Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Eduard Kejnovský + Roman Hobza
EVOLUČNÍ GENOMIKA III. EVOLUCE GENOMŮ

2
OBSAH 1. Velikost genomu 2. Extrémní genomy
3. Změny ve velikosti genomu 4. Nekódující DNA a velikost genomu 5 .Topografie genomu 6. Genomové projekty

3
Definice genomu - celková genetická informace organizmu
u autonomních forem života vždy v DNA prokaryota: cirkulární chromosom + plazmidy eukaryota: chromosomy v jádře, mitochondrie a chloroplasty

4
První genomy – vznik fúzováním malých kružnic DNA
(Trifonov 1995) první genomy: lineární nebo cirkulární? malé kružnicové DNA genomy, disperzní genom fúzování, geny jako autonomní DNA, počty kopií statisticky stejné – podobné přenosům plazmidů Fáze: pregenomická rekombinační genomická
 první genomy: lineární nebo cirkulární malé kružnicové DNA genomy, disperzní genom. fúzování, geny jako autonomní DNA, počty kopií statisticky stejné – podobné přenosům plazmidů. Fáze: pregenomická. rekombinační. genomická.")
5
První genomy – vznik fúzováním malých kružnic DNA
(Trifonov 1995) Důkazy: periodicita délek proteinů (123AK u prokaryot, 152 AK u eukaryot) nejsnazší cirkularizace bp periodicita výskytu Met pozůstatkem extrachromosomální DNA – 10% genomu mobilní elementy, lyzogenní fágy, genomy organel, horizontální přenos replikace genů a satelitů prostřednictvím eccDNA
 Důkazy: periodicita délek proteinů (123AK u prokaryot, 152 AK u eukaryot) nejsnazší cirkularizace bp. periodicita výskytu Met. pozůstatkem extrachromosomální DNA – 10% genomu. mobilní elementy, lyzogenní fágy, genomy organel, horizontální přenos. replikace genů a satelitů prostřednictvím eccDNA.")
6
VELIKOST GENOMU

7
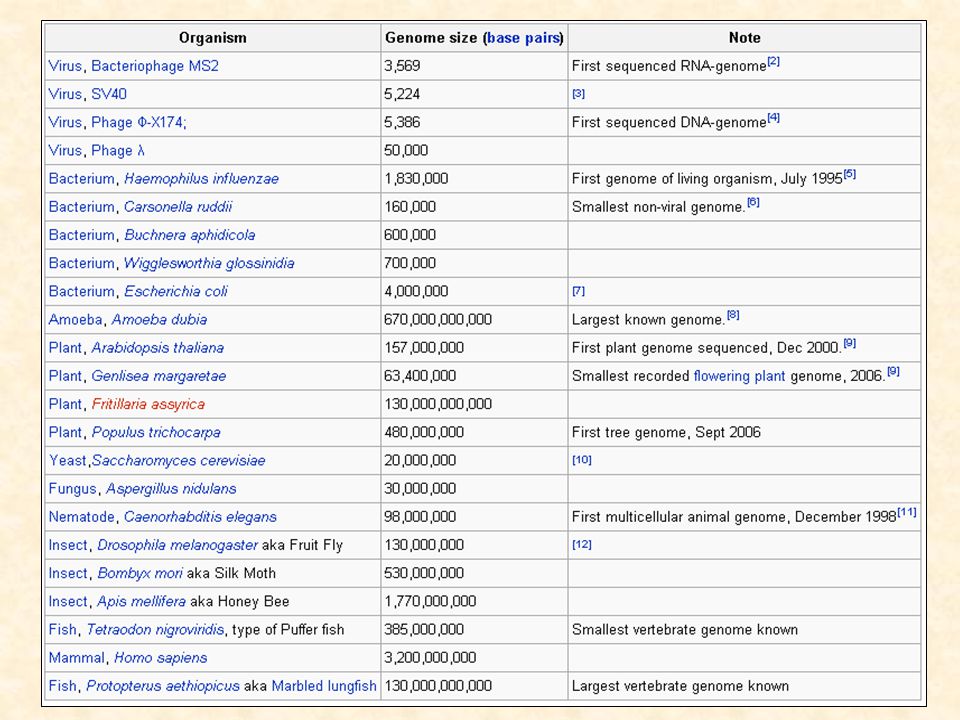
Velikosti genomů 106 107 108 109 1010 1011 savci ptáci plazi
ptáci plazi obojživelníci ryby kostnaté ryby chrupavčité ostnokožci korýši hmyz měkkýši červi plísně rostliny řasy houby grampozitivní bakterie gramnegativní bakterie mykoplazmata 106 107 108 109 1010 1011

8
Drosophila melanogaster
Paradox hodnoty C Definice: velikost genomu organizmu není v korelaci s komplexitou organizmu (H. sapiens 200x menší genom než Amoeba dubia) obsah kódující DNA podobný, příčinou je nekódující DNA, sobecká DNA 180 Mb 18,000 Mb Drosophila melanogaster Podisma pedestris Skutečnost, že genomy eukaryot se liší až 80,000-krát - snad největší výzva pro genomové holisty
 obsah kódující DNA podobný, příčinou je nekódující DNA, sobecká DNA. 180 Mb. 18,000 Mb. Drosophila melanogaster. Podisma pedestris. Skutečnost, že genomy eukaryot se liší až 80,000-krát - snad největší výzva pro genomové holisty.")
9
Klíč k řešení paradoxu hodnoty C se nachází v negenových oblastech
podíl genů podíl TE genová frakce (%) velikost genomu velikost genomu
 velikost genomu. velikost genomu.")
10
Hlavní komponenty eukaryotického genomu
Kódující části genů: - u prokaryot tvoří většinu genomu - u eukaryot méně, člověk genů – 1.5% Introny: původně považovány za příčinu C-paradoxu, tvoří většinu genů Pseudogeny: klasické, retropseudogeny, 19000 člověk, myš, 51 kur, 33 kvasinka, 176 drosophila Mobilní elementy: - LTR, nonLTR – SINE, LINE, DNA transposony- MITE Numt, Nupt: - inzerce promiskuitní DNA

11
Velikosti genomů a zbytečná DNA
„The majority of the genome consisted of now-extinct genes“ - Susumu Ohno “Triumphs as well as failures of nature's past experiments appear to be contained in our genome” - Susumu Ohno „The evolution of the large-scale features of the genome is one of the most difficult, perharps the most difficult, question in evolutionary biology“ - Maynard Smith

12
Velikosti genomů prokaryot
Bakteriální genom: 6105 ->107 (=0.6-10Mb) nejmenší známý: Mycoplasma genitalium (480 genů kódujících proteiny, 3 rRNA geny, 37 tRNA genů) Procesy ovlivňující velikost bakteriálního genomu: Genová duplikace, malé delece a inzerce, transpozice, horizontální přenos, ztráta genů v parazitických liniích, atd. Velikost genomů prokaryot je zhruba úměrná počtu genů
 nejmenší známý: Mycoplasma genitalium (480 genů kódujících proteiny, 3 rRNA geny, 37 tRNA genů) Procesy ovlivňující velikost bakteriálního genomu: Genová duplikace, malé delece a inzerce, transpozice, horizontální přenos, ztráta genů v parazitických liniích, atd. Velikost genomů prokaryot je zhruba úměrná počtu genů.")
13
Velikosti genomů prokaryot

14
EXTRÉMNÍ GENOMY

15
Minimální velikost genomu
„One day a scientist will drop gene number 297 into a test tube, then number 298, then 299… and presto: what was not alive a moment ago will be alive now. The creature will be as simple as life can be. But it will still be life. And humans will have made it, in an ordinary glass tube, from off-the-shelf chemicals…“ - Hayden, 1999: Co je život? Strategie: 1. odstranění či inaktivace genů 2. syntéza minimálního genomu

16
“Minimální” genom – Mycoplasma
580 kb genom/ 480 genů pro proteiny/ 37 genů pro tRNA 2209 inzercí transpozonů/ ve 140 genech 1354 míst, kde inzerce nebyla letální genů nepostradatelných (glykolýza) genů postradatelných 100 genů má neznámou funkci!!! různý vliv inzercí podle polohy inzerce v genu
 genů postradatelných. 100 genů má neznámou funkci!!! různý vliv inzercí podle polohy inzerce v genu.")
17
“Minimální” genom: Bacillus subtilis život s 271 geny
192 genů zcela nepostradatelných na bohatém prostředí: zpracování informace, buněčná stěna, energie 79 genů podstatných Jen 4% genů má neznámou funkci

18
Minimální velikost genomu: (a) Analytický přístup
(Mushegian a Koonin, 1996): Srovnání kompletních bakteriálních genomů: E. coli H.influenzae M.genitalium 1,146 1,129 889 18 10 239 1 Translation Replication Transcription Recombination and DNA repair Chaperone-like proteins Anaerobic metabolism Lipid or cofactor biosynthesis Transmembrane transporter Some unknown function [Překrývající se ortologní geny (239)] + [přesun neortologních genů] – [geny specifické pro parazitické bakterie nebo funkční redundance] = 256 genes
: Srovnání kompletních bakteriálních genomů: E. coli. H.influenzae. M.genitalium. 1,146. 1, Translation. Replication. Transcription. Recombination and DNA repair. Chaperone-like proteins. Anaerobic metabolism. Lipid or cofactor biosynthesis. Transmembrane transporter. Some unknown function. [Překrývající se ortologní geny (239)] + [přesun neortologních genů] – [geny specifické pro parazitické bakterie nebo funkční redundance] = 256 genes.")
19
Minimální velikost genomu: (b) Experimentální přístup
(Itaya, 1995): Knock-out 79 náhodně vybraných genů Bacillus subtilis: pouze 6 letálních, 73 je postradatelných 7.5% (6/79) genomu je nepostradatelných - B.subtilis genome: 4.2106bp 7.5% = 3.2105bp Průměrná velikost genu je 1.25kb, takže minimální velikost genomu 254 genů
: Knock-out 79 náhodně vybraných genů Bacillus subtilis: pouze 6 letálních, 73 je postradatelných. 7.5% (6/79) genomu je nepostradatelných. - B.subtilis genome: 4.2106bp 7.5% = 3.2105bp. Průměrná velikost genu je 1.25kb, takže minimální velikost genomu 254 genů.")
20
Největší prokaryotické genomy
Pseudomonas aeruginosa (bakterie): genů přes 6 Mb přirozeně rezistentní k antibiotikům (ochranný obal) R-faktor, žije ve společenství jiných bakterií, konjugace lidský patogen (kožní n., močové, dýchací a trávicí cesty) Nostoc punctiforme (sinice): ORF - 8.9 Mb - repetice, transpozony - fotoautotrofní, také fakultativně heterotrofní - možnost symbiózy s rostlinami i houbami Repetice N.p.
: genů. přes 6 Mb. přirozeně rezistentní k antibiotikům (ochranný obal) R-faktor, žije ve společenství jiných bakterií, konjugace. lidský patogen (kožní n., močové, dýchací a trávicí cesty) Nostoc punctiforme (sinice): ORF Mb. - repetice, transpozony. - fotoautotrofní, také fakultativně heterotrofní. - možnost symbiózy s rostlinami i houbami. Repetice N.p.")
21
Velikosti genomů eukaryot
Eukaryotický genom: 106~ 6.91011 (8.8Mb – 670Gb) Nejmenší známý: Saccharomyces cerevisiae Největší známý: Amoeba dubia Velikosti eukaryotických genomů NEjsou úměrné počtu genů nebo komplexitě organizmu
 Nejmenší známý: Saccharomyces cerevisiae. Největší známý: Amoeba dubia. Velikosti eukaryotických genomů NEjsou úměrné počtu genů nebo komplexitě organizmu.")
23
ZMĚNY VE VELIKOSTI GENOMU

24
Zvětšení genomu celkové zvětšení: polyploidizace (duplikace celého genomu) duplikace části genomu, zmnožení počtu chromosomů duplikace genů a skupin genů amplifikace transpozonů inzerce virové DNA inzerce organelové DNA expanze (mikro)satelitů Vzrůstající komplexita živých forem byla doprovázena vzrůstem velikosti genomů a počtu genů
satelitů. Vzrůstající komplexita živých forem byla doprovázena vzrůstem velikosti genomů a počtu genů.")
25
Evoluční síly ovlivňující velikost genomu

26
Změny ve velikosti genomů: plynulé nebo skoky?
Plynulost: - savci, ptáci, kostnaté ryby postupná akumulace nebo delece, malé segmenty DNA normální distribuce velikostí genomů v těchto skupinách podíl jedinečných sekvencí konstantní, mění se spolu se změnou velikosti genomu, je dán velikostí intronů Skoky (kvantové změny): bezobratlí, rostliny (50% nahosemenných je polyploidní), vyjímečně obratlovci polyplodizace, přidatná replikace některého chromosomu, kryptopolyploidizace (koordinovaná replikace všech repeticí v genomu – Chironomus) kvantové rozdíly mezi blízkými příbuznými, velké rozdíly ve velikostech genomů
: bezobratlí, rostliny (50% nahosemenných je polyploidní), vyjímečně obratlovci. polyplodizace, přidatná replikace některého chromosomu, kryptopolyploidizace (koordinovaná replikace všech repeticí v genomu – Chironomus) kvantové rozdíly mezi blízkými příbuznými, velké rozdíly ve velikostech genomů.")
27
Růst velikosti genomu: Polyplodizace
Fúze diploidní a haploidní gamety vede k triplodnímu jádru, triploidní organizmus je však sterilní Chyba v meioze vede k diploidním gametám

28
Celkové zvětšení genomu
Polyploidizace (duplikace celého genomu): Autopolyploidie: multiplikace jedné základní sady chromosomů, jeden organizmus, chyba při meioze (růže…) Allopolyploidie: kombinace geneticky odlišných sad chromosomů (pšenice…) Kryptopolyploidie: dávná polyplodizace, organizmus se jeví jako diploidní, přestavby – delece, translokace…, kvasinka, obratlovci, obilniny, odhalení až sekvenací Duplikace části genomu: chromosomů - polysomie (trisomie), většinou letální, genová imbalance, evolučně irelevantní genů nebo skupin genů
: Autopolyploidie: multiplikace jedné základní sady chromosomů, jeden organizmus, chyba při meioze (růže…) Allopolyploidie: kombinace geneticky odlišných sad chromosomů (pšenice…) Kryptopolyploidie: dávná polyplodizace, organizmus se jeví jako diploidní, přestavby – delece, translokace…, kvasinka, obratlovci, obilniny, odhalení až sekvenací. Duplikace části genomu: chromosomů - polysomie (trisomie), většinou letální, genová imbalance, evolučně irelevantní. genů nebo skupin genů.")
29
Výhody a nevýhody polyploidů
Skoková (kvantová) změna velikosti genomu – nadějné monstrum musí najít partnera Schopnost obsazovat nová prostředí, převládá v evoluci rostlin, i u obratlovců Noví polyploidi – nestabilní – radikalní přestavby genů – rychlá diverzifikace - speciace (změněný počet chromosomů vede k reprodukční izolaci) Multiplikace klíčových genů v regulaci vývoje – multiplikace klastrů homeotických genů vedla k enormní diverzitě druhů ryb
 změna velikosti genomu – nadějné monstrum musí najít partnera. Schopnost obsazovat nová prostředí, převládá v evoluci rostlin, i u obratlovců. Noví polyploidi – nestabilní – radikalní přestavby genů – rychlá diverzifikace - speciace (změněný počet chromosomů vede k reprodukční izolaci) Multiplikace klíčových genů v regulaci vývoje – multiplikace klastrů homeotických genů vedla k enormní diverzitě druhů ryb.")
30
Polyploidní plodiny - tetraploidi: kukuřice, bavlna, brambor, zelí
- hexaploidi: pšenice, chrysantéma - oktoploidi: jahodník

31
Genom kvasinky: Polyploidizace nebo duplikace segmentů?
54 nepřekrývajících se duplikovaných oblastí 50 z nich si zachovalo stejnou orientaci vzhledem k centroméře nebyla nalezena žádná triplikovaná oblast, přičemž 7 oblastí by bylo očekáváno dle Poissonova rozložení příbuzné druhy bez duplikace – Kluveromyces waltii a Ashbya gossypii 457 genových párů – 17% se vyvíjí rychleji než jejich homology u K. waltii Duplikace jsou výsledkem tetraploidie spíše než postupných regionálních duplikací

32
Polyploidizace v linii obratlovců
S. Ohno: - dvě genomové duplikace u obratlovců – hypotéza „2R“ (2 rounds) - některé genové rodiny mají 4 členy u obratlovců a jednu u bezobratlých - srovnání ježovky (bezobratlý) s obratlovci (člověkem, myší a rybou Fugu) - v genomu člověka dlouhé segmenty ve čtyřech kopiích podél všech chromosomů Duplikace genomu před 450 mil let napomohla vzrůstu komplexity a diverzifikaci obratlovců
 - některé genové rodiny mají 4 členy u obratlovců a jednu u bezobratlých. - srovnání ježovky (bezobratlý) s obratlovci (člověkem, myší a rybou Fugu) - v genomu člověka dlouhé segmenty ve čtyřech kopiích podél všech chromosomů. Duplikace genomu před 450 mil let napomohla vzrůstu komplexity a diverzifikaci obratlovců.")
33
Polyploidizace u rostlin
Arabidopsis: - duplikace před 200 a 80 mil. let možná i více než 4 genomové duplikace asi 60% genomu leží v duplikovaných segmentech 50% genů v nich je konzervováno

34
Počet genů a evoluce duplikovaného genomu
ABCDEFGHIJKLM a b c d e f g h i j k l m NOPQRSTUVWXYZ n o p q r s t u v w x y z 26 genes, 2 chomosomes 26 genes, 2 chomosomes Tetraploidizace ABCDEFGHIJKLM a b c d e f g h i j k l m NOPQRSTUVWXYZ 52 genes, 4 chomosomes Ztráta genů n o p q r s t u v w x y z AB DEF HI KLM AB DEF H s t v w x y b c e g h j k m Translokace b c e g h j k m N PQ TUV X Z N PQ TUV X Z n o p r s t v w x y n o p r I KLM Crossing over bc E’F H s t v w x y AB D e’ g h j k m N PQ TUV X Z 36 genes, 4 chomosomes n o p r I KLM

35
Genomová obezita u rostlin – jednosměrný proces?
Teorie sobecké DNA (Dawkins, 1976) jediným cílem DNA je se replikovat Dva modely: pouze růst genomů růst i zmenšování genomů Limity růstu genomů: fitness hostitele mechanizmy potlačující amplifikaci – metylace mechanizmy odstraňování repeticí – rekombinace - BARE – 42x více solo LTR než vnitřních částí - rekombinace uvnitř i mezi elementy - selekce na kratší LTR, solo LTR nejdelší
 jediným cílem DNA je se replikovat. Dva modely: pouze růst genomů. růst i zmenšování genomů. Limity růstu genomů: fitness hostitele. mechanizmy potlačující amplifikaci – metylace. mechanizmy odstraňování repeticí – rekombinace. - BARE – 42x více solo LTR než vnitřních částí. - rekombinace uvnitř i mezi elementy. - selekce na kratší LTR, solo LTR nejdelší.")
36
Vzrůst počtu genů, potlačení šumu a biologická komplexita
Prokaryota/eukaryota: Potlačení šumu - separace transkripce a translace, jaderná membrána a histony Obratlovci: Potlačení šumu – metylace genů, mobilních elementů a duplikátů obratlovci eukaryota prokaryota

37
Komplexita vzniká snadno, kontroluje se ale obtížně
Nekódující DNA: Regulační systém na bázi RNA? Komplexita vzniká snadno, kontroluje se ale obtížně introny MicroRNA

38
Regulační systém na bázi RNA
Vznik nového regulačního systému

39
Mechanizmy zvětšení genomů
Duplikativní (retro)transpozice Nerovnoměrný (unequal) crossing-over Replikační klouzání (Replication slippage) Genová amplifikace (rolling circle replication)
transpozice. Nerovnoměrný (unequal) crossing-over. Replikační klouzání (Replication slippage) Genová amplifikace (rolling circle replication)")
40
NEKÓDUJÍCÍ DNA a VELIKOST GENOMU

41
Teorie úlohy nekódující DNA: adaptivní role vs. sobecká DNA
Jaké evoluční síly produkují „zbytečnou“DNA? Jaká je role „zbytečné“DNA? Proč selekce toleruje „zbytečnou“ DNA? Adaptivní role: vliv nadbytečné DNA na fenotyp, vliv na velikost jádra a buňky, ochrana kódujících sekvencí před mutacemi, pufrování koncentrace regulačních proteinů Sobecká DNA (junk DNA): Parazitické sekvence, mobilní elementy, fixace genetickým driftem, velikost genomu je tolerovatelné maximum závisející na ekologických a vývojových potřebách organizmu
: Parazitické sekvence, mobilní elementy, fixace genetickým driftem, velikost genomu je tolerovatelné maximum závisející na ekologických a vývojových potřebách organizmu.")
42
Big Bang v genomu kukuřice
polyploidizace amplifikace transpozonů a retroelementů Exploze před 6 mil let Více inzercí do mezigenových oblastí Exponenciální růst mezigenových vzdáleností

43
Závislost velikosti genomu na zaměpisné šířce a nadmořské výšce
- korelace mezi velikostí genomu a teplotním režimem větší genomy nebo polyploidi: - arktické lososovité ryby - zooplankton arktických jezer (Daphnia, Bosmina) - rostliny v polárních oblastech - populace v teplých oblastech jsou diploidní
 - rostliny v polárních oblastech. - populace v teplých oblastech jsou diploidní.")
44
Obsah DNA je proměnlivý i v rámci jedince
Zvýšení obsahu DNA - endopolyploidie a polytenie: drosophila - polytenní chromosomy ve slinných žlázách Daphnia - tkáňově-specifické rozdíly v ploidii, C, vliv na morfologii hlavy indukovanou predátorem Bombyx mori – ploidní buňky žláz produkujících hedvábí korelace ploidie a velikosti buňky Snížení obsahu DNA v somatických buňkách: nematoda, bičíkovci, dvoukřídlí, např. Cyclops strenuus – somatické b. mají 5% obsahu DNA oproti oplozenému vajíčku vysvětlení z hlediska sobecké DNA – delece DNA ze slepé somatické linie a ne ze zárodečné linie, jak repetitivní DNA může tak významně ovlivnit genetické mechanizmy svého hostitele změna obsahu DNA během života jako odpověď na stimuly – rozpor s Darwinismem

45
- Vliv velikosti genomu na fenotyp + Velikost genomu koreluje s:
velikostí jádra velikostí buňky (nucleotypic effect) dobou mitózy a meiózy - minimální generační dobou velikostí semen odpovědí letniček vůči CO2 dobou vývoje embrya u mloků - rychlostí bazálního metabolismu u obratlovců (negativní korelace) (malý genom ptáků a netopýrů - rychlý metabolismus při letu, velký genom ryb - estivace za hypoxických podmínek) morfologickou komplexitou mozků u žab a mloků (negativní korelace)
 dobou mitózy a meiózy. - minimální generační dobou. velikostí semen. odpovědí letniček vůči CO2. dobou vývoje embrya u mloků. - rychlostí bazálního metabolismu u obratlovců (negativní korelace) (malý genom ptáků a netopýrů - rychlý metabolismus při letu, velký genom ryb - estivace za hypoxických podmínek) morfologickou komplexitou mozků u žab a mloků (negativní korelace)")
46
TOPOGRAFIE GENOMU

47
Uspořádání genů v genomech

48
Přestavby pořadí genů u bakterií
(Watanabe et al. 1997)
")
49
Syntenie - konzervativita pořadí genů
druhy III. I. II. A A B B C1 C C2 D konzervativita genů rozdílné mezigenové sekvence E E

50
Makro- vs. mikro- kolinearita
- konzervativita v dlouhých úsecích mikrostruktura více dynamická, mix s jinými geny polyploidie urychlí evoluci mikrostruktury změny již u prvních generací syntetického alotetraploida u Brassica Dávná divergence: A. thaliana vs. tomato: divergence před 150mil. let Nedávná divergence: A. thaliana vs. Capsella rubella: divergence před mil. let (Bancroft, 2001)
")
51
Rychlá evoluce oblasti rezistence k patogenu
(Bancroft, 2001) Columbia Lansberg erecta gen „RPP5“ – rezistence k Perenospora parasitica rozdíly mezi A.t. ekotypy Lansberg erecta a Columbia většinou pseudogeny tandemové duplikace a nerovnoměrný crossing-over je mechanizmem rychlé evoluce
 Columbia. Lansberg erecta. gen „RPP5 – rezistence k Perenospora parasitica. rozdíly mezi A.t. ekotypy Lansberg erecta a Columbia. většinou pseudogeny. tandemové duplikace a nerovnoměrný crossing-over je mechanizmem rychlé evoluce.")
52
Počty chromosomů u různých druhů
Extrémisté: mravenec Myrmecia pilosula – 1 pár, samec 1 chromosom kapradina Ophioglossum reticulatum – 630 párů

53
Různé počty chromosomů u blízce příbuzných druhů jelínků Muntiacus
různé druhy

54
Recyklace pohlavních chromosomů u drozofily
První Y chromosom: „original Y“: - vymizel před více než 60 mil. lety Druhý Y chromosom: „ancestral Y“: - párování B chromosomu s X chromosomem ~ 60 mil years ago - získání úžitečných genů z autosomů - degenerace Třetí Y chromosom: „neo Y“ - fúze X s autosomem - připojení ancestrálního Y na A - opět degenerace neo-Y

55
B chromosomy - parazité
Definice: přidatné postradatelné chromosomy přítomné v některých jedincích některých populací určitých druhů, které zřejmě vznikly z A chromosomů Vlastnosti B chromosomů: 1. postradatelnost 2. původ z A chromosomů, odlišnost od A chromosomů 3. nerekombinují 4. neúčastní se meiózy, nemendelovská dědičnost Historie: Wilson (1906) - „extra“ chromosomy u hmyzu Metapodius Stevens (1908) - u brouka Diabrotica Kuwada (1915) – kukuřice Rozšíření: 10 druhů hub, 1300 rostlin, 500 živočichů Převaha u určitých taxonů – Compositae, Graminae, Liliaceae, Orthoptera, např. u hmyzu Orthoptera ~ u 10-15% druhů
 - „extra chromosomy u hmyzu Metapodius. Stevens (1908) - u brouka Diabrotica. Kuwada (1915) – kukuřice. Rozšíření: 10 druhů hub, 1300 rostlin, 500 živočichů. Převaha u určitých taxonů – Compositae, Graminae, Liliaceae, Orthoptera, např. u hmyzu Orthoptera ~ u 10-15% druhů.")
56
B chromosomy - parazité
Vznik: aneuploidie, ze sex chromosomů (i naopak), z jiných druhů Počet: - vyjímečně více než 3-4 B chromosomy, rozdíly mezi populacemi i mezi jedinci - extrémy: 50 u Pachyphytum fittkaui, 34 u kukuřice, 26 u Fritillaria japonica, 24 u myši Apodemus peninsulae - mitoticky stabilní – stejný počet v buňkách téhož jedince Velikost a struktura: - velikost průměrných chromosomů, extrémy – největší (ryba Astyanax scabripinnis) nebo nejmenší (myš Reithrodontomys megalotis) - metacentrické nebo akrocentrické, heterochromatické, - obsahují repetice, satelitní DNA, rDNA, transpozony (mikrodisekce a PCR), nerekombinují Chování v meióze: - nepárujíc se s partnerem, univaletní, segregují přednostně k určitému pólu, podobnost s pohlavními chromosomy, MSCI,
, z jiných druhů. Počet: - vyjímečně více než 3-4 B chromosomy, rozdíly mezi populacemi i mezi jedinci. - extrémy: 50 u Pachyphytum fittkaui, 34 u kukuřice, 26 u Fritillaria japonica, 24 u myši Apodemus peninsulae. - mitoticky stabilní – stejný počet v buňkách téhož jedince. Velikost a struktura: - velikost průměrných chromosomů, extrémy – největší (ryba Astyanax scabripinnis) nebo nejmenší (myš Reithrodontomys megalotis) - metacentrické nebo akrocentrické, heterochromatické, - obsahují repetice, satelitní DNA, rDNA, transpozony (mikrodisekce a PCR), nerekombinují. Chování v meióze: - nepárujíc se s partnerem, univaletní, segregují přednostně k určitému pólu, podobnost s pohlavními chromosomy, MSCI,")
57
Srovnání lidských a myších chromosomů
člověk myš Od evoluční divergence došlo k četným přestavbám, které umístily bloky genů do různých kombinací.

58
Inverze v genomech člověka a šimpanze
Člověk: červená-žlutá-zelená Šimpanz a gorila: červená-zelená-žlutá velikost inverzí

59
Chromosomové přestavby
zlom a znovuspojení rekombinace mezi repeticemi (Introduction to Genetic Analysis. 7th ed) Griffiths, Anthony J.F.; Gelbart, William M.; Miller, Jeffrey H.; Lewontin, Richard C.
 Griffiths, Anthony J.F.; Gelbart, William M.; Miller, Jeffrey H.; Lewontin, Richard C.")
60
Izochorový model organizace genomu
Isochory: bloky genomové DNA (stovky kb-Mb) s charakteristickým GC-obsahem, ostrý přechod - fragmentace genomové DNA na fragmenty kb separace fragmentů podle obsahu bází shlukování do skupin s diskrétními GC-obsahy, u člověka typy L1, L2, H1, H2 a H3 (GC nejbohatší, tvoří jen 3% ale obsahuje 25% genů) Mozaiková struktura genomu obratovců a rostlin klasický model Původ izochor: 1. Výsledek mutací? 2. Výsledek selekce? 3. Genová konverze? lidská MHC oblast
 s charakteristickým GC-obsahem, ostrý přechod. - fragmentace genomové DNA na fragmenty kb. separace fragmentů podle obsahu bází. shlukování do skupin s diskrétními GC-obsahy, u člověka typy L1, L2, H1, H2 a H3 (GC nejbohatší, tvoří jen 3% ale obsahuje 25% genů) Mozaiková struktura genomu obratovců a rostlin. klasický model. Původ izochor: 1. Výsledek mutací 2. Výsledek selekce 3. Genová konverze lidská MHC oblast.")
61
GENOMOVÉ PROJEKTY

62
První genomové projekty - začátek éry genomiky
1977: první kompletní genom (virus) - bakteriofág phiX174 (5 386 bp) Sanger et al., Nature 265, 1995: první kompletní genom autonomního organizmu - bakterie Haemophilus infuenzae (1.38 Mb) Fleischmann et al., Science 269, 1996: první eukaryotický organizmus - kvasinka Saccharomyces cerevisiae Goffeau et al., Science 274,
 - bakteriofág phiX174 (5 386 bp) Sanger et al., Nature 265, : první kompletní genom autonomního organizmu - bakterie Haemophilus infuenzae (1.38 Mb) Fleischmann et al., Science 269, : první eukaryotický organizmus - kvasinka Saccharomyces cerevisiae. Goffeau et al., Science 274,")
63
Genomové projekty 1996: Saccharomyces cerevisiae (12 Mb/ 6548 genes)
1998: Caenorhabditis elegans (97 Mb/ genů) 2000: Drosophila melanogaster (137 Mb/ genů) Mus musculus (3 300 Mb) 2000: Arabidopsis thaliana (125 Mb/ genů)
 2000: Drosophila melanogaster. (137 Mb/ genů) Mus musculus. (3 300 Mb) 2000: Arabidopsis thaliana. (125 Mb/ genů)")
64
Jak zjistit stav genomových projektů

65
Objem dat v databázích roste exponenciálně
> bp > sekvencí > struktur ~ organizmů

66
Hlavní databáze Genbank - sekvence DNA, veřejně přístupná
SWISS-PROT - sekvence proteinů, popis funkce, struktury domén dbEST - sekvence cDNA z různých organizmů REBASE - restriktázy a metylázy PEDANT - kompletní nebo parciální sekvence genomů PDB - 3D koordináty makromolekulárních struktur REPBASE – repetitivní sekvence

67
Genomové sekvenační projekty duben 2008
762 ukončených projektů: 53archea 626 bakterie 83 eukaryota 975 probíhajících projektů: 1749 prokaryota 935 eukaryota 89 archea Ukončené eukaryotické genomy: protozoa: Plasmodium falciparum (23 Mb) fungi: Saccharomyces cerevisiae (12 Mb) Neurospora crassa (43 Mb) nematodes: Caenorhabditis elegans (97 Mb) insects: Drosophila melanogaster (137 Mb) Anopheles gambiae ( 278 Mb) fish: Ciona intestinalis (117 Mb) mammals: Homo sapiens (2900 Mb) Mus musculus (3300 Mb) Rattus norvegicus (2750 Mb) plants: Arabidopsis thaliana (115 Mb) Oryza sativa (420 Mb)
 fungi: Saccharomyces cerevisiae (12 Mb) Neurospora crassa (43 Mb) nematodes: Caenorhabditis elegans (97 Mb) insects: Drosophila melanogaster (137 Mb) Anopheles gambiae ( 278 Mb) fish: Ciona intestinalis (117 Mb) mammals: Homo sapiens (2900 Mb) Mus musculus (3300 Mb) Rattus norvegicus (2750 Mb) plants: Arabidopsis thaliana (115 Mb) Oryza sativa (420 Mb)")
68
Genomové sekvenační projekty
Kompletní/nekompletní Kompletní genomy Bakterie/eukaryota/archea Sekvenační centra

69
Sekvenování lidského genomu

Podobné prezentace