Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Neurochemie: Morfologie neuronu (dendrity a axon, synapse), synaptický přenos (skladování, výlev a zpětné vychytávání neuropřenašečů), neuropřenašečové systémy v mozku 2. přednáška

2
Neuron = vysoce polarizovaná komunikační buňka vyskytující se v rámci nervového systému ve velkém množství subtypů, propojených vzájemně do okruhů Morfologicky pak lze v typickém neuronu pozorovat tři hlavní oblasti: tělo neuronu (perikaryon), obsahující jádro a hlavní cytoplasmatické organely různý počet dendritů, které odstupují z perikaryonu, liší se velikostí, tvarem, přítomností dendritických trnů a větví se různě daleko a hustě do šedé hmoty; a jediný axon, který u většiny axonů odstupuje z perikaryonu do daleko větší vzdálenosti než kterákoliv z větví dendritické stromu. každý neuron je vybaven v průměru 7 000 synaptickými spojeními s dalšími neurony tříleté dítě má asi 10 15 synapsí, počet klesá s věkem u dospělého člověka je odhadován na 10 14 až 5 × 10 14 synapsí jiní autoři pro dospělý lidský mozek udávají počet cca 86 miliard neuronů z nich 16,3 miliardy je v kůře mozkové a 69 miliard v mozečku předpokládaná distribuce neuronů a non- neuronálních buněk v lidském mozku: všimněte si, že majorita neuronů se nachází v mozečku, kde je jich cca 4 více než non- neuronálních buněk, zatímco v kortexu je podle této práce poměr neuronů a non- neuronálních buněk opačný.

3
Včela 960 000 neuronů asi 10 9 synapsí Hlemýžď zahradní 11 000 neuronů Mravenec 250 000 neuronů Medůza 800 neuronů Šváb 10 6 neuronů Žába 16 x 10 6 neuronů Chobotnice 300 x 10 6 neuronů Krysa 200 x 10 6 neuronů asi 448 x 10 9 synapsí Kočka 10 9 neuronů asi 10 12-13 synapsí Myš 75 x 10 6 neuronů asi 10 11 synapsí Šimpanz 6,7 x 10 9 neuronů Slon 23 x 10 9 neuronů

4
u primátů existuje lineární závislost velikosti mozku na počtu neuronů větší savci mají obvykle větší mozky ? Gorila dorůstá trojnásobné tělesné hmoty člověka, ale mozek má cca 3x menší ? 1 neuron = asi 25 kJ K udržení velikosti mozku odpovídající 2% tělesné hmotnosti je potřeba značného množství energie. Energetická výtěžnost „hrubé“ stravy není vysoká delší doba krmení se. Proc Natl Acad Sci U S A. 2012 Nov 6;109(45):18571-6 Zatímco gorila stráví 80% 12h dne krmením se, Homo erectus si začal vařit
: Zatímco gorila stráví 80% 12h dne krmením se, Homo erectus si začal vařit.")
5
Časová škála vlivu různých efektů na objem mozkovny Proc Biol Sci. 2011 Apr 7;278(1708):961-9.
:961-9.")
6
Encefalizační koeficient = poměr mezi velikostí mozku a celkovou (předpokládanou) velikostí těla
 velikostí těla")
7
Eric Abelson, ekolog a konzervační biolog, Stanford University - měření více než 1650 zvířat - 160 různých druhů - zvířata vyššího stupně encefalizace jsou více ohrožena vyhynutím - toto riziko je největší pro živočichy malé velikosti těla a k ní relativně velkého mozku - zatím bez vysvětlení Přehled porovnávaných druhů: http://rspb.royalsocietypublishing.org/content/royprsb/suppl/2016/02/16/rspb.2015.2772.DC1/rspb20152772supp1.pdf Opravdu se velký mozek vyplatí?

8
velikost perikaryonu se může mezi jednotlivými neuronálními subtypy značně lišit (rozmezí cca 5-100 µm) perikaryon je bohatý na cisterny endoplasmatického retikula, proloženými řetězci volných polyribosomů označovaných jako Nisslova substance a má bohatý cytoskelet, který ve vřetenech vybíhá do dendritů a axonů Podle počtu z těla neuronu odstupujících výběžků lze morfologicky dělit neurony na unipolární, bipolární či multipolární: dendrit a axon unipolárních (pseudounipolárních) neuronů odstupuje ze stejného výběžku; u bipolárních neuronů odstupují axon a jeden dendrit na opačných koncích perikaryonu; multipolární neurony mají více než dva dendrity a morfologicky se dělí na -multipolární neurony Golgiho typu I s dlouhým axonem (pyramidové buňky, Purkyněho buňky, buňky předních rohů míšních) -multipolární neurony Golgiho typu II, jejichž axony projikují lokálně (granulární buňky)
 perikaryon je bohatý na cisterny endoplasmatického retikula, proloženými řetězci volných polyribosomů označovaných jako Nisslova substance a má bohatý cytoskelet, který ve vřetenech vybíhá do dendritů a axonů Podle počtu z těla neuronu odstupujících výběžků lze morfologicky dělit neurony na unipolární, bipolární či multipolární: dendrit a axon unipolárních (pseudounipolárních) neuronů odstupuje ze stejného výběžku; u bipolárních neuronů odstupují axon a jeden dendrit na opačných koncích perikaryonu; multipolární neurony mají více než dva dendrity a morfologicky se dělí na -multipolární neurony Golgiho typu I s dlouhým axonem (pyramidové buňky, Purkyněho buňky, buňky předních rohů míšních) -multipolární neurony Golgiho typu II, jejichž axony projikují lokálně (granulární buňky)")
9
morfologie axonu a jeho cesty nervovou tkání závisí na typu informace, kterou přenáší, a na propojení neuronu s dalšími neurony odstupuje v místě nazývaném axonální hrbolek (axon hillock); tato struktura je zvláště dobře patrná u velkých yramidových buněk axon bývá v případě dlouhých projekcí myelinizovaný pokud zasahuje jen do blízkého okolí neuronu, myelinovou pochvu mít nemusí Axon Axon vs. dendrit (i)Průměr axonu je po celé délce víceméně konstantní, oproti tomu dendritů klesá směrem od hlavní větve k větvím vyššího řádu. (ii)Dendrity obsahují organely, které axon nemá. (iii)Dendrity a axon lze odlišit díky specifickým axonálním a dendritickým proteinům. často jen vybíhá z jednoho z hlavních dendritů axony obvykle obsahují více neurofilament než dendrity často značně rozvětvené – např. neurony propojující různé kortikální oblasti větvit se ale nemusí: kortikální motorické neurony mají axon v podstatě přímě spojující subkortikální centra s předními rohy míšními
Průměr axonu je po celé délce víceméně konstantní, oproti tomu dendritů klesá směrem od hlavní větve k větvím vyššího řádu. (ii)Dendrity obsahují organely, které axon nemá. (iii)Dendrity a axon lze odlišit díky specifickým axonálním a dendritickým proteinům. často jen vybíhá z jednoho z hlavních dendritů axony obvykle obsahují více neurofilament než dendrity často značně rozvětvené – např. neurony propojující různé kortikální oblasti větvit se ale nemusí: kortikální motorické neurony mají axon v podstatě přímě spojující subkortikální centra s předními rohy míšními.")
10
Axony jsou zodpovědné za přenos informace zpracované neurony. synapse = rozhraní mezi axonálním zakončením (terminálou, terminální arborizací) a cílovou buňkou synapse = specializovaný útvar obecně složený s presynaptického (axonálního) elementu, úzké synaptické štěrbiny a elementu postsynaptického. Podle polohy elementu pre- a postsynaptického se obvykle rozlišují axodendritické či axosomatické synapse: presynaptický element inervuje dendrity nebo tělo jiného neuronu; axoaxonální synapse: neuropřenašeč se vylévá poblíž jiné nervové terminály; autoreceptorové „synapse“: leží na tom samém nervovém zakončení, z něhož se neuropřenašeč vylévá; dendroaxonální synapse: neuropřenašeč uvolněný z dendritů či těla neuronu ovlivňuje zakončení axonu neuronu jiného (vzácné); dendrodenritické synapse: vyskytující se např. v čichovém laloku či sítnici někdy mohou vzniknout synapse i mezi dvěma perikaryony; zakončení en passant (synapse en passant): spoje „mimochodem“ se utvářející při průchodu axonu nad dendritickým stromem jiného postsynaptického neuronu
 a cílovou buňkou synapse = specializovaný útvar obecně složený s presynaptického (axonálního) elementu, úzké synaptické štěrbiny a elementu postsynaptického. Podle polohy elementu pre- a postsynaptického se obvykle rozlišují axodendritické či axosomatické synapse: presynaptický element inervuje dendrity nebo tělo jiného neuronu; axoaxonální synapse: neuropřenašeč se vylévá poblíž jiné nervové terminály; autoreceptorové „synapse : leží na tom samém nervovém zakončení, z něhož se neuropřenašeč vylévá; dendroaxonální synapse: neuropřenašeč uvolněný z dendritů či těla neuronu ovlivňuje zakončení axonu neuronu jiného (vzácné); dendrodenritické synapse: vyskytující se např. v čichovém laloku či sítnici někdy mohou vzniknout synapse i mezi dvěma perikaryony; zakončení en passant (synapse en passant): spoje „mimochodem se utvářející při průchodu axonu nad dendritickým stromem jiného postsynaptického neuronu.")
11
Dendrity ( δένδρον [déndron ] - strom) = mnohočetné jemné výběžky odstupující z těla neuronu a spolu s perikaryonem představující primární recepční strukturu dostávají informace ze synaptických vstupů jiných neuronů kromě recepce též procesují a integrují tyto vstupní informace větví se ve výběžky vyšších řádů, přičemž vytvářejí tzv. dendritický strom geometrie dendritického stromu může být velice komplexní a často velmi půvabná poloha určitého dendritu v rámci dendritického stromu determinuje i jeho příspěvek k celkovému informačnímu vstupu: pokud je „vybaven“ přibližně stejným množstvím stejných synapsí jako jiný dendrit, tak čím dále od integrujícího místa leží, tím je jeho příspěvek nižší než příspěvek stejně „vybaveného“ dendritu, odstupujícího blíže u těla neuronu
![Dendrity ( δένδρον [déndron ] - strom) = mnohočetné jemné výběžky odstupující z těla neuronu a spolu s perikaryonem představující primární recepční strukturu dostávají informace ze synaptických vstupů jiných neuronů kromě recepce též procesují a integrují tyto vstupní informace větví se ve výběžky vyšších řádů, přičemž vytvářejí tzv.](http://images.slideplayer.cz/46/11637446/slides/slide_11.jpg "dendritický strom geometrie dendritického stromu může být velice komplexní a často velmi půvabná poloha určitého dendritu v rámci dendritického stromu determinuje i jeho příspěvek k celkovému informačnímu vstupu: pokud je „vybaven přibližně stejným množstvím stejných synapsí jako jiný dendrit, tak čím dále od integrujícího místa leží, tím je jeho příspěvek nižší než příspěvek stejně „vybaveného dendritu, odstupujícího blíže u těla neuronu.")
12
Pyramidové buňky z různých korových oblastí a jejich bazální a apikální dendritická síť a | Layer V pyramidal neurons have longer apical dendrites and fewer oblique apical dendrites than layer II/III pyramidal neurons. The apical dendrites of hippocampal CA3 pyramidal neurons branch closer to the soma than those of CA1 pyramidal neurons, which typically have a more distinctive main apical dendrite and tuft. CA3 pyramidal neurons also have a cluster of large spines in the first 100 mum of the apical dendrite. The full vertical length of each cell, from left to right, is: unknown; 1,180 m; 580 m; 730 m; 790 m. All cells are from the rat, except for the layer II/III cell, which is from the rabbit. b | Apikální dendrity /fialové pozadí/ pyramidových buněk dostávají excitační synaptické vstupy z různých presynaptických oblastí oproti dendritům proximálním nebo bazálním /zelené pozadí /. Bazální a proximální dendrity korové vrstvy II/III dostávají vstupy z vrstvy IV a z lokálních excitačních obvodů. Apikální dendrity vrstev II/III dostávají vstupy z jiných korových oblastí a nespecifické vstupy z thalamu. Podobné uspořádání lze pozorovat i ve vrstvě pyramidových buněk. Bazální a vzdálenější apikální dendrity CA1 pyramidových buněk primárně dostávají vstupy z CA3 buněk, zatímco apikální dendrity z entorhinální kůry a z jader thalamu.. c | Schéma pyramidového neuronu ilustrující různé dendritické domény, které dostávají specifické a jedinečné synaptické vstupy. Různé barvy pozadí značí oblasti s různými synpatickými vstupy nebo synapsemi odlišných vlastností.

13
Dendritické trny - malé membránové protruze odstupující z mnoha typů neuronů dostávají zejména excitační synaptické vstupy určené jejich neuronu, ale mohou na hlavičce obsahovat i aditivní synaptické vstupy mohou strukturálně obklopovat a biochemicky izolovat danou synapsi; takto „informačně nejoddělenější“ jsou dlouhé, tenké dendritické trny dendritické trny se nacházejí na všech hlavních neuronálních typech v CNS (kortikální neurony, Purkyněho buňky atd.) dendrity jednoho neuronu mohou souhrnně obsahovat stovky až tisíce trnů typická hustota trnů až 20 (50) trnů na 10 µm délky dendritu dendritické trny kortikálních neuronů mohou obsahovat až desítky tisíc synapsí, Purkyněho buňky i o řád více dendritické trny obsahují 90% synapsí CNS Trny jsou útvary složené z „hlavičky“ a nožky, která je spojuje s dendritem. objem hlavičky je obvykle 0,01-0,8 µm 3 podle tvaru hlavičky se trny obvykle dělí na "tenké - thin", "pahýlovité (useknuté) - stubby", "hřibovité - mushroom", a "větvené - branched„ jejich membrána bývá vybavena AMPA či NMDA receptory (excitační transmise) obsahují i TrkB receptor pro BDNF (udržení trnu na dendritu
 - stubby , hřibovité - mushroom , a větvené - branched„ jejich membrána bývá vybavena AMPA či NMDA receptory (excitační transmise) obsahují i TrkB receptor pro BDNF (udržení trnu na dendritu.")
14
a | Two spine-studded dendrites of a stained CA1 pyramidal neuron. The higher density of spines in the right-hand dendritic segment resulted from estrogen treatment45, suggesting that spines and synapses might respond dynamically to changes in circulating hormones. b | A dendrite that has been labelled for microtubule-associated protein 2 (MAP2; red) and actin (green). MAP2 is concentrated in the dendritic shafts. Actin filaments in the spine head mediate spine motility. c | A three-dimensional reconstruction of spines and synapses in a typical pyramidal cell, based on electron micrographs of a single stretch of dendrite from a filled cell. Every spine is contacted by at least one synapse. The dendrite and its spines are shown in grey; synaptic boutons forming single synapses are shown in blue; boutons forming multiple synapses onto more than one cell are shown in green; boutons forming multiple synapses onto the same cell are shown in yellow; spines from other dendrites are shown in orange. d | The left-hand panels show electron micrographs of two non-perforated postsynaptic densities (PSDs; top panel, indicated by arrows) and a perforated PSD (bottom panel, the perforation is indicated by the large arrow). The right-hand panel contains schematic diagrams of a non-perforated synapse (top) and a perforated synapse (bottom). e | Two-photon glutamate uncaging at various locations on a dendritic segment. The colours indicate the somatically recorded current amplitude that was measured when uncaging was carried out at each location. Note that the largest response (yellow/orange) occurred when the glutamate was uncaged on a large spine head.
 and actin (green). MAP2 is concentrated in the dendritic shafts. Actin filaments in the spine head mediate spine motility. c | A three-dimensional reconstruction of spines and synapses in a typical pyramidal cell, based on electron micrographs of a single stretch of dendrite from a filled cell. Every spine is contacted by at least one synapse. The dendrite and its spines are shown in grey; synaptic boutons forming single synapses are shown in blue; boutons forming multiple synapses onto more than one cell are shown in green; boutons forming multiple synapses onto the same cell are shown in yellow; spines from other dendrites are shown in orange. d | The left-hand panels show electron micrographs of two non-perforated postsynaptic densities (PSDs; top panel, indicated by arrows) and a perforated PSD (bottom panel, the perforation is indicated by the large arrow). The right-hand panel contains schematic diagrams of a non-perforated synapse (top) and a perforated synapse (bottom). e | Two-photon glutamate uncaging at various locations on a dendritic segment. The colours indicate the somatically recorded current amplitude that was measured when uncaging was carried out at each location. Note that the largest response (yellow/orange) occurred when the glutamate was uncaged on a large spine head..")
15
Plasticita dendritických trnů: vejmi výrazně mění svůj tvar, objem a počet v relativně krátkém časovém úseku remodelace v řádu vteřin až minut díky aktinovému cytoskeletu počet dendritických trnů se výrazně mění v čase: 10-20% trnů v pyramidových buňkách kortexu může vznikat a zanikat v řádu hodin „hřibovité“ trny jsou nejstabilnější. tvorba paměťových stop: růst nových dendritických trnů či prodlužování stávajících, aby byla posílena schopnost presynaptického neuronu aktivovat ten postsynaptický a aby došlo k posílení synaptického kontaktu Počet a tvar dendritických trnů kolísá i v závislosti na různých patofyziologických stavech. Syndrom fragilního chromosomu X: jedním z projevů je mentální retardace, pravděpodobně podložená chyběním proteinu FMRP (fragile X mental retardation protein). díky absenci tohoto proteinu vzniká neúměrně mnoho dendritických trnů FMRP normálně funguje jako „brzda“ vzniku nových synapsí a vyvažuje „posilující“ efekt metabotropních glutamátových receptorů (mGluR 5 ), které vznik synapsí na trnech potencují /trny zdravé myši (vlevo), myši s nedostatkem FMRP (uprostřed) a myši rovněž knock-outované pro FMRP, které byl ovšem uměle redukován i počet mGluR 5, takže nedošlo tak dramatickému nárostu počtu dendritických trnů jako při prostém knock-outu pro FMRP/
. díky absenci tohoto proteinu vzniká neúměrně mnoho dendritických trnů FMRP normálně funguje jako „brzda vzniku nových synapsí a vyvažuje „posilující efekt metabotropních glutamátových receptorů (mGluR 5 ), které vznik synapsí na trnech potencují /trny zdravé myši (vlevo), myši s nedostatkem FMRP (uprostřed) a myši rovněž knock-outované pro FMRP, které byl ovšem uměle redukován i počet mGluR 5, takže nedošlo tak dramatickému nárostu počtu dendritických trnů jako při prostém knock-outu pro FMRP/.")
16
Epileptiformní aktivita: redukce velikosti dendritických trnů. Na obrázku vidíte rekonstrukci dendritické větve před a po záchvatu, dole v grafu si prohlédněte poměrně výraznou redukci délky vybraného dendritického trnu. Zvýšení počtu dendritických trnů: posiluje LTP v hippokampu (Duffy et al. 2001, ale kontra: Lang et al. 2004, Zhou et al. 2004) zvyšuje kognitviní výkon a učení (Hymovitch 1952, Brown 1968, Gardner et al. 1975, Rampon et al. 2000, Duffy et al. 2001; reviewed by Lewis 2004, Leggio et al. 2005) ovlivňuje genovou expresi (Rampon et al. 2000, Lazarov et al. 2005) ovlivňuje synaptogenesi (Greenough et al. 1985, Rampon et al. 2000, Briones et al. 2004) a morfologii neuronu (Schapiro & Vukovich 1970, Volkmar & Greenough 1972, Globus et al. 1973, Greenough et al. 1978, Kozorovitskiy et al. 2005, Leggio et al. 2005) ovlivňuje proliferaci buněk a jejich přežití (Altman & Das 1964, Szeligo & Leblond 1977, van Praag et al. 1999) může souviset s některými neuropsychiatrickými onemocněními (Penzes et al., Dendritic spine pathology in neuropsychiatric disorders, Nature Neuroscience 14, 285–293, 2011 – hezká práce)
 zvyšuje kognitviní výkon a učení (Hymovitch 1952, Brown 1968, Gardner et al. 1975, Rampon et al. 2000, Duffy et al. 2001; reviewed by Lewis 2004, Leggio et al. 2005) ovlivňuje genovou expresi (Rampon et al. 2000, Lazarov et al. 2005) ovlivňuje synaptogenesi (Greenough et al. 1985, Rampon et al. 2000, Briones et al. 2004) a morfologii neuronu (Schapiro & Vukovich 1970, Volkmar & Greenough 1972, Globus et al. 1973, Greenough et al. 1978, Kozorovitskiy et al. 2005, Leggio et al. 2005) ovlivňuje proliferaci buněk a jejich přežití (Altman & Das 1964, Szeligo & Leblond 1977, van Praag et al. 1999) může souviset s některými neuropsychiatrickými onemocněními (Penzes et al., Dendritic spine pathology in neuropsychiatric disorders, Nature Neuroscience 14, 285–293, 2011 – hezká práce).")
17
Bars across the top indicate the period of emergence of symptoms and diagnosis. In normal subjects, spine numbers increase before and after birth; spines are selectively eliminated during childhood and adolescence to adult levels. In ASD, exaggerated spine formation or incomplete pruning may occur in childhood leading to increased spine numbers. In schizophrenia, exaggerated spine pruning during late childhood or adolescence may lead to the emergence of symptoms during these periods. In Alzheimer's disease, spines are rapidly lost in late adulthood, suggesting perturbed spine maintenance mechanisms that may underlie cognitive decline.

18
CITIZEN (NEURO)SCIENCE: http://www.openconnect omeproject.org/ http://www.eyewire.org/ ! http://blog.eyewire.org/eyewires-first-scientific-discovery-and- nature-paper/ Nature. 2014 May 15;509(7500):331-6.
:")
19
„Příjem“ chemické informace ze sousedních i vzdálenějších neuronů se odehrává primárně na dendritech a jejich trnech. Synaptický přenos je proces přenosu signálu, které se odehrává ve třech krocích: 1. elektrická informace v axonu presynaptické buňky je konvertována na nervovém zakončení axonu v informaci chemickou 2. tento chemický signál je předán jiné buňce 3. chemický signál obdržený postsynaptickou buňkou je opět přeměněn v signál elektrický, který se šíří dále.

20
Synaptický přenos se odehrává na rozhraní mezi presynaptickou a postsynaptickou buňkou na subcelulární struktuře nazývané synapse. Synapse (ex synaptein, z řecké předpony syn- (spolu) a slovesa haptein – spojit): termín navrhl jako modifikaci termínu syndesm Sira Charlese Scotta Sherringtona (1897) Verral synapse jako element složený ze tří částí (Ramón y Cajal) průměrný počet spojů vytvářených jedním neuronem je asi 7 000 typická Purkyňova buňka mozečku: více než 100 000 synapsí malé kulovité a v elektronovém mikroskopu jevící se jako průhledné váčky o průměru asi 50 nm: zpravidla acetylcholin, glutamát a podobné nízkomolekulární neuropřenašeče malé průhledné váčky oválnějšího tvaru: zpravidla obsahují inhibiční neuropřenašeče (glycin) velké měchýřky s průměrem nad 60 nm, často s denzními jádry: katecholaminy největší synaptické váčky s průměry okolo 175 nm: peptidy většina synaptických váčků obsahuje více než jen jeden neuropřenašeč; i měchýřky naplněné acetylcholinem nebo glycinem obsahují dále ATP a rozpustné proteiny. Katecholaminové váčky obsahují kromě ATP i ATPasy a dopamin- -hydroxylasu. Velké váčky peptidergních neuronů: ATP, ATPasa, adenylycyklasa, Ca 2+ ionty a zřejmě také některými enzymy potřebnými pro konečné stadium posttranslačních úprav
 a slovesa haptein – spojit): termín navrhl jako modifikaci termínu syndesm Sira Charlese Scotta Sherringtona (1897) Verral synapse jako element složený ze tří částí (Ramón y Cajal) průměrný počet spojů vytvářených jedním neuronem je asi typická Purkyňova buňka mozečku: více než synapsí malé kulovité a v elektronovém mikroskopu jevící se jako průhledné váčky o průměru asi 50 nm: zpravidla acetylcholin, glutamát a podobné nízkomolekulární neuropřenašeče malé průhledné váčky oválnějšího tvaru: zpravidla obsahují inhibiční neuropřenašeče (glycin) velké měchýřky s průměrem nad 60 nm, často s denzními jádry: katecholaminy největší synaptické váčky s průměry okolo 175 nm: peptidy většina synaptických váčků obsahuje více než jen jeden neuropřenašeč; i měchýřky naplněné acetylcholinem nebo glycinem obsahují dále ATP a rozpustné proteiny. Katecholaminové váčky obsahují kromě ATP i ATPasy a dopamin- -hydroxylasu. Velké váčky peptidergních neuronů: ATP, ATPasa, adenylycyklasa, Ca 2+ ionty a zřejmě také některými enzymy potřebnými pro konečné stadium posttranslačních úprav.")
21
A)plnění váčku nízkomolekulárními látkami typu acetylcholinu: transportéry neuropřenašečů. = velké proteiny tvořené 12 transmembránovými smyčkami (12TM proteiny), dělené do 5 tříd: transportér pro acetylcholin (VAChT, vesicular ACh transporter), transportér pro katecholaminy, transportér pro glutamát a transportér pro GABA a glycin transportér pro adenosin Jejich specifita zajišťuje naplnění váčků příslušným neuropřenašečem. Princip: protonová pumpa (ATPasa energii spotřebovávající děj) čerpá protony do měchýřku na membráně měchýřku vznikne energetický gradient pokud je membrána váčku propustná pro difusibilní anionty jako Cl -, ustaví se také gradient elektrochemický při zpětném „úniku“ protonů do cytoplasmy je pak energetický rozdíl vytvořený jejich předchozím načerpáním do váčku využit na transport neuropřenašeče ATP ADP+P i H+H+ NTH+H+ + + +_ _ + + + _ _ _ _ Životní cyklus váčků na nervovém zakončení: naplnění výlev vtažení zpět do terminály
, dělené do 5 tříd: transportér pro acetylcholin (VAChT, vesicular ACh transporter), transportér pro katecholaminy, transportér pro glutamát a transportér pro GABA a glycin transportér pro adenosin Jejich specifita zajišťuje naplnění váčků příslušným neuropřenašečem. Princip: protonová pumpa (ATPasa energii spotřebovávající děj) čerpá protony do měchýřku na membráně měchýřku vznikne energetický gradient pokud je membrána váčku propustná pro difusibilní anionty jako Cl -, ustaví se také gradient elektrochemický při zpětném „úniku protonů do cytoplasmy je pak energetický rozdíl vytvořený jejich předchozím načerpáním do váčku využit na transport neuropřenašeče ATP ADP+P i H+H+ NTH+H _ _ _ _ _ _ Životní cyklus váčků na nervovém zakončení: naplnění výlev vtažení zpět do terminály.")
22
B) výlev váčku C) synaptický váček vylije svůj obsah do synaptické štěrbiny je vtažen zpět do cytoplasmy dochází k jeho znovunaplnění neurotransmitery buď rovnou v terminále (v případě ACh a podobných nízkomolekulárních látek), nebo se váčky „spojí“ do tzv. multivesikulárních tělísek odtransportovány zpět do perikaryonu tam znovunaplněny (např. neuropeptidy). * Glycine transporters, including: o Glycine transporter type 1 (GlyT1) o Glycine transporter type 2 (GlyT2) * Monoamine transporters, including: o Dopamine transporter (DAT) o Norepinephrine transporter (NET) o Serotonin transporter (SERT) o Vesicular monoamine transporter 1 (VMAT1) o Vesicular monoamine transporter 2 (VMAT2) * Adenosine transporters, including: o Equilibrative nucleoside transporter 1 (ENT1) o Equilibrative nucleoside transporter 2 (ENT2) o Equilibrative nucleoside transporter 3 (ENT3) o Equilibrative nucleoside transporter 4 (ENT4) * Vesicular acetylcholine transporter (VAChT) * Glutamate/aspartate transporters, including: o Excitatory amino acid transporter 1 (EAAT1) o Excitatory amino acid transporter 2 (EAAT2) o Excitatory amino acid transporter 3 (EAAT3) o Excitatory amino acid transporter 4 (EAAT4) o Excitatory amino acid transporter 5 (EAAT5) o Vesicular glutamate transporter 1 (VGLUT1) o Vesicular glutamate transporter 2 (VGLUT2) o Vesicular glutamate transporter 3 (VGLUT3) * GABA transporters, including: o GABA transporter type 1 (GAT1) o GABA transporter type 2 (GAT2) o GABA transporter type 3 (GAT3) o Betaine transporter (BGT1) o Vesicular GABA transporter (VGAT) Přehled typů transportérů neuropřenašečů
. * Glycine transporters, including: o Glycine transporter type 1 (GlyT1) o Glycine transporter type 2 (GlyT2) * Monoamine transporters, including: o Dopamine transporter (DAT) o Norepinephrine transporter (NET) o Serotonin transporter (SERT) o Vesicular monoamine transporter 1 (VMAT1) o Vesicular monoamine transporter 2 (VMAT2) * Adenosine transporters, including: o Equilibrative nucleoside transporter 1 (ENT1) o Equilibrative nucleoside transporter 2 (ENT2) o Equilibrative nucleoside transporter 3 (ENT3) o Equilibrative nucleoside transporter 4 (ENT4) * Vesicular acetylcholine transporter (VAChT) * Glutamate/aspartate transporters, including: o Excitatory amino acid transporter 1 (EAAT1) o Excitatory amino acid transporter 2 (EAAT2) o Excitatory amino acid transporter 3 (EAAT3) o Excitatory amino acid transporter 4 (EAAT4) o Excitatory amino acid transporter 5 (EAAT5) o Vesicular glutamate transporter 1 (VGLUT1) o Vesicular glutamate transporter 2 (VGLUT2) o Vesicular glutamate transporter 3 (VGLUT3) * GABA transporters, including: o GABA transporter type 1 (GAT1) o GABA transporter type 2 (GAT2) o GABA transporter type 3 (GAT3) o Betaine transporter (BGT1) o Vesicular GABA transporter (VGAT) Přehled typů transportérů neuropřenašečů.")
23
Co je to neuropřenašeč? první nároky stanoveny na základě vlastností acetylcholinu: neuropřenašeč je syntetizován v neuronu, uvolňován na jeho zakončení, pokud je podán uměle, vyvolává odpověď, jejíž charakteristiky jsou velmi podobné klasickému přenosu signálu, a konečně je nějakým způsobem inaktivován, pokud možno enzymem vázáným na postsynaptické membráně Smith (2003) uvádí tato kriteria pro definování neuropřenašeče: (i)Molekula musí být syntetizována v neuronu, ze kterého je uvolňována. Enzymy a substráty nutné pro tuto syntézu musí být v neuronu detekovány. (ii)Molekula musí být skladována v neuronu, ze kterého je uvolňována. (iii)Presynaptická stimulace (obvykle elektrická) musí vést k uvolnění této molekuly. (iv)Aplikace této molekuly za definovaných podmínek na patřičné místo (synapsi) musí vyvolat tutéž odpověď jako presynaptická stimulace. (v)Látky blokující postsynaptickou odpověď (např. blokátory kanálů na postsynaptické membráně) na presynaptickou stimulaci musí také blokovat odpověď vyvolanou exogenním podáním látky (bod iv). (vi)Postsynaptická odpověď na tuto molekulu musí být krátká a definovaná. (vii)Musí mít stejné farmakologické charakteristiky jako endogenní neurotransmiter (např. potenciace, inhibice, inaktivace). neuropřenašeč … signální molekula, která reguluje elektrochemický stav přilehlých buněk
 uvádí tato kriteria pro definování neuropřenašeče: (i)Molekula musí být syntetizována v neuronu, ze kterého je uvolňována. Enzymy a substráty nutné pro tuto syntézu musí být v neuronu detekovány. (ii)Molekula musí být skladována v neuronu, ze kterého je uvolňována. (iii)Presynaptická stimulace (obvykle elektrická) musí vést k uvolnění této molekuly. (iv)Aplikace této molekuly za definovaných podmínek na patřičné místo (synapsi) musí vyvolat tutéž odpověď jako presynaptická stimulace. (v)Látky blokující postsynaptickou odpověď (např. blokátory kanálů na postsynaptické membráně) na presynaptickou stimulaci musí také blokovat odpověď vyvolanou exogenním podáním látky (bod iv). (vi)Postsynaptická odpověď na tuto molekulu musí být krátká a definovaná. (vii)Musí mít stejné farmakologické charakteristiky jako endogenní neurotransmiter (např. potenciace, inhibice, inaktivace). neuropřenašeč … signální molekula, která reguluje elektrochemický stav přilehlých buněk.")
24
Roku 1903 Otto Loewi předeslal, že nervová zakončení mohou obsahovat chemické látky z nich uvolňované po stimulaci, a že tyto látky mohou stimulovat patřičný efektorový orgán. V té době ale nevěděl, jak svou hypotézu prokázat. Až roku 1920 se mu ve snu zjevilo schéma zásadního experimentu: "The night before Easter Sunday of that year I awoke, turned on the light, and jotted down a few notes on a tiny slip of thin paper. Then I felt asleep again. It occurred to me at six o'clock in the morning that I had written down something most importnat, but I was unable to decipher the scrawl. The next night, at three o'clock, the idea returned. It was the design of an experiment to determine whether or not the hypothesis of chemical transmission that I had uttered seventeen years ago was correct. I got up immediately, went to the laboratory, and performed a simple experiment on a frog heart according to the nocturnal design." [...] (Otto Loewi: An Autobiographic Sketch, Univ. of Chicago Press, 1960) "These results unequivocally proved that the nerves do not influence the heart directly but liberate from their terminals specific chemical substances which, in their turn, cause the well-known modifications of the function of the heart characteristic of the stimulation of the heart."
 These results unequivocally proved that the nerves do not influence the heart directly but liberate from their terminals specific chemical substances which, in their turn, cause the well-known modifications of the function of the heart characteristic of the stimulation of the heart. .")
25
Neurotransmitery a neuromodulátory A) nízkomolekulární oxid dusnatý (NO), CO, H 2 S kanabinoidy (anandamid) acetylcholin (ACh) aminokyseliny: glutamát (Glu), aspartát (Asp) glycin (Gly) deriváty aminokyselin: a) tryptofan serotonin b) fenylalanin dopamin, adrenalin, noradrenalin, oktopamin c) glycin taurin d) glutamát -aminomáselná kyselina (GABA) e) histidin histamin b) a) c)d) e) - aspartyl Neuropřenašeče mají přímý účinek na subsynaptickou membránu (mění pravděpodobnost otevření iontového kanálu a podobně). Neuromodulátory upravují účinek neuropřenašečů: mohou měnit citlivost postsynaptické membrány k neuropřenašeči nebo např. mohou ovlivňovat výlev neurotransmiteru na presynaptické membráně.

26
C) látky peptidické povahy opioidní peptidy: , , - endorphin, dynorphin, enkephaliny neurohypofysální peptidy: vasopresin, oxytocin, neurophysiny tachykininy: substance P, neurokinin A (NKA, = substance K, SK), neurokinin B (NKB), kassinin, eledosin gastriny: gastrin, cholecystokininy (CCKs) od glukagonu odvozené: VIP (vasoactive intestinal polypeptide) od pankreatických polypeptidů odvozené: neuropeptid Y (NPY) jiné: bombesin, neurotensin (NT), bradykinin, angiotensin, CGRP (calcitonin gene-related peptide) (Zdroj: Cooper, Bloom and Roth: The Biochemical basis of neuropharmacology, 6th edition, Oxford University Press, New York) Neurotransmitery a neuromodulátory B) Puriny adenosin, AMP, ADP, ATP
 látky peptidické povahy opioidní peptidy: , , - endorphin, dynorphin, enkephaliny neurohypofysální peptidy: vasopresin, oxytocin, neurophysiny tachykininy: substance P, neurokinin A (NKA, = substance K, SK), neurokinin B (NKB), kassinin, eledosin gastriny: gastrin, cholecystokininy (CCKs) od glukagonu odvozené: VIP (vasoactive intestinal polypeptide) od pankreatických polypeptidů odvozené: neuropeptid Y (NPY) jiné: bombesin, neurotensin (NT), bradykinin, angiotensin, CGRP (calcitonin gene-related peptide) (Zdroj: Cooper, Bloom and Roth: The Biochemical basis of neuropharmacology, 6th edition, Oxford University Press, New York) Neurotransmitery a neuromodulátory B) Puriny adenosin, AMP, ADP, ATP")
27
Rothman (90. léta): tzv. SNARE hypothesa SNARE odvozeno z soluble N-ethylmaleimide-sensitive fusion protein attachment protein receptor) schéma interakcí proteinů presynaptické membrány (souhrnně označovaných jako t-SNARE proteiny) a proteinů membrány měchýřku (v-SNARE proteiny; v – vesicle) samotnému výlevu měchýřku podle této hypothesy předchází (vyjma plnění váčku neurotransmiterem) tři kroky: docking (dopravení měchýřku a jeho zakotvení v aktivní zóně) priming (fáze, kdy je váček připraven v aktivní zóně a čeká na spouštěcí signál k exocytose) a samotné spuštění exocytosy influxem Ca 2+ iontů do nervového zakončení procesu exocytosy se účastní i řada proteinů regulačních měchýřky s neuropřenašeči dopraveny na synapsi rychlým axonálním transportem rychlostí 2-4mm/s, tj. asi 20-40 cm za den celková doba exocytosy je méně než 1 ms uvolnění složitějších látek z velkých denzních váčku také netrvá déle než 10 ms. Výlev váčku - exocytosa
 schéma interakcí proteinů presynaptické membrány (souhrnně označovaných jako t-SNARE proteiny) a proteinů membrány měchýřku (v-SNARE proteiny; v – vesicle) samotnému výlevu měchýřku podle této hypothesy předchází (vyjma plnění váčku neurotransmiterem) tři kroky: docking (dopravení měchýřku a jeho zakotvení v aktivní zóně) priming (fáze, kdy je váček připraven v aktivní zóně a čeká na spouštěcí signál k exocytose) a samotné spuštění exocytosy influxem Ca 2+ iontů do nervového zakončení procesu exocytosy se účastní i řada proteinů regulačních měchýřky s neuropřenašeči dopraveny na synapsi rychlým axonálním transportem rychlostí 2-4mm/s, tj. asi cm za den celková doba exocytosy je méně než 1 ms uvolnění složitějších látek z velkých denzních váčku také netrvá déle než 10 ms. Výlev váčku - exocytosa.")
28
ProteinMol. hmotnost (kDa) Vlastnosti/Důleži té oblasti molekuly Funkce VAMP/synaptobrevi n 18 13 cíl BoTX B, D, F a Gv-SNARE protein, docking měchýřků, fúze membrán Synaptotagmin65dvě C2A a C2B domény vápníkový sensor, docking měchýřků Synaptophysin384 centrální transmembránové domény molekulární „zarážka“ regulující kontakt v- a t- SNARE, fúze membrán Synapsin80, fosfoproteininteraguje s měchýřky a aktinem spojuje váčky s cytoskeletem, reguluje exocytosu Rab3A25, protein vážící GTP cykluje mezi membránou váčku a cytoplasmou správná lokalizace a docking váčku, fúze membrán Rabphilin3A86C2 motivy, zinc- finger doména regulace pohybu rab3A v Ca- dependentním modu Cysteine string protein 22centrální doména bohatá na cystein kontransport dalších proteinů (?) Proteiny asociované se synaptickými váčky
 Vlastnosti/Důleži té oblasti molekuly Funkce VAMP/synaptobrevi n cíl BoTX B, D, F a Gv-SNARE protein, docking měchýřků, fúze membrán Synaptotagmin65dvě C2A a C2B domény vápníkový sensor, docking měchýřků Synaptophysin384 centrální transmembránové domény molekulární „zarážka regulující kontakt v- a t- SNARE, fúze membrán Synapsin80, fosfoproteininteraguje s měchýřky a aktinem spojuje váčky s cytoskeletem, reguluje exocytosu Rab3A25, protein vážící GTP cykluje mezi membránou váčku a cytoplasmou správná lokalizace a docking váčku, fúze membrán Rabphilin3A86C2 motivy, zinc- finger doména regulace pohybu rab3A v Ca- dependentním modu Cysteine string protein 22centrální doména bohatá na cystein kontransport dalších proteinů ( ) Proteiny asociované se synaptickými váčky.")
29
ProteinMol. hmotnost (kDa) Vlastnosti/Důležité oblasti molekuly Funkce Syntaxin 35cíl BoTX C1t-SNARE protein, dockin měchýřků, fúze membrán SNAP-2525 23 a 29; 23 kDa predominantní v dospělém NS palmitoylován v oblasti centrální domény fúze membrán, stechiometricky se váže na synaptotagmin v Ca 2+ -závislé fázi výlevu Munc-1865interakce se syntaxinemmolekulární „zarážka“ bránící předčasnému spojeni v- a t-SNARE, docking měchýřků / může výlev i facilitovat Hrs-2115interakce se SNAP-25docking měchýřků, fúze membrán (?) RIMasi 200PDZ a C2 domény, motiv zinkových prstů na N- konci docking měchýřků, fúze membrán (?), regulace Rab3 proteinu ProteinMol. hmotnost (kDa) Vlastnosti/Důležité oblasti molekuly Funkce NSF75homotrimer nebo hexamer, ATPasová aktivita transport váčků od Golgiho aparátu, priming měchýřků SNAPs35-39nejméně tři podtypy (alfa, beta a gama) zapojuje NSF do tzv. docking komplexu PEP protein?kinasa, transfer protein pro inositolfosfáty priming měchýřků CAPS145dimer, interakce s cytoskeletem, afinita k vápníku Ca-dependentní krok primingu synaphin (complexin) ?protein synapsí obřích axonů sépie indukuje vznik SNARE komplexů vyššího řádu po dockingu Proteiny asociované s presynaptickou membránou Cytoplasmatické proteiny
 Vlastnosti/Důležité oblasti molekuly Funkce Syntaxin 35cíl BoTX C1t-SNARE protein, dockin měchýřků, fúze membrán SNAP a 29; 23 kDa predominantní v dospělém NS palmitoylován v oblasti centrální domény fúze membrán, stechiometricky se váže na synaptotagmin v Ca 2+ -závislé fázi výlevu Munc-1865interakce se syntaxinemmolekulární „zarážka bránící předčasnému spojeni v- a t-SNARE, docking měchýřků / může výlev i facilitovat Hrs-2115interakce se SNAP-25docking měchýřků, fúze membrán ( ) RIMasi 200PDZ a C2 domény, motiv zinkových prstů na N- konci docking měchýřků, fúze membrán ( ), regulace Rab3 proteinu ProteinMol. hmotnost (kDa) Vlastnosti/Důležité oblasti molekuly Funkce NSF75homotrimer nebo hexamer, ATPasová aktivita transport váčků od Golgiho aparátu, priming měchýřků SNAPs35-39nejméně tři podtypy (alfa, beta a gama) zapojuje NSF do tzv. docking komplexu PEP protein kinasa, transfer protein pro inositolfosfáty priming měchýřků CAPS145dimer, interakce s cytoskeletem, afinita k vápníku Ca-dependentní krok primingu synaphin (complexin) protein synapsí obřích axonů sépie indukuje vznik SNARE komplexů vyššího řádu po dockingu Proteiny asociované s presynaptickou membránou Cytoplasmatické proteiny.")
30
docking = fyzický kontakt váčku a presynaptické plasmatické membrány je zprostředkován specifickými protein-proteinovými interakcemi mezi v- a t-SNARE proteiny docking váčků je řízená událost, jíž se účastní regulační proteiny bránící předčasnému kontaktu v- a t-SNARE proteinů a vzniku jejich ternárního komplexu důležitou úlohu hrají také GTP-vážící proteiny t-SNARE v dockingu: modulují vznik v/t-SNARE komplexu přes protein Munc-18 (Munc-18) Munc-18 se může vázat syntaxin a jako „zarážka“ (molecular clamp) blokovat kontakt presynaptické membrány s membránou váčku / nebo může naopak fúzi váčku s membránou urychlit v-SNARE v dockingu: rab3, malý GTP-vázající protein, který je v aktivovaném stavu (s navázaným GTP) lokalizován na membráně váčku rab3 je spojen s tzv. RIM proteinem vyskytujícím se na presynaptické membráně rab3 eliminuje působení unc-18 – díky uvolnění GTP a následného odpojení unc-18 od syntaxinu katalyzuje vznik v/t SNARE komplexu Fáze dockingu končí spojením v-SNARE (VAMP/synaptobrevin) a t-SNARE (syntaxin, SNAP-25) proteinů a vznikem stabilního 7S komplexu (docking komplex) První fáze exocytosy - docking
 a t-SNARE (syntaxin, SNAP-25) proteinů a vznikem stabilního 7S komplexu (docking komplex) První fáze exocytosy - docking.")
31
docking komplex: in vitro odolný proti působení SDS a štěpení neurotoxiny je složený 1:1:1 z VAMP/synaptobrevinu, syntaxinu a SNAP-25 bývá také označován jako „minimal fusion machinery“ – minimální fúzní aparát, tedy jádro proteinového komplexu nutného pro neuroexocytosu priming = konverse v aktivní zóně zakotveného měchýřku na měchýřek připravený po patřičném signálu sfúzovat s membránou nervového zakončení poslední na ATP závislý děj v celém procesu kvantového výlevu účastní se jej nejméně dva enzymy hydrolyzující ATP: NSF (N-ethylmalimide- sensitive fusion protein) a fosfatidylinositol-4-fosfát-5-kinasa (PEP protein) Druhá fáze exocytosy - priming
 a fosfatidylinositol-4-fosfát-5-kinasa (PEP protein) Druhá fáze exocytosy - priming")
32
NSF formuje se SNARE proteiny 20 S komplex (fusion complex), v němž po hydrolýze ATP dojde k přeskupení SNARE proteinů tak, aby co nejvhodněji interagovaly se svými protějšky na partnerské membráně. NSF opouští fúzní komplex ještě před vstupem vápníku do nervového zakončení. PEP protein lokálně zvyšuje hladiny fosfatidylinositol-4,5-bisfosfátu (PIns-4,5-P2, PIP2). Kyselé fosfolipidy váže např. synaptotagmin, VAMP a syntaxin mají v oblasti transmembránových domén basické aminokyselinové zbytky, které také mohou s těmito fosfolipidy reagovat. Klíčová pro proces exocytosy je zřejmě reakce různých fosfolipidů (PIP2: PIns-4,5-P2 a PIns-3,4-P2, dále PIns-3,4,5-P3) se synaptotagminem. Complexin: protein kompetující se synaptotagminem o vazbu na SNARE complex antiparalelním helixem se váže na 4 šroubovicovou strukturu SNARE komplexu umožní plné uspořádání SNARE komplexu brání dokonční fúzní rekace, dokud nedojde ke vtoku Ca 2+
. Kyselé fosfolipidy váže např. synaptotagmin, VAMP a syntaxin mají v oblasti transmembránových domén basické aminokyselinové zbytky, které také mohou s těmito fosfolipidy reagovat. Klíčová pro proces exocytosy je zřejmě reakce různých fosfolipidů (PIP2: PIns-4,5-P2 a PIns-3,4-P2, dále PIns-3,4,5-P3) se synaptotagminem. Complexin: protein kompetující se synaptotagminem o vazbu na SNARE complex antiparalelním helixem se váže na 4 šroubovicovou strukturu SNARE komplexu umožní plné uspořádání SNARE komplexu brání dokonční fúzní rekace, dokud nedojde ke vtoku Ca 2+.")
33
9 isoforem, synaptotagmin I se koncentruje v membránách synaptických váčků obsahují jeden transmembránový segment, N-terminální glykosylovanou doménu orientovanou dovnitř měchýřku a cytoplasmatickou C-terminální doménu C-doména tvořena dvěma homologickými motivy: C2A a C2B domény C2A blíže položená k membráně měchýřku; s různě vysokou afinitou 4 ionty Ca 2+ C2B doména váže negativně nabité fosfolipidy (ev. 1 ion Ca 2+ při 3 Ca 2+ na C2A doméně) za klidového stavu C2B doména synaptotagminu váže PIns-3,4,5-P 3, který je po vtoku vápníku do zakončení vytěsněn a C2B doména naváže PIP 2 synaptotagmin dále váže t-SNARE proteiny syntaxin a neurexin a N-typ vápníkových kanálů Synaptotagminy = cca 65 kDa velké integrální membránové proteiny
 za klidového stavu C2B doména synaptotagminu váže PIns-3,4,5-P 3, který je po vtoku vápníku do zakončení vytěsněn a C2B doména naváže PIP 2 synaptotagmin dále váže t-SNARE proteiny syntaxin a neurexin a N-typ vápníkových kanálů Synaptotagminy = cca 65 kDa velké integrální membránové proteiny.")
34
Spuštění exocytosy – Ca 2+ trigerring jedna z klíčových událostí exocytosy u rychlého výlevu nastává exocytosa cca 60 s po influxu Ca 2+ intracelulární vápníkový sensor musí ležet ve velmi těsné blízkosti Ca 2+ kanálu musí vázat ionty Ca 2+ velmi rychle a s relativně nízkou afinitou tímto Ca 2+ sensorem je synaptotagmin po vazbě fosfatidylinositolbisfosfátů na synaptotagmin je váček „přitažen“ k membráně nervového zakončení prudce vzrůstá pravděpodobnost vzniku fúzního póru či hemipóru vznik komplexu synaptotagmin- syntaxin brzdí nízká teplota, inhibice CaM kinasy II a vyvázání extracelulárního Ca 2+ posiluje jej naopak např. stimulace CaM kinasy II doména C2B doména C2A

35
Kompetice complexin–synaptotagmin: (A) Complexin po navázání na SNARE komplex neumožní vazbu synaptotagminu (na jednotlivé proteiny ani na synaptotagmin se neváže, jen na SNARE komplex). (B) Přechodný stav: synaptotagmin rozpozná a váže complexin–SNARE komplex v nepřítomnosti Ca 2+. (C) Výlev: po vtoku Ca 2+ se synaptotagmin váže na Ca 2+ i membrány. Tato konformační změna uvolní complexin a tvarová změna SNARE komplexu přitáhne membrány, do kterých je insertován, k sobě (Martens et al, 2007). Stav s navázaným complexinem bývá ozančována jako metastabilní či superprimovaný -'superprimed' (Tang et al., 2006).
 Přechodný stav: synaptotagmin rozpozná a váže complexin–SNARE komplex v nepřítomnosti Ca 2+. (C) Výlev: po vtoku Ca 2+ se synaptotagmin váže na Ca 2+ i membrány. Tato konformační změna uvolní complexin a tvarová změna SNARE komplexu přitáhne membrány, do kterých je insertován, k sobě (Martens et al, 2007). Stav s navázaným complexinem bývá ozančována jako metastabilní či superprimovaný - superprimed (Tang et al., 2006)..")
36
Poslední fází exocytosy je fúze membrány váčku s membránou terminály a vylití obsahu měchýřku. V současné době jsou nejčastěji uvažovány model kiss-and-run/hemifúzní a fúzní. Fúzní model počítá se vznikem proteinové struktury podobné kanálu, která zprostředkuje vodné spojení mezi vnitřkem váčku a synaptickou štěrbinou. Proteinem SNARE komplexu, tvořícím tento kanál, by mohl být synaptophysin, jehož membránová topologie umožňuje vytvořit strukturu podobnou vodivému spojení (gap junction). Jako fusogen by mohl působit i synaptotagmin, který je schopen přímo reagovat s kyselými fosfolipidy membrán. Zdá se, že SNARE komplexy by také mohly tvořit kolem fúzního póru jakýsi rosetovitý útvar – SNARE super komplex. Mělo by jít o prstenec jednotlivých SNARE komplexů spojených proteinem SNAP- 25. Tento prstenec je pravděpodobně nezbytý pro rychlou, regulovanou exocytosu. Dále by tato SNARE „růžice“ mohla sloužit jako lešení pro vazbu přídatných proteinů s regulační či fusogenní aktivitou. Na základě experimentů prováděných na neuroendokrinních nádorových liniích se zdá, že vylití jednoho váčku je přítomna nejméně trojice SNARE komplexů a že fúzní pór vzniká za účasti mnohočetných kopií transmembránového segmentu syntaxinu. Za nejpravděpodobnější počet SNARE komplexů účastnících se neuroexocytosy je dnes považováno číslo mezi pěti a osmi. Podle hemifúzního modelu jsou dva vodné oddíly (vnitřek měchýřku a extracelulární prostor terminály) odděleny dvojvrstvou lipidů, i když jedna nebo dvě vrstvy jejich membrán už sfúzovaly do stabilního intermediátu. Ke vzniku vodního póru je třeba jen prasknutí sfúzovaných membrán. Úloha SNARE komplexu je v tomto modelu pasivní: napomáhá vhodné orientaci membrán váčku a terminály a „přitlačí“ je co nejblíže k sobě. Energie uvolněná při vzniku stabilní ternární struktury SNARE komplexu je použita k překonání energetické bariéry a přímo přispívá k fúzi membrán. Hemifúze je pravděpodobněji jen přechodnou fází výlevu, který je plně fúzní.
. Jako fusogen by mohl působit i synaptotagmin, který je schopen přímo reagovat s kyselými fosfolipidy membrán. Zdá se, že SNARE komplexy by také mohly tvořit kolem fúzního póru jakýsi rosetovitý útvar – SNARE super komplex. Mělo by jít o prstenec jednotlivých SNARE komplexů spojených proteinem SNAP- 25. Tento prstenec je pravděpodobně nezbytý pro rychlou, regulovanou exocytosu. Dále by tato SNARE „růžice mohla sloužit jako lešení pro vazbu přídatných proteinů s regulační či fusogenní aktivitou. Na základě experimentů prováděných na neuroendokrinních nádorových liniích se zdá, že vylití jednoho váčku je přítomna nejméně trojice SNARE komplexů a že fúzní pór vzniká za účasti mnohočetných kopií transmembránového segmentu syntaxinu. Za nejpravděpodobnější počet SNARE komplexů účastnících se neuroexocytosy je dnes považováno číslo mezi pěti a osmi. Podle hemifúzního modelu jsou dva vodné oddíly (vnitřek měchýřku a extracelulární prostor terminály) odděleny dvojvrstvou lipidů, i když jedna nebo dvě vrstvy jejich membrán už sfúzovaly do stabilního intermediátu. Ke vzniku vodního póru je třeba jen prasknutí sfúzovaných membrán. Úloha SNARE komplexu je v tomto modelu pasivní: napomáhá vhodné orientaci membrán váčku a terminály a „přitlačí je co nejblíže k sobě. Energie uvolněná při vzniku stabilní ternární struktury SNARE komplexu je použita k překonání energetické bariéry a přímo přispívá k fúzi membrán. Hemifúze je pravděpodobněji jen přechodnou fází výlevu, který je plně fúzní..")
37
Kiss-and-run hypotéza: 1955: Katz, del Castillo 1973: název pochází od Bruna Ceccarellilo váček je stále zakotven v aktivní zóně a přechodně se otevírá Extended kiss-and-run hypotéza: váček se po vylití asi 40% obsahu opět uzavře a je připraven na další výlev pór není uzavřen kompletně, před vlastním výlevem i po něm protékají z váčku neuropřenašeče post-spike proud je nižší o snížený koncentrační gradient přes délku póru (z váčku vytekla část neuropřenašeče) Lisa Mellander a kol., Sci Reports 2:907, 2012
 Lisa Mellander a kol., Sci Reports 2:907, 2012")
38
Kononenko, N.L., Pechstein, A., Haucke, V. (2013) The tortoise and the hare revisited. eLife, 2, e01233. Modely exocytózy: (B) Utrarychlý mechanismus recyklace váčků. Po výlevu se vytvoří endocytotické jamka bez účasti klathrinu. (C) Endocytóza založená na klathrinu. Váček je plně absorbován do membrány, a to velmi pomalu. Jak model B, tak C vyžadují k odpojení nově vzniklých endocytotických váčků od membrány zakončení účast GTPasy dynaminu. (A)"kiss-and-run" model – synaptické váčky se po vzniku přechodného póru a uvolnění obsahu uzavřou, během cca 1 s jsou pak v místě výlevu recyklovány, aniž by by byly absorbovány do membrány Proč ta variabilita? Různé frekvence aktivity neuronů, mezidruhová variabilita…
 Utrarychlý mechanismus recyklace váčků. Po výlevu se vytvoří endocytotické jamka bez účasti klathrinu. (C) Endocytóza založená na klathrinu. Váček je plně absorbován do membrány, a to velmi pomalu. Jak model B, tak C vyžadují k odpojení nově vzniklých endocytotických váčků od membrány zakončení účast GTPasy dynaminu. (A) kiss-and-run model – synaptické váčky se po vzniku přechodného póru a uvolnění obsahu uzavřou, během cca 1 s jsou pak v místě výlevu recyklovány, aniž by by byly absorbovány do membrány Proč ta variabilita. Různé frekvence aktivity neuronů, mezidruhová variabilita….")
39
BoNT/B-Syt-II komplex při 2.6 Å rozlišení: už před lety byl navržen "double receptor" model popisující současnou vazbu BoNT na gangliosidy nervových terminál a současně na proteinové receptory ToxinJeho neuronální cíl botulinotoxin ASNAP-25 botulinotoxin Bsynaptobrevin botulinotoxin C1syntaxin botulinotoxin Dsynaptobrevin botulinotoxin ESNAP-25 botulinotoxin Fsynaptobrevin botulinotoxin Gsynaptobrevin tetanotoxinsynaptobrevin -latratoxin neurexin; CIRL/latrophilin Proteiny, zejména toxiny = eventuální proteasy štěpící specifické proteinové cíle a tak narušující proces exocytosy neurotoxiny Clostridia botulinum navozují díky paralýze hlavových a periferních nervů asymetrickou descendentní paralýzu tetanotoxin Clostridia tetanum působí na motorických neuronech i jejich neuronech inhibičních a vyvolává křeče pavoučí jed („černá vdova“) -latrotoxin vyvolává hlubokou paralýzu způsobenou masivním uvolněním a následnou deplecí acetylcholinu na nervosvalovém spojení
 -latrotoxin vyvolává hlubokou paralýzu způsobenou masivním uvolněním a následnou deplecí acetylcholinu na nervosvalovém spojení")
41
My se zastavíme u: cholinergních drah dopaminergních drah serotoninergních drah gabaergních drah glutamátergních drah noradrengních drah histaminergních drah Různé neuropřenašeče mají v mozku své poměrně dobře popsané neuropřenašečové systémy.

42
Cholinergní neurony se nacházejí v jádrech roztroušených v celém mozku a jejich axony inervují většinu oblastí CNS. Þ hlavní cholinergní vstupy do kortexu a hippokampu jsou z jader na spodině předního mozku, zejména z jader septa a ncl. basalis Þ pro cholinergní neurony těchto jader jsou typické rozvětvené difúzní projekce inervující kůru mozkovou, hippokampus, amygdalu, thalamus a mozkový kmen. Þ cholinergní systém v CNS se obecně účastní procesů učení, kognice a vzniku paměťových stop Þ látky blokující muskarinové ACh receptory (atropin nebo skopolamin) nabourávají proces získávání a vykonávání neučeného chování, stejně jako léze ncl. basalis Þ naopak farmaka blokující acetylcholinesterasu (enzym degradující ACh), např. fysostygmin, zlepšují výkon v paměťových a učebních úlohách a mohou částečně zmírnit následky lézí některých oblastí předního mozku Þ jsou to především modulačními vstupními cestami ke kortikálním a hippokampálním neuronům
 nabourávají proces získávání a vykonávání neučeného chování, stejně jako léze ncl. basalis Þ naopak farmaka blokující acetylcholinesterasu (enzym degradující ACh), např. fysostygmin, zlepšují výkon v paměťových a učebních úlohách a mohou částečně zmírnit následky lézí některých oblastí předního mozku Þ jsou to především modulačními vstupními cestami ke kortikálním a hippokampálním neuronům.")
43
v CNS je acetylcholin uvolňován na synapsích tvořených kolaterálami spinálních motoneuronů, které končí na Renshawových buňkách tato rychlá excitační nikotinická synapse je nicméně pro CNS poměrně atypická v CNS je cholinergní odpověď zprostředkovaná spíše ACh receptory muskarinovými to se projevuje mj. vzrůstem propustnosti membrán neuronů pro kationy, vzrůstem či poklesem různých typů draslíkových vodivostí a poklesem propustnosti membrány pro vápník nikotinické ACh receptory jsou v CNS také exprimovány, ale jejich synaptické funkce jsou hůře popsány účinky ACh, obzvláště na kortikální úrovni, jsou velmi komplexní a specifické pro danou oblast mozku interakce s dalšími neuromodulačními systémy: ACh může na pyramidové buňky kůry mozkové působit buď excitačně, nebo inhibičně excitace je navozena působením ACh na M1 receptory na vlastních pyramidových buňkách, inhibici způsobí navázání ACh na M2 receptory GABAergních interneuronů

44
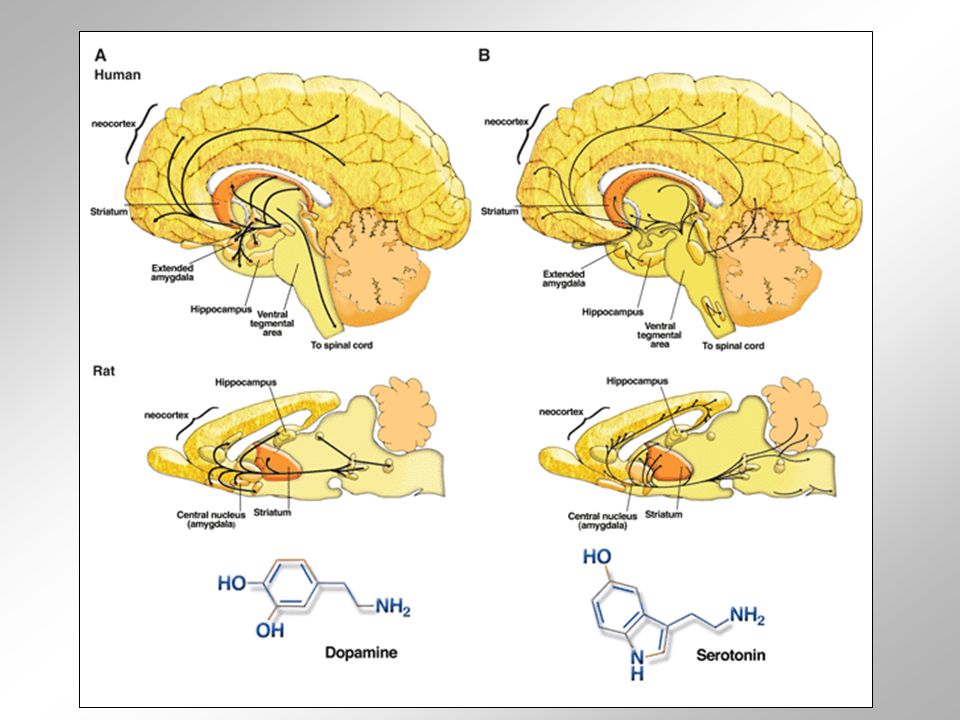
Dopamin = jeden ze zástupců biogenních aminů v CNS v mozkovém kmeni se nalézají čtyři hlavní dopaminergní jádra jedno z nich se nachází v ncl. arcuatus a vybíhají z něj axony inervující hypothalamus zbylá tři jádra leží ve středním mozku a projikují primárně do bazálních ganglií relativně malé množství dopaminergních neuronů bohatě ramifikuje a inervuje značné oblasti mozku v mozku potkana např. každý z cca 7000 dopaminergních neuronů ležících v substantia nigra (jedno z dopaminergních jader středního mozku), zásobuje zhruba 250 000 varikozitami své cíle v bazálních gangliích progresivní degenerace dopaminergních neuronů (zejména v pars compacta substatiae nigrae) vede ke ztrátě dopaminergní inervace neuronů bazálních ganglií a charakteristickým motorickým projevům spojeným s Parkinsonovou ch.
, zásobuje zhruba varikozitami své cíle v bazálních gangliích progresivní degenerace dopaminergních neuronů (zejména v pars compacta substatiae nigrae) vede ke ztrátě dopaminergní inervace neuronů bazálních ganglií a charakteristickým motorickým projevům spojeným s Parkinsonovou ch..")
45
Serotonin = další biogenní amin CNS serotoninergní neurony lokalizovány jen do několika málo jader mozkového kmene jde o dvě rostrální a dvě kaudální nuclei raphe, která leží ve středové ose mozkového kmene mezi středním mozkem a prodlouženou míchou (termín „raphe“ pochází z francouzského názvu pro šev, spojnici) jádra uložená v prodloužené míše projikují do míchy hřbetní a ovlivňují míšní dráhy zapojené do přenosu bolestivých signálů, stejně jako aktivitu míšních interneuronů a motoneuronů jádra uložená ve středním mozku a Varolově mostě inervují téměř celý mozek společně s projekcemi z locus coeruleus tvoří část vzestupného retikulárního aktivačního systému.
 jádra uložená v prodloužené míše projikují do míchy hřbetní a ovlivňují míšní dráhy zapojené do přenosu bolestivých signálů, stejně jako aktivitu míšních interneuronů a motoneuronů jádra uložená ve středním mozku a Varolově mostě inervují téměř celý mozek společně s projekcemi z locus coeruleus tvoří část vzestupného retikulárního aktivačního systému.")
47
Kyselina -aminomáselná (GABA) = dominantní inhibiční aminokyselinou CNS obratlovců její koncentrace je vyšší než koncentrace ACh a noradrenalinu zhruba 25-45% synapsí v CNS obratlovců je GABAergních v CNS byly identifikovány tři třídy receptorů pro GABA: ionotropní GABA A Rs ionotropní GABA C Rs a metabotropní GABA B receptory mnoho podjednotkových kombinací receptoru, jejichž distribuce, farmakologie a projevy se v CNS lokálně liší (velká diverzita podjednotek) GABAergní neurony (interneurony i projikující neurony) jsou umístěny mj. v mozkovém kmeni a předním mozku v retikulární formaci mozkového kmene jsou relativně malé GABAergní neurony zastoupeny spolu s většími GABAergními neurony projikujícími a zřejmě lokálně inhibují glutamátergní neurony ascendentní retikulární aktivační formace (ARAS). Rozdílná distribuce mRNA různých GABA A podjednotek v mozku potkana. Autoradiogramy (A) 1, 3 (B)a 6 (C) podjednotky GABA A receptorů. (B)(C) (A)
. Rozdílná distribuce mRNA různých GABA A podjednotek v mozku potkana. Autoradiogramy (A) 1, 3 (B)a 6 (C) podjednotky GABA A receptorů. (B)(C) (A).")
48
Glutamát = hlavním excitační neuropřenašeč CNS působí na NMDA a non-NMDA (zejména AMPA) receptory distribuce těchto receptorů zahrnuje oblasti kortexu i rozsáhlé oblasti subkortikální, přičemž zastoupení NMDA a non-NMDA receptorů se v různých oblastech liší mnoho velkých glutamátergních neuronů se nachází v retikulární formaci mozkového kmene a zřejmě slouží jako primární neuropřenašeč ARAS glutamátergní neurony také projikují do thalamu a kortexu v kortexu je glutamát uvolňován zejména ve stavu spontánní bdělosti. Distribuce glutamátových receptorů v mozku potkana. (A), distribuce NMDA receptorů označených tritiovaným glutamátem. (B), rozložení AMPA receptorů. Distribuce obou typů receptorů se široce překrývá, zejména v kortexu, nicméně lze pozorovat i oblasti jen s určitým typem receptoru (např. nízký výskyt NMDA receptorů oproti AMPA receptorům v molekulární vrstvě mozečku). C, kortex; G, granulární vrstva mozečku; M, molekulární vrstva mozečku. Z Nichols a ost., 2001. (B)(A)
, distribuce NMDA receptorů označených tritiovaným glutamátem. (B), rozložení AMPA receptorů. Distribuce obou typů receptorů se široce překrývá, zejména v kortexu, nicméně lze pozorovat i oblasti jen s určitým typem receptoru (např. nízký výskyt NMDA receptorů oproti AMPA receptorům v molekulární vrstvě mozečku). C, kortex; G, granulární vrstva mozečku; M, molekulární vrstva mozečku. Z Nichols a ost., (B)(A).")
49
Noraderenergní neurony = málo zmůže mnoho ;) v CNS sdruženy do locus coeruleus ve Varolově mostu u spodiny čtvrté komory mozkové v CNS potkana obsahuje každý locus coeruleus (párově na obou stranách mozkového kmene) zhruba 1500 buněk: těchto dohromady 3000 neuronů představuje přibližně polovinu všech noradrenergních buněk mozku potkana toto poměrně malé množství buněk vysílá rozsáhlé projekce do mozečku, kůry mozkové, thalamu, hippokampu a hypothalamu. stimulace locus coeruleus nebo aplikace noradrenalinu vyvolává různou odpověď v závislosti na typu zúčastněného receptoru v pyramidových buňkách hippokampu blokuje noradrenalin cestou -adrenergních receptorů vznik následné hyperpolarizace po salvě akčních potenciálů, čímž dramaticky zvyšuje množství akčních potenciálů vznikajících při prodloužené depolarizaci membrány projekce locus coeruleus tvoří část vzestupného retikulárního aktivačního systému (ARAS) tato projekční dráha reguluje pozornost, probouzení a cirkadiální rytmy.
 tato projekční dráha reguluje pozornost, probouzení a cirkadiální rytmy..")
50
Histamin byl objeven roku 1920 v tkáni jater a plic na periferii jsou hlavními místy uskladnění a výlevu mastocyty Histamin ovlivňuje různé periferní tkáně a účastní se mnoha fyziologických procesů, jakými jsou např. alergické reakce, odpověď tkáně na poranění nebo žaludeční sekrece přes své receptory spřažené s G-proteiny slouží histamin jako neuropřenašeč i v mozku cestou H 1 receptorů histamin depolarizuje cholinergní neurony ncl. basalis snížením propustnosti jejich membrány pro draslík a aktivací na tetrodotoxin necitlivých sodíkových kanálů v buňkách ganglion nodosum blokuje přes H 1 receptory histamin draslíkové kanály generující následnou hyperpolarizci po akčním potenciálu, což také vede ke zvýšení jejich excitability. těla histaminergních neuronů jsou uložena v ncl. tuberomamillaris hypothalamu a jejich axony s bohatými kolaterálami inervují podobně jako u ostatních biogenních aminů téměř všechny části CNS. Jejich synapse jsou difúzní a jen příležitostně vytvářejí klasické pre- a postsynaptické uspořádání. Histaminergní neurony inervují také glie a malé cévy a kapiláry. zdá se, že regulují obecné aktivity mozku (úroveň stavu bdělosti či energetický metabolismus.

51
Co si pamatovat z dnešní přednášky axon, dendrity, dendritické trny chemická synapse příklady neuropřenašečů plnění váčku neuropřenašeči, priming, docking, exocytosa neuropřenašečové systémy mozku

Podobné prezentace







>")










 vylučované kůrou nadledvinek (aldosteron, kortisol); 2) vylučované pohlavními žlázami (progesteron, testosteron, estradiol)>")

