné, důležité v evoluci. např. gulonolakton oxidasa pro biosyntézu vitamínu C
 Neprocesované (nebo duplikované) pseudogeny. vznikly duplikací a následnýma spontánníma mutacema. 3) Jedinečné pseudogeny. jedinečné, důležité v evoluci. např. gulonolakton oxidasa pro biosyntézu vitamínu C.")
7
REVERZE Proces kdy mutant získá zpět wild-typový fenotyp, dvě cesty:
1. zpětná mutace (přesná, málo častá) 2. supresorová mutace (mutace na jiném místě genu, která potlačuje (supresuje) původní mutaci. Mutant se nazývá REVERTANT. Je častá u mutací s posunem ORF.
 2. supresorová mutace (mutace na jiném místě genu, která potlačuje (supresuje) původní mutaci. Mutant se nazývá REVERTANT. Je častá u mutací s posunem ORF.")
8
Amesův Test test měří mutagenicitu různých látek jako zvýšenou frekvenci spontánní reverze mutanta his- to his+ POUŽITÍ: testování nových látek na mutagenitu, potravinářská aditiva, pesticidy, kosmetika atd. histidin-vyžadující mutant (his-) Salmonella typhimurium Test chemikálie + bakterie minimální medium bez histidinu 48 hrs Backgroundová spontánní REVERZE Chemicky indukovaná REVERZE
 Salmonella typhimurium. Test chemikálie + bakterie. minimální medium bez histidinu. 48 hrs. Backgroundová spontánní REVERZE. Chemicky indukovaná. REVERZE.")
9
gen rezistence k antibiotiku
TRANSPOZOMY mobilní elementy DNA které se pohybují v rámci genomu, s četnosti až 10-2 přenosu na generaci. tento přenos je TRANSPOZICE pokud se tento transpozom dostane do genu chová se jako inzertní mutace, s tím že tuto mutaci nelze vrátit zpět reverzí. transpozice je řídký jev, můžeme však do transpozomu vložit gen pro rezistenci k antibiotiku, využívá se při mutaci zprostředkované transpozomem Transpozomy jsou důvodem proč spousta kmenů bakterií v nemocnicích je rezistantní vůči širokému spektru antibiotik. Většina resistence je buď na plasmidech nebo transpozomech, které se mohou mezi jednotlivými kmeny vyměňovat, což je posíleno selekčním tlakem prostředí (spousta antibiotik v nemocnici). Staphylococcus aureus gen rezistence k antibiotiku IS Element
. Staphylococcus aureus. gen rezistence k antibiotiku. IS Element.")
10
SPONTÁNNÍ MUTACE FLUKTUAČNÍ TEST mutace je náhodný proces
obecně dochází ke spontánní mutaci s pravděpodobnosti 10-6 až 10-9 na gen za jednu generaci každý gen mutuje s jinou pravděpodobností (pozice v genomu) mutace ovlivňující fenotyp je ještě daleko řidší pravděpodobnost reverze je daleko nižší než přímé mutace EVOLUCE letitý spor zda je variabilita mezi organismy způsobena ADAPTACI NA PROSTŘEDÍ nebo SPONTÁNNÍMI MUTACEMI a až zpětnou adaptaci FLUKTUAČNÍ TEST spontánní mutace vznikají chybou v replikaci DNA, 3-5 exonukleasová (PROOF-READING) aktivita DNA polymeras (ne u PCR!) působení fága 1943
 mutace ovlivňující fenotyp je ještě daleko řidší. pravděpodobnost reverze je daleko nižší než přímé mutace. EVOLUCE letitý spor zda je variabilita mezi organismy způsobena ADAPTACI NA PROSTŘEDÍ nebo SPONTÁNNÍMI MUTACEMI a až zpětnou adaptaci. FLUKTUAČNÍ TEST. spontánní mutace vznikají chybou v replikaci DNA, 3-5 exonukleasová (PROOF-READING) aktivita DNA polymeras (ne u PCR!) působení fága ")
11
GENETICKÉ MAPOVÁNÍ často používaná metoda pro zjišťování funkce neznámých genů
náhodná mutageneze semen Arabidopsis způsobující fenotypovou změnu mutagen ethylmethan sulfonát (EMS), záření rentgenové nebo gamma. semen se pěstuje a pozoruje fenotyp - M1 generace, mutace je heterozygotní. M1 generace se samozkříží a semena se seberou. M2 semena se vysejí a mutanti s požadovaným fenotypem identifikují (ti co produkují pouze potomstvo s fenotypem jsou homozygotní na mutaci).
, záření rentgenové nebo gamma semen se pěstuje a pozoruje fenotyp - M1 generace, mutace je heterozygotní. M1 generace se samozkříží a semena se seberou. M2 semena se vysejí a mutanti s požadovaným fenotypem identifikují (ti co produkují pouze potomstvo s fenotypem jsou homozygotní na mutaci).")
12
GENETICKÉ MAPOVÁNÍ ? mapa Arabidopsisových molekulárních markerů I
121.4 cM II 99.1 cM III 101.7 cM IV 92.0 cM V 104.0 cM nga 63 nga 126 nga 225 nga 8 nga 151 DET1 G4711 nga 139 mutace a její pozice ? CH42 GPA1 GAPB NIT1 nga 168 nga 280 PRHA DFR I-V 518.2 cM nga 111 mapa Arabidopsisových molekulárních markerů

13
GENETICKÉ MAPOVÁNÍ mutaci mapujeme pomocí molekulárních markerů u Arabidopsis thaliana ekotyp Columbia další rozšířený ekotyp Arabidopsis thaliana je Landsberg erecta pomocí DNA markerů můžeme rozlišit mezi těmito dvěmi ekotypy existuje na sekvenčních polymorfismů mezi ekotypy Landsberg erecta a Columbia a jejich pozice na chromozomech je přesně známa. tyto rozdíly se detekují především pomocí PCR variabilita v rámci inzertů a delecí způsobující rozdílnou délku amplifikovaných fragmentů, nebo vznikem nového restrikčního místa. příklad polymorfismu mezi Landsberg erecta a Columbia – vznik nového restrikčního místa DraI v PCR produktu markeru

14
crossing over a rekombinace během meiozy při tvorbě gamet u heterozygotů (výměna části chromozomů)
x x Strain 1 Columbia mut (-/-) Strain 2 Landsberg mut (+/+) F1 rostliny mut (-/+) BACK CROSSING markery ve vazbě mut { mut (-/-) mut (+/-) F2 semena
 Strain 2. Landsberg. mut (+/+) F1 rostliny. mut (-/+) BACK. CROSSING. markery. ve vazbě. mut { mut (-/-) mut (+/-) F2 semena.")
15
četnost genotypu po crossing overu
GENETICKÉ MAPOVÁNÍ homozygotní selektovaný mutant ekotypu Columbia homozygotní nemutovaný ekotypu Landsberg erecta heterozygot tzv. BACKCROSSING (zpětné křížení s mutantem) semena v F2 generaci poskytující mutantní fenotyp musí být homozygoti vzniklí crossing overem četný genotyp marker A není ve vazbě s mutaci četnost genotypu po crossing overu vzácný genotyp marker B je ve slabé vazbě s mutaci velmi vzácný genotyp marker C je ve vazbě s mutaci
 semena v F2 generaci poskytující mutantní fenotyp musí být homozygoti vzniklí crossing overem. četný genotyp marker A není ve vazbě s mutaci. četnost genotypu po crossing overu. vzácný genotyp marker B je ve slabé vazbě s mutaci. velmi vzácný genotyp marker C je ve vazbě s mutaci.")
16
analýza zpětně zkřížených mutantů
frekvence rekombinace (RF) mezi mutovaným lokusem a různými markery RF [%] = # chromozomů Landsberg / # celkový počet chromozomů x 100 čím nižší procento, tím silnější vazba marker I ve vazbě 5 mutantů 1/10x100 = 10% marker II bez vazby na mutaci 6 mutantů 5/12x100 = 58% většinou se analyzuje až 1000 zpětně zkřížených mutantů, ten s nejméně četným genotypem na testovaný marker pro druhý ekotyp má tento marker nejblíže mutaci určení dvou nejbližších markeru a následná sekvenace chromosomu mezi a nalezení hledané mutace resp. genu nesoucího tuto mutaci
 mezi mutovaným lokusem a různými markery. RF [%] = # chromozomů Landsberg / # celkový počet chromozomů x 100. čím nižší procento, tím silnější vazba. marker I ve vazbě. 5 mutantů. 1/10x100 = 10% marker II bez vazby na mutaci. 6 mutantů. 5/12x100 = 58% většinou se analyzuje až 1000 zpětně zkřížených mutantů, ten s nejméně četným genotypem na testovaný marker pro druhý ekotyp má tento marker nejblíže mutaci. určení dvou nejbližších markeru a následná sekvenace chromosomu mezi a nalezení hledané mutace resp. genu nesoucího tuto mutaci.")
17
GENETICKÉ MAPOVÁNÍ člověk
shromážděno největší dostupné množství příslušníků rodin ve kterých se nachází znak (nemoc), kterou chceme mapovat – určit gen, který nemoc způsobuje. Odebrány vzorky krve, izolována DNA a provedeno PCR na zhruba 200 genových markerů - polymorfismů (pokrývající celý genom cM od sebe) nalezen nejbližší marker tzn. jedna forma (alela) se nejčastěji vyskytuje u nemocného a u jeho zdravých sourozenců se nejčastěji vyskytuje druhá forma (marker ve vazbě k mapovanému genu – nejmenší pravděpodobnost crossing overu) 2 kolo – provedeno PCR na další nejbližší markery v nejbližším okolí markeru nalezeného v prvním kole testování. nakonec nalezeny dva nejbližší markery a DNA mezi nimi sekvencována u nemocného jedince a porovnána se zdravým.
, kterou chceme mapovat – určit gen, který nemoc způsobuje. Odebrány vzorky krve, izolována DNA a provedeno PCR na zhruba 200 genových markerů - polymorfismů (pokrývající celý genom cM od sebe) nalezen nejbližší marker tzn. jedna forma (alela) se nejčastěji vyskytuje u nemocného a u jeho zdravých sourozenců se nejčastěji vyskytuje druhá forma (marker ve vazbě k mapovanému genu – nejmenší pravděpodobnost crossing overu) 2 kolo – provedeno PCR na další nejbližší markery v nejbližším okolí markeru nalezeného v prvním kole testování. nakonec nalezeny dva nejbližší markery a DNA mezi nimi sekvencována u nemocného jedince a porovnána se zdravým.")
18
GENETICKÉ MAPOVÁNÍ člověk - příklad
Geny ve vazbě 2/13 transkripční faktor LMX1B

19
GENETICKÉ MAPOVÁNÍ myš
Inbrední linie myší – homozygotní téměř ve všech lokusech, vzniká křížením sourozenců minimálně po 20 generací

20
Kongenní myš – vzniklá zpětným křížením potomstva dvou inbredních linií, z niž jedna nesla znak, který chceme alokovat, minimálně přes deset generací a selekcí na sledovaný znak Velikost diferenciálního segmentu v průběhu kongenizace Alokace lokusů pomocí rozdílnosti mikrosatelitních markerů u jednotlivých inbredních myších linií Průměrná vzdálenost u myší – 6.7 cM

21
MUTACE CÍLENÁ studuje efekt změny v genetickém materiálu
- modifikace promotorové sekvence za účelem studia účinnosti transkripce - studium významu jednotlivých AK v proteinu zlepšování kvality např. expresního systému KOZAKOVÁ SEKVENCE je důležitá pro efektivitu translace (umožňuje pevnou vazbu malé ribozomální podjednotky na mRNA) u eukaryot. nebo proteinu, který exprimujeme změna jedné nebo i více aminokyselin může zlepšit kvalitu či aktivitu léčiva (enzymu) start codon Kozak seq U A A A C A A U G G C U 60% % % AtCKX G U A G A A A U G G G A AtCKX A A A C A A A U G G C U HvCKX A G A G C C A U G A G G
 u eukaryot. nebo proteinu, který exprimujeme. změna jedné nebo i více aminokyselin může zlepšit kvalitu či aktivitu léčiva (enzymu) start codon Kozak seq. U A A A C A A U G G C U. 60% 100% 70% AtCKX1 G U A G A A A U G G G A. AtCKX2 A A A C A A A U G G C U. HvCKX2 A G A G C C A U G A G G.")
22
mutace místně cílená (site-directed mutagenesis)
Stratagene gen, či sekvenci kterou budeme chtít mutovat, je třeba naklonovat do vhodného vektoru navržení dvou komplementárních primerů, v místě kde chceme mutovat, nesoucí tuto mutaci M G A L L W L původní sekvence ’ ATG GGA GCT CTA TTA ACC TTA 3’ forward primer ’ TAC CCT CGA GAT AAT TCG AAT 5’ reverse primer ’ ATG GGA GCT CTA TTA AGC TTA 3’ M G A L L S L PCR s těmito primery na templátový plasmid a s Pfu polymerasou vytváří se nové cirkulární DNA nesoucí mutaci, jsou k sobě komplementární a drží u sebe, mají přerušení v místě konce primerů (tzv. nick) ošetření restrikční endonukleasou DpnI (štěpí pouze methylovanou DNA, templátový plasmid) transformace do bakterie a namnožení mutovaného plasmidu
 ošetření restrikční endonukleasou DpnI (štěpí pouze methylovanou DNA, templátový plasmid) transformace do bakterie a namnožení mutovaného plasmidu.")
23
TRANSFORMACE baktérii a kvasinek přímý genetický přenos informace (DNA) z okolí do organismu
buňka schopná přijmout DNA (plasmid) se nazývá KOMPETENTNÍ přirozeně kompetentní jsou některé kmeny Bacillus subtilis, Hemorheae influenze atd. všechny ostatní se mohou transformovat po uvedení do kompetentního stavu dvěma způsoby: A) ELEKTROPORACE B) CHEMICKÁ METODA buňky se pořádně promyjí diH2O smíchají s plasmidovou DNA rozpuštěnou také v diH2O a vloží do elektroporátoru buňky se ošetří roztokem rubidné a vápenaté soli, které způsobují větší permeabilitu membrány DNA se smísí s těmito buňkami a provede se tzv. HEAT SHOCK (45 sec. 42°C) EFEKTIVITA NÁROČNOST
 se nazývá KOMPETENTNÍ. přirozeně kompetentní jsou některé kmeny Bacillus subtilis, Hemorheae influenze atd. všechny ostatní se mohou transformovat po uvedení do kompetentního stavu dvěma způsoby: A) ELEKTROPORACE B) CHEMICKÁ METODA. buňky se pořádně promyjí diH2O. smíchají s plasmidovou DNA rozpuštěnou. také v diH2O a vloží do elektroporátoru. buňky se ošetří roztokem rubidné a vápenaté soli, které způsobují větší permeabilitu membrány. DNA se smísí s těmito buňkami a provede se tzv. HEAT SHOCK (45 sec. 42°C) EFEKTIVITA NÁROČNOST ")
24
využití mutageneze při studiu regulačních sekvencí genů
vytvoření konstruktu studovaného promotoru a reportérového genu promotor reportérový gen vytvoření mutantů pro skenování promotorové aktivity DNA transformace každého konstruktu do buněk lýze buněk a měření aktivity reportérového genu

25
využití mutageneze při studiu regulačních sekvencí genů
konstrukt aktivita reportérového genu reportérové geny: ß-galaktosidasa (kolorimetricky) CAT (chloramfenicolacetyl transferasa) (kolorimetricky) luciferasa (luminiscenčně)
 CAT (chloramfenicolacetyl transferasa) (kolorimetricky) luciferasa (luminiscenčně)")
26
C) mutace kazetová
 mutace kazetová")
27
C) mutace kazetová
 mutace kazetová")
28
C) mutace kazetová dvojitá
 mutace kazetová dvojitá")
29
D) mutace supresorovou tRNA
mutace se ztrátou smyslu (mutace na stop kodon) muže být potlačená tzv. supresorovou tRNA existují mutantní tRNA geny (mutace v antikodonu), které rozpoznávají tyto kodony jako funkční a začleňují do vznikajícího polypetidu místo ukončení další AK tzv. supresorové tRNA. ty mohou být pro libovolnou AK, používají se např. pro vnášení speciálních AK do proteinu (nutno vnést i nový gen pro aminoacyl tRNA syntasu)
 muže být potlačená tzv. supresorovou tRNA. existují mutantní tRNA geny (mutace v antikodonu), které rozpoznávají tyto kodony jako funkční a začleňují do vznikajícího polypetidu místo ukončení další AK tzv. supresorové tRNA. ty mohou být pro libovolnou AK, používají se např. pro vnášení speciálních AK do proteinu (nutno vnést i nový gen pro aminoacyl tRNA syntasu)")
30
- příprava knockoutovaných linií organismů
CÍLENÁ MANIPULACE GENOMU - příprava knockoutovaných linií organismů HR – homologní rekombinace NHEJ – nehomologní lepení konců

31
ZINC FINGER NUCLEASES chimerické proteiny vytvořené spojením Fok I nukleasy a specifických sekvencí zvaných „zinkové prsty“

32
ZINC FINGER NUCLEASES

33
Testování specifity ZFN

34
Knock-out genu pomocí ZFN
Shukla et al (2009): Precise genome modification in the crop species using ZFN. Nature 459
: Precise genome modification in the crop species using ZFN. Nature 459.")
35
SEKVENCOVÁNÍ GENOMŮ

36
sekvencování jednotlivých klonů (srovnávací a integrační)
SEKVENCOVÁNÍ GENOMU příprava knihovny sekvencování jednotlivých klonů sestavení genomu vyplnění mezer předpovězení genů anotace genů analýza genomu (srovnávací a integrační)
")
37
SEKVENCOVÁNÍ GENOMU genetická mapa 5.264 mikrosatelitů
mapování genomu (chromozomu): genomové mapování bylo umožněno objevem specifických abundantních genetických markeru (mikrosatelitů) do 1994, bylo na lidské genomové mapě lokalizováno: 5.264 mikrosatelitů na chromozomových lokusech (průměrná vzdálenost mezi markery je 599 kb) bylo také odsekvenováno tisíce STS sequence tagged site (STS) genetická mapa mikrosatelitů
: genomové mapování bylo umožněno objevem specifických abundantních genetických markeru (mikrosatelitů) do 1994, bylo na lidské genomové mapě lokalizováno: mikrosatelitů na chromozomových lokusech. (průměrná vzdálenost mezi markery je 599 kb) bylo také odsekvenováno tisíce STS sequence tagged site (STS) genetická mapa mikrosatelitů.")
38
SEKVENCOVÁNÍ GENOMU Dva typy přístupů pro sekvencování genomů: 1. Metoda PROCHÁZENÍ CHROMOZOMU (Chromosome walking) vychází z mapy genomu, začíná se od markeru a sekvencuje se klon za klonem levné, ale zdlouhavé pro sestrojení knihovny se používají BAC vektory (kapacita 300kb)
")
39
SEKVENCOVÁNÍ GENOMU 2. Metoda SHOTGUN SEKVENCOVÁNÍ
vytvoření BAC klonů pro každý chromozom (jeden lidský chromozom se vejde asi do tři stovek BAC klonů). rozrušení inzertů (Sau3AI, nebo kombinace RE), separace na gelu podle délek a překlonování ±2kbp fragmentů do normálních plasmidových vektorů koncové sekvencování pomocí univerzálních vektorových primerů sekvenace všech inzertů (jedna sekvenační reakce umožní přečtení max. 700 bází) počítačové poskládáni překrývajících se úseků (kontigů) do větších celků a vytvoření superkontigů (superrychlé počítače) drahá, ale rychlá metoda (většina velkých genomů byla odsekvencována takto)
. rozrušení inzertů (Sau3AI, nebo kombinace RE), separace na gelu podle délek a překlonování ±2kbp fragmentů do normálních plasmidových vektorů. koncové sekvencování pomocí univerzálních vektorových primerů. sekvenace všech inzertů (jedna sekvenační reakce umožní přečtení max. 700 bází) počítačové poskládáni překrývajících se úseků (kontigů) do větších celků a vytvoření superkontigů (superrychlé počítače) drahá, ale rychlá metoda (většina velkých genomů byla odsekvencována takto)")
40
SEKVENCOVÁNÍ GENOMU genom náhodné štípání plazmidy (2 – 10 Kbp)
kosmidy (40 Kbp) BAC (300Kbp) ~500 bp ~500 bp ~500 bp ~500 bp koncové sekvenování ( bp)
 BAC (300Kbp) ~500 bp. ~500 bp. ~500 bp. ~500 bp. koncové sekvenování ( bp)")
41
SEKVENCOVÁNÍ GENOMU

42
U VELKÝCH EUKARYOTICKÝCH GENOMŮ JE S 2 kb KLONOVANÝMI ÚSEKY PROBLÉM
SEKVENCOVÁNÍ GENOMU U VELKÝCH EUKARYOTICKÝCH GENOMŮ JE S 2 kb KLONOVANÝMI ÚSEKY PROBLÉM velikost repetitivních sekvencí může být až 5 kb mnoho 2 kb klonů proto obsahovalo dlouhé repetitivní sekvence výsledkem bylo zastavení sestavování genomu a přerušení sekvence dané úseky se musely naklonovat znova ve větších fragmentech tak aby bylo možno sestavit kontigy (10kb)
")
43
REPETITIVNÍ SEKVENCE EUKARYOTICKÉHO GENOMU

44
REPETITIVNÍ SEKVENCE EUKARYOTICKÉHO GENOMU
Nekódující DNA (90-95%) nachází se mezi geny (extragenová) nebo v genech (intragenová) Single Copy DNA jedinečná Repetitivní DNA 30 % celkové jaderné DNA repetitivní geny (histony, rRNA, tRNA) >105 kopií kopií Mírně repetitivní Vysoce repetitivní (tandemová) SINEs Short interspersed elements. např. lidské ALU segmenty LINEs Long interspersed elements. např. lidské Kpn segmenty Minisatelity 15bp repetice náhodně roztroušené Satelitní DNA dlouhé repetice kolem centromer a telomer Mikrosatelity 2-6 bp repetice náhodně roztroušené a variabilní v počtu opakování v rámci populace VNTRs Variable number Tandem Repeats GC-rich 300 bp bp AT-rich 1.5-6 kb bp
 nachází se mezi geny (extragenová) nebo v genech (intragenová) Single Copy DNA. jedinečná. Repetitivní DNA. 30 % celkové jaderné DNA. repetitivní geny (histony, rRNA, tRNA) >105. kopií kopií. Mírně repetitivní. Vysoce repetitivní (tandemová) SINEs. Short interspersed elements. např. lidské ALU segmenty. LINEs. Long interspersed elements. např. lidské Kpn segmenty. Minisatelity. 15bp repetice. náhodně roztroušené. Satelitní DNA. dlouhé repetice kolem centromer a telomer. Mikrosatelity. 2-6 bp repetice. náhodně roztroušené a variabilní v počtu opakování v rámci populace. VNTRs. Variable number. Tandem Repeats. GC-rich. 300 bp bp. AT-rich kb bp.")
45
PARAZITICKÁ DNA

46
PROJEKT SEKVENACE LIDSKÉHO GENOMU
Dva hlavní projekty: Human Genome Project (HGP) mezinárodní konsorcium placeno z veřejných peněz Francis Collins, National Human Genome Res. Inst. (NHGRI) začalo fungovat 1990 v USA sekvenování pomocí genomových map Celera Genomics Corporation (CRA) soukromá firma J. Craig Venter, založeno 1998 shotgun sekvencování Obě skupiny vycházely ze vzorku DNA izolované z krve a spermatu anonymních dárců mužského i ženského pohlaví různých etnik
 mezinárodní konsorcium placeno z veřejných peněz. Francis Collins, National Human Genome Res. Inst. (NHGRI) začalo fungovat 1990 v USA. sekvenování pomocí genomových map. Celera Genomics Corporation (CRA) soukromá firma. J. Craig Venter, založeno shotgun sekvencování. Obě skupiny vycházely ze vzorku DNA izolované z krve a spermatu anonymních dárců mužského i ženského pohlaví různých etnik.")
47
PROJEKT SEKVENACE LIDSKÉHO GENOMU
16 únor 2001: první kompaktní výsledky publikováné současně v Nature & Science. HGP: ~22.1 miliard odsekvenovaných nukleotidů 7x překryv CELERA: ~14.5 miliard odsekvenovaných nukleotidů 4.6x překryv 26.4 millionů 550 bp sekvenačních reakcí POKRYTO >99% genomu 20,000 CPU hodin (833 CPU dnů) na superpočítači
 na superpočítači.")
48
PROJEKT SEKVENACE LIDSKÉHO GENOMU
Identifikace genů v DNA sekvenci: ANOTACE GENU – identifikace a popis pravděpodobného genu a předpovězeni jeho funkce počítačový algoritmus pro všechny ORF open reading frame ORF – potenciální kódující sekvence pro protein začínající start kodónem a končící stop kodónem ne všechny ORFs kódují proteiny (6-7% nekódují u kvasinek) minimální délka ORF (100bp) velmi složitý algoritmus pro ORF s introny (eukaryota) hledání homologie v databázích – předpovědění funkce a evolučních vztahu
 minimální délka ORF (100bp) velmi složitý algoritmus pro ORF s introny (eukaryota) hledání homologie v databázích – předpovědění funkce a evolučních vztahu.")
49
Solitární gen: • v celém genomu v jediné kopii (asi polovina genů) Genová rodina: • skupina genů evolučně pocházející z jediného genu, v evoluci postupná diverzifikace sekvence a funkce Pseudogen: • gen který zmutoval natolik že nemůže být přepisován (v celém genomu > ) Zpracovaný (“processed”) pseudogen: • pseudogen vzniklý zpětným přepisem mRNA a integrací do genomu
 Zpracovaný ( processed ) pseudogen: • pseudogen vzniklý zpětným přepisem mRNA a integrací do genomu.")
51
PROJEKT SEKVENACE LIDSKÉHO GENOMU
CELKOVÁ VELIKOST ± bp (haploidní stav) méně genů než se očekávalo: 30-40,000 (2001) předpovídalo se 150,000 (před sekvenací) poslední odhady (2007) méně než Arabidopsis (25.478), červi (19.500) mezidruhové odlišnosti druhu Homo sapiens v rámci celého genomu kolem 0.1% (většina je v nekódujících sekvencích) 99% homologie s ostatními primáty (v genech), 96% (celkem) 1.5% z celkové sekvence genomu kóduje proteiny, 28% je však transkribováno do mRNA geny se nacházejí obecně v GC-bohatých regionech parazitická DNA tvoří 45% genomu. Tyto transpozomální elementy však už většinou nejsou aktivní (stále jsou ale aktivní u myši) 223 genů je z bakterií získaných paraziticky (nebyly nalezeny v kvasinkách ani octomilce)
 méně genů než se očekávalo: 30-40,000 (2001) předpovídalo se 150,000 (před sekvenací) poslední odhady (2007) méně než Arabidopsis (25.478), červi (19.500) mezidruhové odlišnosti druhu Homo sapiens v rámci celého genomu kolem 0.1% (většina je v nekódujících sekvencích) 99% homologie s ostatními primáty (v genech), 96% (celkem) 1.5% z celkové sekvence genomu kóduje proteiny, 28% je však transkribováno do mRNA. geny se nacházejí obecně v GC-bohatých regionech. parazitická DNA tvoří 45% genomu. Tyto transpozomální elementy však už většinou nejsou aktivní (stále jsou ale aktivní u myši) 223 genů je z bakterií získaných paraziticky (nebyly nalezeny v kvasinkách ani octomilce)")
52
pomáhá rozpoznat důležité regulační regiony.
KOMPARATIVNÍ GENOMIKA: sleduje konzervativní sekvence mezi různými organismy pomáhá rozpoznat důležité regulační regiony. myš a člověk mají téměř totožné geny rozdíly pravděpodobně v jejích regulaci a síle exprese homologní úseky jsou na různých chromozomech uplatňuje se ve velké míře alternativní sestřih (jedná mRNA produkuje různé proteiny) ostrovy genetické stability
 ostrovy genetické stability.")
53
ZAJÍMAVOSTI Chromozom 1 nese nejvíc genů (2968), a nejméně mužský Y chromozom 231 (78 krátké raménko). bylo objeveno kolem 3 miliónu míst kde dochází k mutacím (SNP – single nucleotide polymorfism) – příslib pro léčbu geneticky podmíněných chorob největší rozdíly jsou v genech spojených se sluchem a čichem v oblasti genomu kde jsou obsaženy geny spojeny s čichem, bylo nalezeno nejvíce mutací 50 genů šimpanzům chybí
 – příslib pro léčbu geneticky podmíněných chorob. největší rozdíly jsou v genech spojených se sluchem a čichem. v oblasti genomu kde jsou obsaženy geny spojeny s čichem, bylo nalezeno nejvíce mutací. 50 genů šimpanzům chybí.")
54
OSTATNÍ ODSEKVENOVANÉ GENOMY

55
OSTATNÍ ODSEKVENOVANÉ GENOMY první odsekvenovaný genom žijícího organismu Haemophilus influenzae 1995 baktérie způsobující zánět mozkových blan u dětí rok 2001 1981 2002 2003 2009 ??? 1998 2000 1996 organismus Velikost (Mbp) počet genů člověk (Homo sapiens) lidská mitochondriální DNA 3.200 0.016 32 laboratorní myš (M. musculus) 2.600 ± rýže (Oryza sativa) huseníček (A. thaliana) kukuřice (Zea mays) pšenice (Triticum aestivum) 430 125 2.500 15.000 ± 25.498 ± hlíst (C. elegans) 97 ± octomilka (D. melanogaster) 137 13.472 kvasinka (S. cerevisiae) 12.1 5.770 bakterie (E. coli) 4.6 4.377 virus (HIV) 0.009 9 kvasinka člověk
 počet genů. člověk (Homo sapiens) lidská mitochondriální DNA laboratorní myš (M. musculus) ± rýže (Oryza sativa) huseníček (A. thaliana) kukuřice (Zea mays) pšenice (Triticum aestivum) ± ± hlíst (C. elegans) 97. ± octomilka (D. melanogaster) kvasinka (S. cerevisiae) bakterie (E. coli) virus (HIV) kvasinka. člověk.")
56
organismus s největším známým genomem: Psilotum nudum 250.000 Mbp
další: česnek, ropucha, borovice, bahník PRUTOVKA HOLÁ (Psilotum nudum) rostlina, která "zapomněla" vymřít Existují pouze dva druhy prutovek. Mají pro botaniku zvláštní význam z hlediska evoluce rostlin. Velmi se podobají zcela vymřelým psilofytům, výtrusným rostlinám známým jen ze silurských až devonských zkamenělin, starých přes 300 milionů let. Psilofyty stály na samém počátku vývoje všech vyšších neboli cévnatých rostlin, jež potom úplně okupovaly zprvu pusté pevniny naší planety. Roste na Havaji.
 rostlina, která zapomněla vymřít Existují pouze dva druhy prutovek. Mají pro botaniku zvláštní význam z hlediska evoluce rostlin. Velmi se podobají zcela vymřelým psilofytům, výtrusným rostlinám známým jen ze silurských až devonských zkamenělin, starých přes 300 milionů let. Psilofyty stály na samém počátku vývoje všech vyšších neboli cévnatých rostlin, jež potom úplně okupovaly zprvu pusté pevniny naší planety. Roste na Havaji.")
57
VÝZNAM SEKVENACE GENOMU
počet genů, přesná poloha a funkce regulace genů struktura chromozomů a jeho organizace funkce nekodující sekvence, typy, množství a distribuce koordinace genové exprese a postranslační úpravy PROTEOMIKA interakce protein – protein předpovězená vs. experimentálně ověřená funkce evolučně konzervativní vztahy mezi organismy MEDICÍNA korelace (SingleNucleotidePolymorfism) s nemocemi předpovídání náchylnosti k nemocem na základě sekvenční variability (identifikace regionů DNA spojených s multigenovými nemocemi) navrhování léčiv na základě molekulární informace navrhování léčiv na míru na základě individuální genetického profilu (farmakogenomika)
 s nemocemi. předpovídání náchylnosti k nemocem na základě sekvenční variability (identifikace regionů DNA spojených s multigenovými nemocemi) navrhování léčiv na základě molekulární informace. navrhování léčiv na míru na základě individuální genetického profilu (farmakogenomika)")
58
mezinárodní program mapující lidskou druhovou variabilitu
HapMap mezinárodní program mapující lidskou druhovou variabilitu • konsorcium vědců ze 6 států od roku 2002 – se sestavují mapy vzorců SNP vyskytující v rámci populací jednotlivých ras v Africe, Asii a Spojených státech • cílem je dramaticky snížit celkový počet nalezených SNP ( ) selektování těch, které souvisejí s geneticky podmíněnými nemocemi
 selektování těch, které souvisejí s geneticky podmíněnými nemocemi.")
59
Pyrosekvencování DNA polymerasa dNTP mix ATP sulfurylasa
Adenosine 5 fosfosulfát Luciferasa Luciferin Apyrasa

60
Izolace genomové DNA a restrikce na vhodné fragmenty (300-800 bp)
Zatupení konců a navázání adaptorové sekvence s biotinovou značkou (modrá) a druhé bez (červená) Navázání na kuličky se streptavidinem Nanesení na mikroreaktorovou destičku Provedení emulsního PCR (červený a modrý primer) Paralelní pyrosekvencování ve všech jamkách
 a druhé bez (červená) Navázání na kuličky se streptavidinem. Nanesení na mikroreaktorovou destičku. Provedení emulsního PCR (červený a modrý primer) Paralelní pyrosekvencování ve všech jamkách.")
61
Roche 454/GS FLX Sequencing Technology

62
Illumina/Solexa metoda cyklické reverzibilní terminace 1. 3. 2. 6. 4.
5.

63
„Bridge“ PCR

64
cyklická reverzibilní terminace
Chemické štěpení ve vodném prostředí za katalýzy paladiem DEALLYLACE

65
Illumina/Solexa - vyhodnocení

66
Cena přečtení lidského genomu
SROVNÁNÍ platforma Příprava templátu chemismus Délka jednoho čtení (bp) Doba běhu Přečtené Gb na jeden běh Cena přístroje (USD) Cena přečtení lidského genomu ROCHE 454 Fragmentace + emulsní PCR pyrosekvencování 330 8 hod. 0.45 ILLUMINA SOLEXA Fragmentace + „bridge“ PCR Cyklická reverzibilní terminace 35 9 dní ROCHE dlouhé běhy, lze dělat i de novo nekomplikované genomy (mikroorganismy) + rychlé -- chyby v homopolymerních repeticích (AAAAAAAAA, GGGGGGGGGGG) ILLUMINA/ + nejpoužívanější SOLEXA + obrovský rozsah, resekvenace celého genomu v jednom běhu -- nízká multiplexita
 Doba běhu. Přečtené. Gb. na jeden běh. Cena přístroje. (USD) Cena přečtení lidského genomu. ROCHE 454. Fragmentace + emulsní PCR. pyrosekvencování hod ILLUMINA. SOLEXA. Fragmentace + „bridge PCR. Cyklická reverzibilní terminace dní ROCHE dlouhé běhy, lze dělat i de novo nekomplikované genomy (mikroorganismy) + rychlé. -- chyby v homopolymerních repeticích (AAAAAAAAA, GGGGGGGGGGG) ILLUMINA/ + nejpoužívanější. SOLEXA + obrovský rozsah, resekvenace celého genomu v jednom běhu. -- nízká multiplexita.")
67
Metagenomika Metagenom Genom X
Pyrosekvencování a technologie Roche 454 umožnili rozvoj metagenomiky: Rychlé a levné sekvencování genomů jednotlivců Nelze sekvencovat neznáme genomy, kvůli obtížnosti přiřazení (repetice) pouze ty u kterých je znám homologický genom (stejného biologického druhu) Metagenomika studium biologické rozmanitosti a genetického materiálu celé komunity (mikro)organismů přímo v jejich přirozeném prostředí bez nutnosti jejich kultivace v laboratoři (např. z půdy, střevní mikroflora) Metagenom Genom X celková genetická informace komunity organismů celková genetická informace jednoho organismu
 pouze ty u kterých je znám homologický genom (stejného biologického druhu) Metagenomika. studium biologické rozmanitosti a genetického materiálu celé komunity (mikro)organismů přímo v jejich přirozeném prostředí bez nutnosti jejich kultivace v laboratoři (např. z půdy, střevní mikroflora) Metagenom. Genom. X. celková genetická informace komunity organismů. celková genetická informace jednoho organismu.")
68
Střevní mikroflóra P. J. Turnbaugh et al. (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. Dec 21;444(7122): Apendikální mikrobiální DNA z ob/ob, ob/+, +/+ myši Složení mikroflóry střeva závislé na obezitě změny v zastoupení bakteriálních kmenů Bacteroidetes a Firmicutes změny v metabolickém potenciálu střeva (ukládání energie)
 An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. Dec 21;444(7122): Apendikální mikrobiální DNA z ob/ob, ob/+, +/+ myši. Složení mikroflóry střeva závislé na obezitě. změny v zastoupení bakteriálních kmenů Bacteroidetes a Firmicutes. změny v metabolickém potenciálu střeva (ukládání energie)")
69
DNA mamuta - Mammoth Project
Hendrik N. Poinar et al (2006): Metagenomics to Paleogenomics: Large-Scale Sequencing of Mammoth DNA. Science Jan. 20, 311 (5759): DNA extrahována z 1 g mamutí kosti (28000 let, Siberia) – 100 ml DNA fragmenty 500 – 700 bp „454“ sekvenování 28 miliónů bp čtení 45,4% čtení se přirovnalo se sekvencí slona afrického, 1,4% - s lidskou a 1,2% se psí sekvencí.
: Metagenomics to Paleogenomics: Large-Scale Sequencing of Mammoth DNA. Science Jan. 20, 311 (5759): DNA extrahována z 1 g mamutí kosti (28000 let, Siberia) – 100 ml. DNA fragmenty 500 – 700 bp. „454 sekvenování. 28 miliónů bp čtení. 45,4% čtení se přirovnalo se sekvencí slona afrického, 1,4% - s lidskou a 1,2% se psí sekvencí.")
70
největší význam DIAGNOSTIKA
zlevnit přesekvencování lidského genomu na USD (do roku 2015) (nové levnější přístupy: SOLID Applied Biosystems, HELIOS) CANCER GENOME ATLAS TRANSCRIPTOME SEQUENCING (RNA-seq) – výhoda oproti DNA čipům je že 100% postihuje alternativní sestřih a alelickou variabilitu SINGLE MOLECULE SEQUENCING (Helios) – preinplantační diagnostika SOLID PHASE CAPTURE – NimbleGene™ čip vyrobený pro vychytání kodujících sekvencí a regulačních oblastí z rozfragmentované genomické DNA před vlastním vysokoúčinným sekvencováním
 (nové levnější přístupy: SOLID Applied Biosystems, HELIOS) CANCER GENOME ATLAS. TRANSCRIPTOME SEQUENCING (RNA-seq) – výhoda oproti DNA čipům je že 100% postihuje alternativní sestřih a alelickou variabilitu. SINGLE MOLECULE SEQUENCING (Helios) – preinplantační diagnostika. SOLID PHASE CAPTURE – NimbleGene™ čip vyrobený pro vychytání kodujících sekvencí a regulačních oblastí z rozfragmentované genomické DNA před vlastním vysokoúčinným sekvencováním.")
71
největší význam DIAGNOSTIKA
zlevnit přesekvencování lidského genomu na USD (do roku 2015) (nové levnější přístupy: SOLID Applied Biosystem, HELIOS) CANCER GENOME ATLAS TRANSCRIPTOME SEQUENCING (RNA-seq) – výhoda oproti DNA čipům je že 100% postihuje alternativní sestřih a alelickou variabilitu SINGLE MOLECULE SEQUENCING (Helios) – preinplantační diagnostika SOLID PHASE CAPTURE – NimbleGene™ čip vyrobený pro vychytání kodujících sekvencí a regulačních oblastí z rozfragmentované genomické DNA před vlastním vysokoúčinným sekvencováním
 (nové levnější přístupy: SOLID Applied Biosystem, HELIOS) CANCER GENOME ATLAS. TRANSCRIPTOME SEQUENCING (RNA-seq) – výhoda oproti DNA čipům je že 100% postihuje alternativní sestřih a alelickou variabilitu. SINGLE MOLECULE SEQUENCING (Helios) – preinplantační diagnostika. SOLID PHASE CAPTURE – NimbleGene™ čip vyrobený pro vychytání kodujících sekvencí a regulačních oblastí z rozfragmentované genomické DNA před vlastním vysokoúčinným sekvencováním.")
72
DIAGNOSTIKA ChIP-seq (chromatin imuno precipitation) Methyl-seq
detekce CpG oblastí (epigenetika) využívá konverze cytidinu na uridin za pomocí bisulfidu methylovaný cytidin není konvertován
 využívá konverze cytidinu na uridin za pomocí bisulfidu. methylovaný cytidin není konvertován.")
73
cDNA a GENOMOVÉ KNIHOVNY
je to soubor náhodně klonovaných fragmentů genomové DNA nebo cDNA, připravené zpětnou transkripci mRNA, příslušného organismu do vhodného vektoru, ve kterém může být tento soubor DNA klonů uchováván a množen

74
cDNA a GENOMOVÉ KNIHOVNY
slouží především ke hledání nových genů a jejích klonování pro další funkční analýzu knihovny lze sestrojit pro všechny živé organismy, pro vědecky významné existuji komerčně dostupné knihovny genomová knihovna obsahuje veškerou informaci obsaženou v genomu (geny a nekódující sekvence) cDNA knihovna obsahuje pouze geny exprimované v určitém stádiu vývoje organismu popř. ve specifické tkáni či pletivu jako klonovací vektory pro konstrukci knihoven se používají především vektory odvozené od bakteriofága λ, fazmidy, kosmidy nebo YAC a BAC
 cDNA knihovna obsahuje pouze geny exprimované v určitém stádiu vývoje organismu popř. ve specifické tkáni či pletivu. jako klonovací vektory pro konstrukci knihoven se používají především vektory odvozené od bakteriofága λ, fazmidy, kosmidy nebo YAC a BAC.")
75
genomové knihovny lambda FIX® II (Stratagene)
XhoI → Sau3A I → 4-místné GATC částečné štěpení RE zaručuje překryvy (kontigy) substituční vektor Spi+ selekce klonované fragmenty 9-23kb
 substituční vektor. Spi+ selekce. klonované fragmenty 9-23kb.")
76
genomové knihovny lambda FIX® II
Avian Baboon Bacterial Bovine Canine Cephalopod Chicken Drosophila Feline Fungus Gorilla Guinea Pig Hamster Horse Human Lobster Marsupial Monkey Mouse Nematode Plant Porcine Rabbit Rat Salamander Xenopus Yeast Zebrafish Product: Horse Genomic, Leukocytes Library Arabian, adult stallion, heterozygous for immunodeficiency, Vector: Lambda FIX® II vector, Insert Size: kb Arabidopsis thaliana Nicotiana Plumbaginifolia barley corn pea soybean Xenopus (drápatka) zebrafish (první transgenní ryba)
 zebrafish (první transgenní ryba)")
77
cDNA knihovny lambda ZAP® II
inzerční vektor fazmid blue/white screening klonované fragmenty 0-10kb

78
cDNA knihovny lambda ZAP® II
Výhody: vysoká efektivnost fágové infekce plazmid (pBlueskript) se vyštěpuje z fazmidu (s plazmidem se lépe manipuluje a sekvencuje) zvýšené zastoupení málo abundantních transkriptů: pomocí metody SSH (supressive and subtractive hybridization) se sníží množství vysoce abundantních transkriptů (např. transkripty Rubisco tvoří často 10% z celkové rostlinné cDNA) pomocný fág (má mutaci tak, že se sám nereplikuje ale po koinfekci s fazmidem mu umožní vytvořit infekční částice, tím že vytvoří sbalovací proteiny, jejichž genetická informace na klonovaném fazmidu chybí. Navíc nese aparát pro tvorbu ssDNA.
 se vyštěpuje z fazmidu (s plazmidem se lépe manipuluje a sekvencuje) zvýšené zastoupení málo abundantních transkriptů: pomocí metody SSH (supressive and subtractive hybridization) se sníží množství vysoce abundantních transkriptů (např. transkripty Rubisco tvoří často 10% z celkové rostlinné cDNA) pomocný fág (má mutaci tak, že se sám nereplikuje ale po koinfekci s fazmidem mu umožní vytvořit infekční částice, tím že vytvoří sbalovací proteiny, jejichž genetická informace na klonovaném fazmidu chybí. Navíc nese aparát pro tvorbu ssDNA.")
79
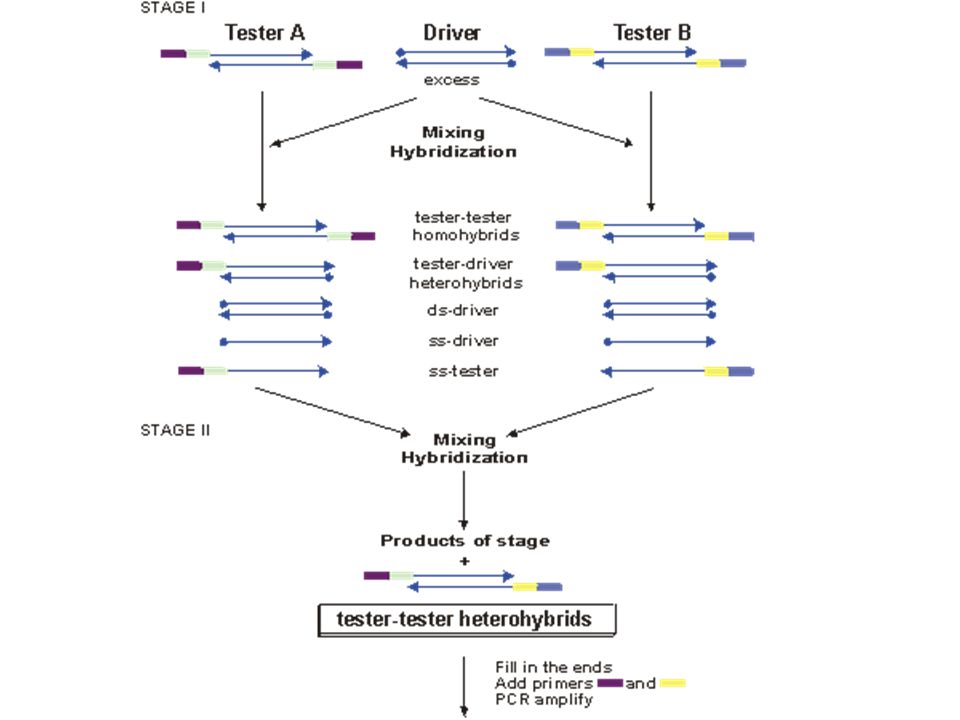
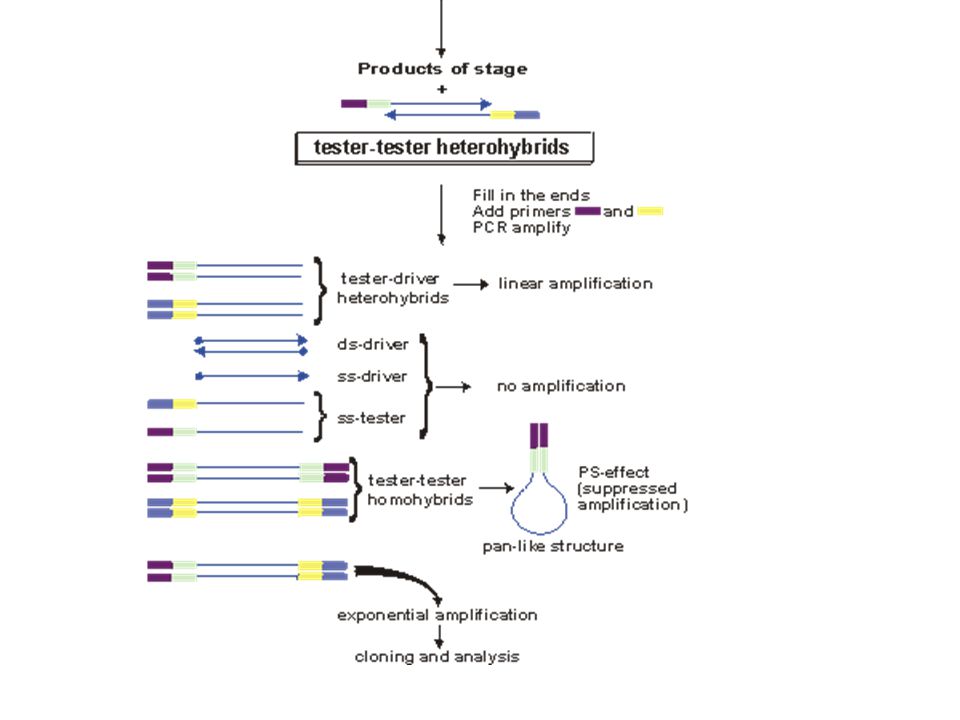
supressive and subtractive hybridization (SSH)
studovaná RNA srovnávací RNA studovaná RNA raritní molekuly zůstávají více nehybridizované hybridizace – kinetika druhého řádu, abundantní molekuly hybridizují velice rychle spolu další využití: zjišťování nebezpečných genů u patogenních organismů

82
EST klony (Expressed Sequence Tags)
Co to je EST? krátká DNA sekvence (okolo bp) odvozená z cDNA reprezentuje geny exprimované ve tkáních ze kterých je odvozena cDNA knihovna - TRANSCRIPTOM pletivově specifické cDNA knihovny listy, kořeny, semena, plody, pyl, květy….. specifické cDNA knihovny po infekci patogenem, po působení hormonů, stresu atd. Výhody EST sekvenci zdroj informaci o genech exprimovaných ve specifických tkáních, nebo v závislosti na odpovědi vůči vnějším vlivům základní srovnávání mezi různými organismy základ pro rozlišování genových rodin a identifikace alelických variací popř. alternativniho sestřihu Nedostatky EST klonů cDNA neobsahuje regulační oblasti a celé kompaktní geny často jenom 3 a 5 konce cDNA klonů ( bp, jedna sekvenační reakce) syrová data, chyby
 odvozená z cDNA. reprezentuje geny exprimované ve tkáních ze kterých je odvozena cDNA knihovna - TRANSCRIPTOM. pletivově specifické cDNA knihovny. listy, kořeny, semena, plody, pyl, květy….. specifické cDNA knihovny. po infekci patogenem, po působení hormonů, stresu atd. Výhody EST sekvenci. zdroj informaci o genech exprimovaných ve specifických tkáních, nebo v závislosti na odpovědi vůči vnějším vlivům. základní srovnávání mezi různými organismy. základ pro rozlišování genových rodin a identifikace alelických variací popř. alternativniho sestřihu. Nedostatky EST klonů. cDNA neobsahuje regulační oblasti a celé kompaktní geny. často jenom 3 a 5 konce cDNA klonů ( bp, jedna sekvenační reakce) syrová data, chyby.")
83
hledání genů in silico v internetových databazích TIGR a GeneBank

84
hledání genů in silico v internetových databazích TIGR a GeneBank
Výsledky hledání v GenBank....

85
počty EST klonů v databázích k roku 2003
výsledek hledání pro polygalakturonasu z rajčete počty EST klonů v databázích k roku 2003 Člověk – přes pět miliónů EST klonů

86
cDNA knihovny lambda ZAP® II
Algae Baboon Bovine Canine Chicken Feline Fish Fungus Hamster Human Fetal Human Human Neuron and Teratocarcinoma Insect Lobster Marsupial Monkey Mouse Nematode Plant Porcine Rabbit Rat Salamander Sheep Xenopus Human cDNA, Aortic Smooth Muscle Cell Library Human cDNA, Brain (Cerebellum) Library Human cDNA, Brain (Frontal cortex) Library Human cDNA, Breast Carcinoma Library Human cDNA, Erythroleukemia cell Library Human cDNA, Heart Library Human cDNA, Kidney Library Human cDNA, Uterus Library (8 pooled normal whole specimens, Caucasian, years, Vector: Uni-ZAP® XR vector, Primer: UdT, Average Insert Size: 1.3 kb) atd. celkem 57 Plant cDNA, Arabidopsis thaliana Library Plant cDNA, Barley Library Plant cDNA, Soybean Library Plant cDNA, Tobacco Leaf Library Plant cDNA, Tomato Library Plant cDNA, Spinach Library (var. Melody, actively growing leaves, Primer: OR, Vector: Lambda ZAP® II vector, Average Insert Size: 1.0 kb)
 Library. Human cDNA, Brain (Frontal cortex) Library. Human cDNA, Breast Carcinoma Library. Human cDNA, Erythroleukemia cell Library. Human cDNA, Heart Library. Human cDNA, Kidney Library. Human cDNA, Uterus Library (8 pooled normal whole specimens, Caucasian, years, Vector: Uni-ZAP® XR vector, Primer: UdT, Average Insert Size: 1.3 kb) atd. celkem 57. Plant cDNA, Arabidopsis thaliana Library. Plant cDNA, Barley Library. Plant cDNA, Soybean Library. Plant cDNA, Tobacco Leaf Library. Plant cDNA, Tomato Library. Plant cDNA, Spinach Library (var. Melody, actively growing leaves, Primer: OR, Vector: Lambda ZAP® II vector, Average Insert Size: 1.0 kb)")
87
otisk plaků na nitrocelulosovou membránu SCREENING cDNA
denaturace DNA na filtru inkubace s hybridizační sondou vyvolání otisku detekce plaku obsahují požadovaný gen vyříznutí a namnožení požadovaného klonu SCREENING cDNA a GENOMOVÝCH KNIHOVEN

88
SCREENING BACových knihoven pomocí „poolování“

89
cDNA a GENOMOVÉ KNIHOVNY
VELIKOST KNIHOVNY vyjadřuje kolik rekombinatních fágu (plaků) musíme screenovat abychom prošli celý genom nebo cDNA pool. P……….pravděpodobnost, že je klon zastoupen n……….poměr průměrné velikosti klonovaného inzertu k velikosti genomu např. při screeningu lidského genomu o velikosti Mbp který je v λ genomové knihovně s velikosti fragmentů 20 kbp musíme na Petriho miskách vypěstovat pro následnou analýzu alespoň 6.5 x 105 plaků abychom s 99% pravděpodobnosti zaručili, že je analyzován celý genom
 musíme screenovat abychom prošli celý genom nebo cDNA pool. P……….pravděpodobnost, že je klon zastoupen. n……….poměr průměrné velikosti klonovaného inzertu k velikosti genomu. např. při screeningu lidského genomu o velikosti Mbp který je v λ genomové knihovně s velikosti fragmentů 20 kbp musíme na Petriho miskách vypěstovat pro následnou analýzu alespoň 6.5 x 105 plaků abychom s 99% pravděpodobnosti zaručili, že je analyzován celý genom.")
90
cDNA a GENOMOVÉ KNIHOVNY
DALŠÍ TYPY KNIHOVEN: EXPRESNÍ GENOMOVÁ KNIHOVNA vychází z cDNA knihovny, která je ale umístěna v expresních vektorech (klonovací vektory, které navíc transgen v nich obsažený exprimují do proteinu) Používají se pro hledaní genů pokud známe pouze protein, který kódují (pomocí protilátky) CHROMOZOMOVÉ KNIHOVNY obsahují informaci pouze z jednoho chromozomu (jsou menší) HYBRIDNÍ KNIHOVNY systém pro studium interakce protein - protein, bait vektor (návnada) obsahuje gen pro protein, který studujeme a fish (prey) vektor (obsahuje danou cDNA expresní knihovnu) s pomocí genomových knihoven se uskutečnila sekvenace celých genomů několika organizmů. díky tomu ale genomové knihovny v současné době ztrácejí na významu (veškerá genetická informace je obsažena in silico a pomocí vhodných primerů si lze kdykoli daný úsek naamplifikovat
 Používají se pro hledaní genů pokud známe pouze protein, který kódují (pomocí protilátky) CHROMOZOMOVÉ KNIHOVNY. obsahují informaci pouze z jednoho chromozomu (jsou menší) HYBRIDNÍ KNIHOVNY. systém pro studium interakce protein - protein, bait vektor (návnada) obsahuje gen pro protein, který studujeme a fish (prey) vektor (obsahuje danou cDNA expresní knihovnu) s pomocí genomových knihoven se uskutečnila sekvenace celých genomů několika organizmů. díky tomu ale genomové knihovny v současné době ztrácejí na významu (veškerá genetická informace je obsažena in silico a pomocí vhodných primerů si lze kdykoli daný úsek naamplifikovat.")
91
YEAST TWO HYBRID SYSTÉM
PRO STUDIUM PROTEIN-PROTEIN INTERAKCÍ – význam pro studium regulačních a signálních drah naklonovaná cDNA knihovna z libovolného euk.organismu konstitutivní promotor kvasinkový chromozom

Podobné prezentace






>")













