Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Onkogenetika, genetické mechanismy vzniku nádorů.
Pavel Vodička 2014 Ústav experimentální medicíny v.v.i AV ČR 1. LF UK Praha

2
VÝCHODISKA Sporadické formy rakoviny jsou charakterizovány polygenní součinností v komplexní interakci s vlivy prostředí (mikro- i makro-) a životního stylu Hodnocení rizika CRC v souvislosti s polymorfismy a haplotypy řady genů s cílem identifikovat relevantní kandidátní geny. Alternativní přístup založený na identifikaci relevantních variant vnímavosti pomocí GWAS. Verifikace a výklad GWAS dat (post-GWAS fine-mapping) cestou meta-analýz kandidátních genů a „next generation“ sekvenováním. Definování jak genetických, tak fenotypických rysů CRC (např. DNA reparační funkční testy)
 a životního stylu. Hodnocení rizika CRC v souvislosti s polymorfismy a haplotypy řady genů s cílem identifikovat relevantní kandidátní geny. Alternativní přístup založený na identifikaci relevantních variant vnímavosti pomocí GWAS. Verifikace a výklad GWAS dat (post-GWAS fine-mapping) cestou meta-analýz kandidátních genů a „next generation sekvenováním. Definování jak genetických, tak fenotypických rysů CRC (např. DNA reparační funkční testy)")
3
Procento variance Nádorová onemocnění ve Švédsku,Dánsku a Finsku
Genetic Shared environmental Nonshared environmental % 100 80 60 40 20 Lung Ovary Stomach Pancreas Prostate Bladder Colorectum Cervix uteri Leukemia Corpus uteri Breast (female) Lichtenstein et al, 2000, NEJM
 Lichtenstein et al, 2000, NEJM.")
4
Funkce genů může být ovlivněna:
genovými variantami (polymorfismy) alteracemi v počtu kopií (amplifikace, delece, duplikace, změny v počtu chromozomů) změny v struktuře genu, chromosomální struktura (translokace, inverse atd.) genové mutace - substituce, delece, inserce v kódujícich sekvencích , na hranicích exonů a intronů epigenetické modifikace (imprinting, DNA metylatce a modifikace histonů - acetylace/deacetylace histonů, metylace nebo fosforylace)
 alteracemi v počtu kopií (amplifikace, delece, duplikace, změny v počtu chromozomů) změny v struktuře genu, chromosomální struktura (translokace, inverse atd.) genové mutace - substituce, delece, inserce v kódujícich sekvencích , na hranicích exonů a intronů. epigenetické modifikace (imprinting, DNA metylatce a modifikace histonů - acetylace/deacetylace histonů, metylace nebo fosforylace)")
5
Předpoklady Xenobiotika životního prostředí vykazují genotoxický účinek na člověka, hlavně díky přímé vazbě na DNA. Předpokládá se, že genotoxický účinek (poškození DNA a chromosomů) je v souvislosti se vznikem sporadických forem rakoviny . Genetický základ může formovat/ovlivňovat vnímavost jednotlivce k maligním onemocněním a riziku rakoviny. Aparát DNA opravy zachovává integritu genomu tím, že opravuje poškození DNA indukované expozicí potenciálním karcinogenům. Vztah mezi genotypem genů DNA opravy a fenotypem je zásadní pro identifikaci kritických časných markerů karcinogeneze Proč incidence rakoviny mezi silně exponovanými jedinci není dramaticky vyšší než je incidence běžné populace? Pavel Vodička,
 je v souvislosti se vznikem sporadických forem rakoviny . Genetický základ může formovat/ovlivňovat vnímavost jednotlivce k maligním onemocněním a riziku rakoviny. Aparát DNA opravy zachovává integritu genomu tím, že opravuje poškození DNA indukované expozicí potenciálním karcinogenům. Vztah mezi genotypem genů DNA opravy a fenotypem je zásadní pro identifikaci kritických časných markerů karcinogeneze. Proč incidence rakoviny mezi silně exponovanými jedinci není dramaticky vyšší než je incidence běžné populace Pavel Vodička,")
6
Kaskáda dějů v genotoxicitě a karcinogeneze
XENOBIOTICS Metabolic activation or deactivation REACTIVE METABOLITES INDIVIDUAL SUSCEPTIBILITY Biotransformation and DNA repair genes DNA ADDUCTS PROTEIN ADDUCTS CYTOTOXICITY, APOPTOSIS DNA REPAIR DNA SSB Transition lesions, CA, SCE, deletions, chromosomal instability MUTAGENESIS HPRT, tumor suppressor genes, oncogenes Additional factors CARCINOGENESIS Pavel Vodicka,

7
Mutageny Fyzikální: radiace
UV (ultrafialové záření) → T-T, C-C, T-C dimery = chyby v replikaci a transkripci ionizing (rtg, γ) přímý účinek→ DNA zlomy nepřímý účinek → ionizace molekul → DNA zlomy Chemické– alkylační činidla - addukty - analoga bazí – chyby v párování bazí - acridinové barviva – inserce - kyselina dusičná –deaminace bazí – chyby v párování bazí přímé mutageny nepřímé mutageny– po metabolické aktivaci (cytochrom dependent oxygenázy) vznikají reaktivní produkty Biologické–viry - virové nukleové kyseliny se integrují do genomu hostitelské buňky
 → T-T, C-C, T-C dimery = chyby v replikaci a transkripci. ionizing (rtg, γ) přímý účinek→ DNA zlomy. nepřímý účinek → ionizace molekul → DNA zlomy. Chemické– alkylační činidla - addukty. - analoga bazí – chyby v párování bazí. - acridinové barviva – inserce. - kyselina dusičná –deaminace bazí – chyby v párování bazí. přímé mutageny. nepřímé mutageny– po metabolické aktivaci (cytochrom dependent oxygenázy) vznikají reaktivní produkty. Biologické–viry - virové nukleové kyseliny se integrují do genomu hostitelské buňky.")
8
METABOLIZMUS fenantrenu

9
Metabolizmus styrénu CYP2E1*5A, *6, *1B, *1D CYP2E1 styrene oxide (SO)
EPHX1 EPHX1 Tyr113His EPHX1 His139Arg GSTM1 GSTT1 GSTP1 GSTs styrene glycol (SG) ADH ADH2 *1/*2 ADH3 *1/*2 mandelic acid (MA) PHEMA 1 PHEMA 2 Metabolizmus styrénu phenylglyoxylic acid (PGA)
 ADH. ADH2 *1/*2. ADH3 *1/*2. mandelic acid. (MA) PHEMA 1. PHEMA 2. Metabolizmus styrénu. phenylglyoxylic acid (PGA)")
10
Biomarkers most frequently applied in our Department:
Transient biomarkers of carcinogenesis Biomarkers of genomic landscape of cancers Phenotypic biomarker-function of substantial biological systems Biomarkers of epigenetic regulations in carcinogenesis Biomarkers of cancer phenotype Biomarkers of treatment response (genetic, epigenetic, target vs. surrogate) Main requirements for biomarkers Sensitivity Validity Reproducibility Availability Informativness Cost effectiveness Complexity vs Interpretability
 Main requirements for biomarkers. Sensitivity. Validity. Reproducibility. Availability. Informativness. Cost effectiveness. Complexity vs Interpretability.")
11
Potřeba validních biomarkerů v časné
Využití biomarkerů : Zlepšení hodnocení expozice Příspěvek ke zpřesnění křivky dávka/účinek při nízkých hladinách expozice. Příspěvek k objasnění mechanismů genotoxicity a karcinogenity Identikace vnímavých jedinců a stanovení stupně vnímavosti Stanovení patogeneze a/nebo prognózy Potřeba validních biomarkerů v časné karcinogenezi

12
Biomarkery používané v in vivo studiích
Comet test – jednořetězcové zlomy DNA Specifické DNA addukty – primární poškození DNA Micronucleus = chromosomální fragment v cytoplazmě erytrocytů nebo binukleárních buněk po zablokování cytokineze cytochalasinem Cytogenetické analýzy buněk kostní dřeně u experimentálních zvířat nebo lidských periferních lymfocytů – chromozomální zlomy nebo přestavba (rearangement) Výměna sesterských chromatid – inkorporace BUdR do DNA během kultivace buněk (pro dva buněčné cykly)– různá substituce obou chromatid = různé zbarvení sesterských chromatid Individuální vnímavost
 Výměna sesterských chromatid – inkorporace BUdR do DNA během kultivace buněk (pro dva buněčné cykly)– různá substituce obou chromatid = různé zbarvení sesterských chromatid. Individuální vnímavost.")
13
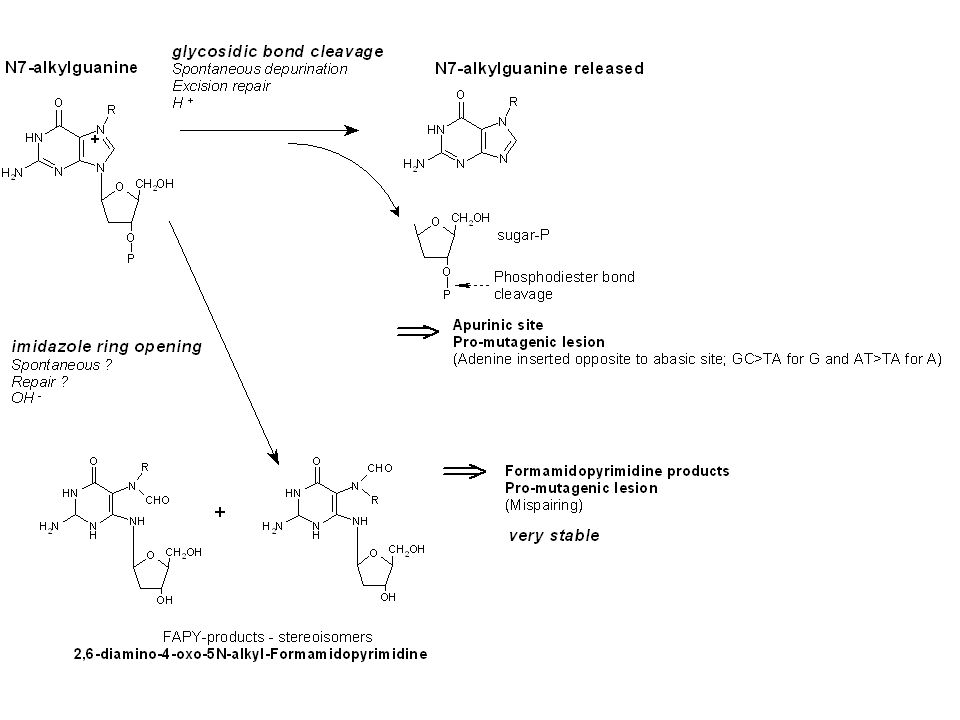
Hlavní místa elektrofilních ataků v nucleosidech

14
A B Spektrum specifických DNA adduktů styrénu (A) a 1,3 butadienu (B)
bN7-gua 45% αN7-gua 29% aN6-ade 6% aN3-ade 6% bN3-ade 3% bN1-ade 2% bN3-ura 2% aN1-hypox 2% aN4-cyt 1% bN6-ade 1% aN2-gua 3% N3-ura 1% N3-cyt 1% N6-ade 1% N3-ade 10% N7-gua 70% N1-ade 17% From: Vodicka et al. Mutat Res 2002; Vodicka et al. Drug Metab Rev 2006; Koskinen and Plna Chem Biol Interact 2002 Pavel Vodicka,

16
Deaminací vzniká hypoxantín a
AT-GC tranzice

17
Mutace = permanentní děditelná změna genetického materiálu = změna v sekvenci nucleotidů nebo přestavba DNA v genomu Mutace: spontánní indukované Mutace: somatické germline či gametické

18
Mutace: genomové mutace – změny v počtu chromozomů: a) euploidní změna = znásobení haploidních chromosomálních setů (triploidy, tetraploidy) b) aneuploidy = další chromozom (trisomie) nebo chybějící chromozom (monosomie) chromozomové mutace= strukturální chromozomální aberace– zlomy a výměny chromozomálních segmentů genové mutace = kvalitativní nebo kvantitativní změny v sekvencích DNA
 aneuploidy = další chromozom (trisomie) nebo chybějící chromozom (monosomie) chromozomové mutace= strukturální chromozomální aberace– zlomy a výměny chromozomálních segmentů. genové mutace = kvalitativní nebo kvantitativní změny v sekvencích DNA.")
19
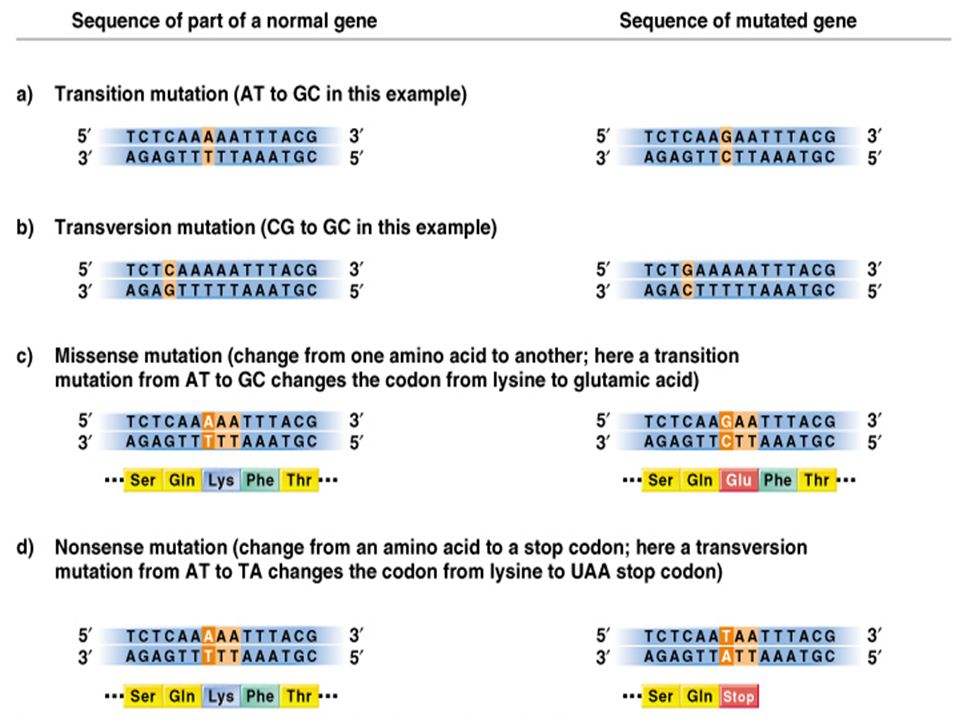
GENOVÉ mutace mutace bez jakékoliv změny aminových kyselin (degenerace genetického kódu) „ MISSENSE mutation“ - záměna jedné aminokyseliny za jinou „ NONSENSE mutation“- mutace způsobí jeden ze tří „stop“ kodonů „ ELONGATION mutations“ změna stop kodónu na triplet kódujíci aminovou kyselinu „FRAME SHIFT mutations“- inserce, delece Mutace v rRNA a tRNA genech - chyby v translaci

20
Found (dominating BS for SO) Vodicka et al. Drug Metab Rev 2006
Nejvýznamnější DNA addukty a jejich předpokládaná role v mutagenezi. Adduct Mutation N7-guanine GCTA N2-guanine GC TA O6-guanine GC AT 8-OH-guanine GC TA 8-OH-adenine AT CG N3-adenine AT TA N1-adenine AT GC N6-adenine AT GC N3-cytosine CG AT, GC TA - AT GC - GC TA, AT TA Expected Found Found (dominating BS for SO) Vodicka et al. Drug Metab Rev 2006 Pavel Vodicka,
 Vodicka et al. Drug Metab Rev Pavel Vodicka,")
21
Schematic representation of Go repair in mammalian cels

24
Předpokládané důsledky různých DNA adduktů
O6-ALKYL- TRANSFERASE REPAIR REPAIR EXCISION REPAIR REPAIR REPAIR REPAIR REPAIR SSB PERSISTANCE ?? MUTATIONS, CYTOTOXICITY MISPAIRING, GCTA, ATTA MUTATIONS ATGC MUTATIONS, CYTOTOXICITY MUTATIONS, CHROMOSOMAL ABERRATIONS, CYTOTOXICITY MUTATIONS, CYTOTOXICITY GCTA MUTATIONS, CYTOTOXICITY MISPAIRING, MUTATIONS, BLOCK OF REPLICATIONS, CYTOTOXICITY CELL DEATH TUMOUR DEVELOPMENT

25
Hlavní cesty DNA opravy
DNA oprava je komplexní systém obrany (cca. 120 genů) s cílem udržet genomovou integritu a je z hlavních cest předcházejích karcinogenezu. Interindividuální rozdíly v kapacitě DNA oprav-významný faktor rizika maligních onemocnění (včetně CRC). CRC-komplexní onemocnění, kde vnímavost může být ovlivněna genetickými variantami v DNA reparačních systémech.
 s cílem udržet genomovou integritu a je z hlavních cest předcházejích karcinogenezu. Interindividuální rozdíly v kapacitě DNA oprav-významný faktor rizika maligních onemocnění (včetně CRC). CRC-komplexní onemocnění, kde vnímavost může být ovlivněna genetickými variantami v DNA reparačních systémech.")
26
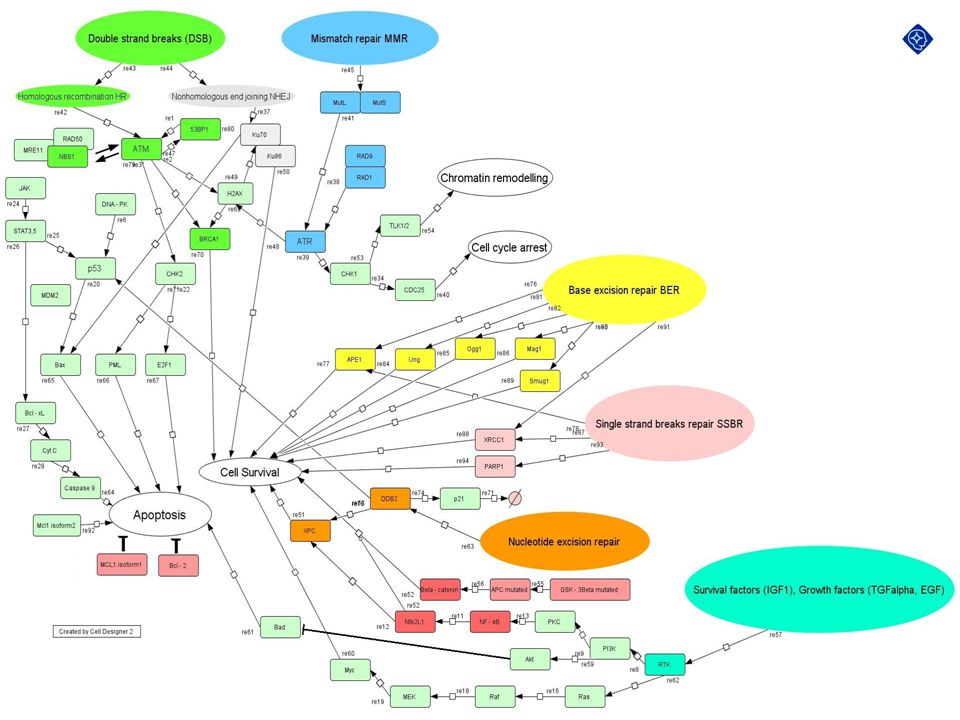
Repair Pathways

27
Klíčová role opravy DNA v nádorové patogenezi
Růstové factory receptors, signal transducers EGFR, IGF-1, MAPK hormony insulin, estrogen onkogeny cytokiny a molekuly zánětu TGF1 / Smad3, interleukins Cell cycle DNA damage sensing epigenetické faktory DNA repair stárnutí Chronický zánět a stres mikroprostředí excesívní exo/endo poškození DNA obezita

29
Telomerová dysfunkce/udržování „DNA damage response mechanisms“
processing DNA damage sensing DNA repair DNA damage signaling Double-strand break repair Telomere maintenance Cell cycle checkpoint control Control of apoptosis Martínez and Blasco, Aging Cell Oct;9(5):
:")
30
Bázově-excizní oprava
damaged base removed by glycosylase glycosylase apurinic site removed by specific endonuclease AP endonuclease gap filled by DNA polymerase and sealed with ligase polymerase, ligase Obr. : R.Štětina

31
Nucleotidová excizní oprava
endonuclease Incision near to the damaged site by endonuclease exonuclease The piece with the damaged base is removed by exonuclease polymerase DNA polymerase fils the gap ligase The process is completed by sealing of strands by the ligase Obr. : R.Štětina

32
Opravy chybného párování bází řízené metylací
Opravy chybného párování bází řízené metylací (mismatch repair, MMR) hrají klíčovou roli v zachování genomické stability. MMR opravuje chyby v replikaci DNA, zahrnující nesprávně zařazané nukleotidy, inzerce a delece. A dále chyby v průběhu mitotické nebo meiotické rekombinace. Složky systému MMR jsou také zahrnuty v odpovědi na poškození DNA, buněčném cyklu, apoptóze a remodelaci chromatinu. Geny MMR jsou důležité jak v etiologii familiárního CRC, tak v souboru sporadicky se vyskytujících nádorů. Dysfunkce MMR vede k nárůstu nestability v mikrosatelitních sekvencích (MSI) napříč genomem.
 hrají klíčovou roli v zachování genomické stability. MMR opravuje chyby v replikaci DNA, zahrnující nesprávně zařazané nukleotidy, inzerce a delece. A dále chyby v průběhu mitotické nebo meiotické rekombinace. Složky systému MMR jsou také zahrnuty v odpovědi na poškození DNA, buněčném cyklu, apoptóze a remodelaci chromatinu. Geny MMR jsou důležité jak v etiologii familiárního CRC, tak v souboru sporadicky se vyskytujících nádorů. Dysfunkce MMR vede k nárůstu nestability v mikrosatelitních sekvencích (MSI) napříč genomem.")
33
Původ dvouřetězcových zlomů (DSB)::
endogenní: oxidativní metabolizmus topoizomerázy (jednořetězcové zlomy-SSB,DSB) chyby v DNA replikaci nebo opravě DNA rekombinace –“ crossing over“ v meiose V(D)J rekombinace, „class switching“ immunoglobulinových genů exogenní: radiace (ionizujíci, ultrafialové), chemikálie, restrikční endonukleázy DSB jsou indukovány přímo – ionizujícím zářením nebo nepřímo–UV zářením, chemikáliemi + enzymatic ká oprava→ SSB(single-strand breaks) → DSB (double-strand breaks)
 chyby v DNA replikaci nebo opravě. DNA rekombinace – crossing over v meiose. V(D)J rekombinace, „class switching immunoglobulinových genů. exogenní: radiace (ionizujíci, ultrafialové), chemikálie, restrikční endonukleázy. DSB jsou indukovány přímo – ionizujícím zářením. nebo nepřímo–UV zářením, chemikáliemi + enzymatic ká oprava→ SSB(single-strand breaks) → DSB (double-strand breaks)")
34
Opravy dvouřetězcových zlomů:
HR = homologní rekombinace - potřeba sesterské chromatidy (v G2, S fázích buněčného cyklu) - či přítomnost homologního chromosomu (meiotic recombination) NHEJ = nonhomologous end joining – hlavně vn G0, G1 - bez přítomnosti homologního templátu – „error prone“ Oba typy: - eliminace DSB - or mutation and chromosomal aberration – consequence of erroneous repair (HR, NHEJ) Řada genů zodpovědných za HR: e.g. BRCA1, BRCA2 XRCC1,XRCC2,NBS1,Rad geny etc.
 - či přítomnost homologního chromosomu (meiotic recombination) NHEJ = nonhomologous end joining – hlavně vn G0, G1. - bez přítomnosti homologního templátu – „error prone Oba typy: - eliminace DSB. - or mutation and chromosomal aberration – consequence of erroneous repair (HR, NHEJ) Řada genů zodpovědných za HR: e.g. BRCA1, BRCA2. XRCC1,XRCC2,NBS1,Rad geny etc.")
35
PŘÍKLADY Z GENETICKÉ TOXICOLOGIE A MOLEKULÁRNÍ EPIDEMIOLOGIE
Molekulárně epidemiologické studie definují biomarkery, jež charakterizují interní expozici, biologicky účinnou dávku xenobotik jakož i biologický účinek (často prediktivní pro vznik rakoviny) a variabilitu v individuální vnímavosti.
 a variabilitu v individuální vnímavosti.")
36
O6 addukty v granulocytech a lymfocytech exponovaných a kontrolních osob
Intraindividual variability: F=0.28, P=0.872

37
Kombinace genotypů GSTM1 + GSTT1
1-THB-AdeninOVe DNA addukty ve vztahu k GSTM1 a GSTT1 genotypům u osob exponovaných 1,3-BD GSTT1 GSTM1 Kombinace genotypů GSTM1 + GSTT1 Pavel Vodicka,

38
Comparison of Animal and Human DNA and Hemoglobin Adducts after Styrene, 1,3-Butadiene and Propylene oxide exposures Styrene 1,3-Butadiene Propylene oxide Mouse Human Rat Exposure (mg/m3) 750 131a/115b 660c 3 1250d ≈10 a1-alkyl-adenine 1 2.3 11 4.5 200 0.7 bAmino-terminal valine (pmol/g) - 3.0 2.7 Ratio in Styrene 1,3-Butadiene Propylene oxide Hemoglobin adducts in humans 1.0 9.1 10.5 1-alkyl-adenine adducts in animals 17.5 40.0 1-alkyl-adenine adducts in humans 85.7 4.0 a,b – exposures corresponding to Hemoglobin and N1-Ade adducts determinations, c – inhalation exposure for 5 days, d – inhalation exposure for 20 days; Values recalculated to make 1-Ade adducts formation comparable Pavel Vodicka,
 a/115b. 660c d. ≈10. a1-alkyl-adenine bAmino-terminal valine (pmol/g) Ratio in. Styrene. 1,3-Butadiene. Propylene oxide. Hemoglobin adducts in humans alkyl-adenine adducts in animals alkyl-adenine adducts in humans a,b – exposures corresponding to Hemoglobin and N1-Ade adducts determinations, c – inhalation exposure for 5 days, d – inhalation exposure for 20 days; Values recalculated to make 1-Ade adducts formation comparable. Pavel Vodicka,")
39
DNA adduct proportion: a lesson from animal experiments
Styrene inhalation in NMRI mice: 40-fold excess of 7-alkylG as compared to 1-alkylA in lungs 1-alkylA 5-fold lower in liver than in lungs, whereas 7-alkylG undetected (!)-efficient repair in liver? Several-fold higher mEH expression in liver? Only persisting 7-alkylG in lungs in comparison to 7-alkylG excreted in urine (spontaneous depurination, BER) accounts for 0.5% of the total 7-G alkylation. Relevance for malignant transformation? The total 7-G alkylation of styrene, calculated as a percentage of the total styrene uptake, accounts for %. Relevance for malignant transformation? Assumption to organ-specific DNA repair Naturally occuring mutational rate ranges from 1-10/109 dNp. Vodicka et al. Toxicol Appl Pharmacol, 2006 Pavel Vodicka Pavel Vodicka
-efficient repair in liver Several-fold higher mEH expression in liver Only persisting 7-alkylG in lungs in comparison to 7-alkylG excreted in urine (spontaneous depurination, BER) accounts for 0.5% of the total 7-G alkylation. Relevance for malignant transformation The total 7-G alkylation of styrene, calculated as a percentage of the total styrene uptake, accounts for %. Relevance for malignant transformation Assumption to organ-specific DNA repair. Naturally occuring mutational rate ranges from 1-10/109 dNp. Vodicka et al. Toxicol Appl Pharmacol, Pavel Vodicka. Pavel Vodicka.")
40
Různé kombinace genotypů v souvislosti s poškozením DNA
A= EPHX1 (high activity), GSTM1 (positive) vs. B= EPHX1 (low activity), GSTM1 (null) P=0.002 B A A= EPHX1 (high activity), GSTT1 (positive) vs. B= EPHX1 (low activity), GSTT1 (null) P=0.005 B A
, GSTM1 (positive) vs. B= EPHX1 (low activity), GSTM1 (null) P= B. A. A= EPHX1 (high activity), GSTT1 (positive) vs. B= EPHX1 (low activity), GSTT1 (null) P= B. A.")
41
XPD (AA), EPHX1 (low activity) XPD (CC), EPHX1 (high activity)
Různé kombinace genotypů v souvislosti s poškozením DNA A=XPD (AA), XPG (GG), XPC (CC) B= XPD (CC), XPG (GC+CC), XPC (AA) vs. P=0.006 A B A B XPD (AA), EPHX1 (low activity) vs. XPD (CC), EPHX1 (high activity) P=0.016
, XPG (GG), XPC (CC) B= XPD (CC), XPG (GC+CC), XPC (AA) vs. P= A. B. A. B. XPD (AA), EPHX1 (low activity) vs. XPD (CC), EPHX1 (high activity) P=")
42
Působení různých exogenních faktorů v indukci DNA poškození
Block of strand elongation Exogenous factors Modified from A.D. Kligerman, Y. Hu / Chemico-Biological Interactions 166 (2007) Pavel Vodicka
 Pavel Vodicka.")
43
CHA: control and exposed populations Frequency of chromosomal damage
Healthy controls (N = 751) compared with healthy exposed subjects (N = 1028)
 compared with healthy exposed subjects (N = 1028)")
44
CHA: control and exposed population - distribution
Distribution of frequencies of CAs (N = 1779)
")
45
CHA: control and exposed population - genotypes

46
CHA: control and exposed population - confounders
Binary logistic regression models – chromosomal damage and genotype variants (adjusted odds ratio) CAs aOR 95% C.I. For OR P-value Smoking 1.36 1.05 – 1.76 0.018 Occupational exposure 1.42 1.08 – 1.87 0.013 EPHX1 high activity genotype 0.67 0.45 – 0.98 0.040 CTA 1.34 1.10 – 1.64 0.004 XPD23 k751Q (T/G) 0.65 0.49 – 0.86 0.003
 CAs. aOR. 95% C.I. For OR. P-value. Smoking – Occupational exposure – EPHX1 high activity genotype – CTA – XPD23 k751Q (T/G) –")
47
Cyclin D1 (CCND1) G870A genotyp, hlavní konfoundery a frekvence ChA.
Variable Persons Significance OR 95% C.I. for OR Lower Upper Age (continuous) 730 .10 1.01 .99 1.03 Sex (M/F) 370/361 .08 1.33 .97 1.82 Smoking (S/NS) 250/481 .34 1.18 .84 1.64 Exposed/Unexposed 553/172 .01 1.68 1.16 2.45 CCND1_GG 200 - 1.00 CCND1_GA 379 .36 .83 1.69 CCND1_AA 151 1.85 1.17 2.93
 Sex (M/F) 370/ Smoking (S/NS) 250/ Exposed/Unexposed. 553/ CCND1_GG CCND1_GA CCND1_AA")
48
Frekvence chromozomálních aberací (%) a CCND1 genotyp
 a CCND1 genotyp")
49
BER kapacita a XRCC1 polymorphismus u zdravých osob
Pavel Vodicka,

50
Results from functional studies:
In a healthy population (n=244) associations between DNA repair polymorphisms and individual DNA repair capacity have been investigated. Significant associations have been found for XRCC1 Arg399Gln and hOGG1 Ser326Cys and for different BER polymorphisms in combination. A significant decrease in the capacity to repair DNA oxidative damage has been associated with variant alleles in hOGG1 Ser326Cys and APE1 Asn148Glu in combination (P=0.018). OGG1 X APE1 8-OH-dG OGG1 Ser326Cys genotype Base Excision Repair : (Modified from Friedberg, 2004) Vodicka et al,Carcinogenesis 2007
 associations between DNA repair polymorphisms and individual DNA repair capacity have been investigated. Significant associations have been found for XRCC1 Arg399Gln and hOGG1 Ser326Cys and for different BER polymorphisms in combination. A significant decrease in the capacity to repair DNA oxidative damage has been associated with variant alleles in hOGG1 Ser326Cys and APE1 Asn148Glu in combination (P=0.018). OGG1. X. APE1. 8-OH-dG. OGG1 Ser326Cys. genotype. Base Excision Repair : (Modified from Friedberg, 2004) Vodicka et al,Carcinogenesis")
51
(Luna et al., Nucleic Acids Res. 2005)
Vztah mezi 8-hydroxy-2´-deoxyguanosine adduckty v lymfocytech a hOGG1 Ser326Cys polymorfismem P=0.080 P=0.006 Variantní alela Ser souvisí se sníženou funkcí oxoguanine glycosylasy 1 (Luna et al., Nucleic Acids Res. 2005) Pavel Vodicka,
 Pavel Vodicka,")
52
Influence of genetic variability on DNA repair
Incision (tail DNA%) Incision (tail DNA%) We have observed a large inter-individual variability in both BER and NER pathway. This variability was determined by the polymorphism in OGG1 gene for BER, and slightly modulated by polymorphism in XPA genes for NER. These findings seems to be biologically relevant. Our findings obtained by phenotypic assay, are supported by some in vitro studies that also showed the decreased activity of enzyme when variant allele is present, in silico analysis showed that SNPs are located in functional sequence of the protein and some association studies showed relation to cancer risk. Slyskova J et al, Environ Mol Mutagen, 2011; Vodicka P et al, Carcinogenesis, 2007
 Incision (tail DNA%) We have observed a large inter-individual variability in both BER and NER pathway. This variability was determined by the polymorphism in OGG1 gene for BER, and slightly modulated by polymorphism in XPA genes for NER. These findings seems to be biologically relevant. Our findings obtained by phenotypic assay, are supported by some in vitro studies that also showed the decreased activity of enzyme when variant allele is present, in silico analysis showed that SNPs are located in functional sequence of the protein and some association studies showed relation to cancer risk. Slyskova J et al, Environ Mol Mutagen, 2011; Vodicka P et al, Carcinogenesis,")
53
Influence of biological and lifestyle factors on DNA repair
340 healthy individuals : 180 women / 160 men with similar age distribution : 150 food items + 12 antioxidants measured in plasma Oxidative lesions + strand breaks Beta t p Sex -.122 -2.168 .031 Fruit intake -.130 -2.295 .022 XPG Asn1104His .104 1.845 .049 NER Finally, we wanted to investigate the influence of diet on DNA damage and DNA repair. We have sampled 340 individuals that were controlled for diet and several antioxidants in their plasma. We observed that 2 most important factors that mostly influence DNA damage and reapir are sex and fruit intake. Higher consumption of antioxidant-rich diet was associated with lower damage and higher repair. Beta t p Sex .176 3.051 .002 Fruit intake .123 2.137 .033 Slyskova J et al, DNA Repair, accepted

54
Examples from humans: DNA damage and DNA repair according to exposure, Ist sampling

55
DNA repair kinetic: a lesson from animal study 2
γ-irradiation specific DNA repair activity in liver cells of mice, during and after the period of inhalation of 1,3-butadiene, and of unexposed control mice at particular time intervals The DNA repair activity in all animals is calculated as a difference between the initial levels of SSBs, measured immediately after γ-irradiation, and the levels of SSBs detected after 40 min. of incubation. Pavel Vodicka

56
Examples from humans: DNA repair rates in relation to exposure and smoking habit
Butadiene exposure In general healthy population: DNA repair capacity vs smoking R=0.168, P=0.006

57
Clonal nature of tumors – from a single cell
CARCINOGENESIS =multistep process – genetic + environmental factors Multiple mutations (growth controlling genes) Multiple causes and mechanisms Environmental factors: chemical carcinogens UV, ionizing radiation tumor viruses – RNA, DNA viruses Life-style Mutations – role in initiation of carcinogenesis
 Multiple causes and mechanisms. Environmental factors: chemical carcinogens. UV, ionizing radiation. tumor viruses – RNA, DNA viruses. Life-style. Mutations – role in initiation of carcinogenesis.")
58
Tentative multistep carcinogenesis process, as designed for CRC,
and potential points for biomarker recruitment General concept adopted and modified from Gareca et al., (2003) Eur. J. Cancer 39, Accumulation of DNA damage Attempt for DNA replication Genetic DNA mutations Signalling pathways Increased proliferation CRC Clinical presentation Polyps number& histology of polyps Adenoma formation TS, oncogenes APC, K-ras, p53 XME CYP, GST Grade of displasia Dysplasia DNA repair enzymes DNA adducts catenin COX-2 Invasive cancer Pavel Vodicka
 Eur. J. Cancer 39, Accumulation of DNA damage. Attempt for DNA replication. Genetic DNA mutations. Signalling pathways. Increased proliferation. CRC Clinical presentation. Polyps number& histology of polyps. Adenoma formation. TS, oncogenes. APC, K-ras, p53. XME. CYP, GST. Grade of displasia. Dysplasia. DNA repair enzymes. DNA. adducts. catenin. COX-2. Invasive cancer. Pavel Vodicka.")
59
Genetické změny v progresi CRC

63
Knudson´s two-hit hypothesis of loss of function of tumor suppressor gene
Journal.cambridge.com

64
Genomic instability The failure to transmit an accurate copy of the entire genome from one cell to its two daughter cells. Genomic instability can be divided into 2 categories Chromosomal instability (CIN) Microsatellite instability CIN has been linked to aneuploidy and CAs, LOH Cancer cells frequently exhibit CAs as translocations, deletions, or gene amplification – an importatnt source of chromosomal instability CINs are related to abnormalities of DNA metabolism, DNA repair, cell cycle control, apoptosis
 Microsatellite instability. CIN has been linked to aneuploidy and CAs, LOH. Cancer cells frequently exhibit CAs as translocations, deletions, or gene amplification – an importatnt source of chromosomal instability. CINs are related to abnormalities of DNA metabolism, DNA repair, cell cycle control, apoptosis.")
65
Sources of genomic instability
Nucleotide-excision instability (NIN): Loss of nucleotide excision repair activity Microsatellite instability (MSI): Deficiency of mismatch repair system Chromosomal instability (CIN): Aneuploidy Translocations Insertions Deletions Amplifications Mutator phenotype Loss of cell cycle control (promoting CIN) Defects in mitotic apparatus (propagating CIN) Defects in DNA repair (inducing CIN) Pavel Vodicka
: Loss of nucleotide excision repair activity. Microsatellite instability (MSI): Deficiency of. mismatch repair system. Chromosomal instability (CIN): Aneuploidy. Translocations. Insertions. Deletions. Amplifications. Mutator phenotype. Loss of cell cycle control. (promoting CIN) Defects in mitotic apparatus. (propagating CIN) Defects in DNA repair. (inducing CIN) Pavel Vodicka")
66
From H. Rajagopalan, C. Lengauer / Nature 432 (2004) 338-341
Some processes implicated in the process of aneuploidy Multiple roads to aneuploidy. Te scematic illustrates a simplified cell cycle, ilitin processes tat ave been implicated in te advert of aneupoidy. Several patways witin te cell cycle (indicated in red) can be disrupted. Enes (indicated in reen) associated wit tese processes and structures ave been found to be mutated or functionally altered in aneuploid cancers. From H. Rajagopalan, C. Lengauer / Nature 432 (2004) Pavel Vodicka
 can be disrupted. Enes (indicated in reen) associated wit tese processes and structures ave been found to be mutated or functionally altered in aneuploid cancers. From H. Rajagopalan, C. Lengauer / Nature 432 (2004) Pavel Vodicka.")
67
CIN Presence of multiple structural or numerical chromosome changes in tumour cells, in practice, often inferred from finding aneuploidy and/or polyploidy It is believed that CIN has a genetic basis The molecular basis of CIN has remained mysterious (many mechanisms have been postulated to be responsible for CIN) Mutations in the mitotic spindle checkpoint; the genes that ensure the proper segregation of duplicated chromosomes Tetraploidization of the genome caused by endoreduplication followed by loss of chromosomes due to instability of tetraploid genome Mutations in the process of duplication of the centrosome Formation of double strand breaks also generates CA Impaired replication fork progression and increased DNA replication stress – CIN in colorectal cancer cell lines; additonally, new CIN-suppressor genes identified (MCD 4/PIGN, MEX3C and KIAA0222) Burrell at al.,Nature, 494, 2013,
 Mutations in the mitotic spindle checkpoint; the genes that ensure the proper segregation of duplicated chromosomes. Tetraploidization of the genome caused by endoreduplication followed by loss of chromosomes due to instability of tetraploid genome. Mutations in the process of duplication of the centrosome. Formation of double strand breaks also generates CA. Impaired replication fork progression and increased DNA replication stress – CIN in colorectal cancer cell lines; additonally, new CIN-suppressor genes identified (MCD 4/PIGN, MEX3C and KIAA0222) Burrell at al.,Nature, 494, 2013,")
68
CHROMOSOMAL DAMAGE ACCORDING TO THE TYPE OF MALIGNANCY
-% of aberrant cells (ACs), chromosomal aberrations (CAs), chromatid type aberrations (CTA), chromosom type aberrations (CSA) Vodicka et al., Carcinogenesis 2010 Pavel Vodicka
, chromosomal aberrations (CAs), chromatid type aberrations (CTA), chromosom type aberrations (CSA) Vodicka et al., Carcinogenesis Pavel Vodicka.")
69
Chromosomal aberrations and cancer risk - distributions
Distribution of frequencies of ACs

70
Distribution of frequencies of CAs (N = 1780)
CHA: control and exposed population - distributions Distribution of frequencies of CAs (N = 1780)
")
71
TNM and Grading characteristics in solid tumors
TNM I +II vs. TMN III + IV CRC Lung cancer Breast cancer Test P-value Percentage of ACs in relation to the TNM categories Mann-Whitney U test 0.649 0.172 0.773 Percentage of CAs in relation to the TNM categories 0.689 0.214 0.966 Percentage of CTAin relation to the TNM categories 0.759 0.348 0.817 Percentage of CSA in relation to the TNM categoriesis 0.428 0.645 0.444 The significance level at 0.05 Grading G+G2 vs. G3+G4 CRC Lung cancer Breast cancer Null Hypothesis Test P-value Percentage of ACs in relation to the categories of grading Mann-Whitney U test 0.701 0.500 0.219 Percentage of CAs in relation to the categories of grading 0.684 0.249 Percentage of CTA in relation to the categories of grading 0.978 0.833 0.174 Percentage of CSA in relation to the categories of grading 0.940 0.333 0.426 The significance level at 0.05

72
Tom van Wezel, Mutagenesis,
2012

73
Repair Pathways

74
Schematic representation of the DNA repair and Growth arrest signaling pathways. Examples of the molecule known to act on the regulatory pathways are shown

75
Implication of DNA repair modulations in cancer
Implication of DNA repair modulations in cancer. DNA repair downregulation can contribute to genomic instability, which promotes malignant transformation of cells, and leads to cellular sensitivity to DNA damaging therapy. DNA repair upregulation can contribute to genomic stability, which lead to acquired resistance to the DNA damaging therapy.

76
Bleomycine Challenge Assay – DSB Repair

77
Bleomycin challenge

78
Mismatch repair pathway
The DNA mismatch repair (MMR) system plays a key role in maintenance of genomic stability. MMR edits mismatches and insertions/deletions generated during DNA replication and mitotic/meiotic recombination. The components of MMR machinery are also involved in DNA damage response, cell cycle, apoptosis, chromatin remodeling and antibody diversity. MMR genes are of importance as „high penetrance“ genes in the aetiology of familial CRC (HNPCC syndrome) and a set of sporadic cancers. MMR disfunction leads to gross instability in microsatellite sequences throughout genome (MSI). A limited knowledge has been available on the role of MMR genes as „low-penetrance“ genes in sporadic CRC risk
 system plays a key role in maintenance of genomic stability. MMR edits mismatches and insertions/deletions generated during DNA replication and mitotic/meiotic recombination. The components of MMR machinery are also involved in DNA damage response, cell cycle, apoptosis, chromatin remodeling and antibody diversity. MMR genes are of importance as „high penetrance genes in the aetiology of familial CRC (HNPCC syndrome) and a set of sporadic cancers. MMR disfunction leads to gross instability in microsatellite sequences throughout genome (MSI). A limited knowledge has been available on the role of MMR genes as „low-penetrance genes in sporadic CRC risk.")
80
CpG island methylation of the hMLH1 promoter region vs MLH1 expression by IHC
We analyzed colon biopsies of 105 CRC patients We found hypermethylation in 8 tumors with MSI-H status. Hypermethylated tumors localized in right colon only (4x caecum C18.0. and 4x ascendens C18.2) There is a strong association between promoter metylation of hMLH1 and reduced protein expression (R=-0.75, P<0.0001). Crc7, 29 and 35 is MSI H 80
 There is a strong association between promoter metylation of hMLH1 and reduced protein expression (R=-0.75, P<0.0001). Crc7, 29 and 35 is MSI H. 80.")
81
Expression of MMR genes
mRNA expression levels of MMR genes in tumor and healthy tissues in the study subjects -1.16 fold lower mRNA levels of EXO1 gene were observed in tumor tissues when compared with adjacent healthy mucosa significantly higher expression levels of MSH3 gene were observed in colon tumors when compared to adjacent healthy mucosa; 1.18 fold change

82
Fold change differences in expression level of MMR genes in colon tumors relative to rectal tumors

83
Ovarial cancer

84
Dlouhé delece – předbežné výsledky
LOH (exony) predbezne
 predbezne.")
85
Sumární změny v MLH1 genu:
MLH1- loss of the exon MLH1+ gain of the exon

86
Dráhy DNA oprav DNA repair is a complex system of defenses evolved to protect the genomic integrity and involved in the process preventing carcinogenesis. Interindividual differences in DNA repair capacities are important determinants of cancer risk including CRC. CRC represents a complex disease where susceptibility may be influenced by genetic polymorphisms in the DNA repair system.

87
FOCUSING ON DNA REPAIR CAPACITY in CANCER PATIENTS
While exposure in healthy subjects may result in compensatory increase in DNA excision repair, Individuals with defective DNA repair are at severely increased risk of developing cancer and other diseases What do we measure by applying functional repair assays in surrogate (blood) cells and in target (tumor) tissues?
 cells and in target (tumor) tissues")
88
DNA repair capacity is multifactorial biologival process
NER-DRC phenotype Transcript Proteín > ERCC1 mRNA XPA XPB XPC XPD XPE XPF XPG Gene SNP 6 SNPs 9 SNPs 8 SNPs 15 SNPs 13 SNPs 14 SNPs 7 SNPs Precursors, availability Epigenetic regulation Transcription regulation Enzyme activation Enzymme inhibition External exposure Nutrition < Biological factors Other interfering proteins Environmental variability Mechanism Source Genetic variability Life style DNA reparacna kapacita je multifaktorialny proces, z genetickou zlozkou a environmentalnou zlozkou. Tu je ukazka toho, od akych moznych faktorov moze zavisiet kapacita NER drahy. NER-DRC je urceny aktivitou mnohych genov, ktore su vsetky v populacii polymorfne a funkcny dopad tychto polymorfizmov zatial nie je dostatocne preskumany. Na druhej strane su to externe faktory, ako strava, expozicia, zivotny styl mozu vplyvat na aktivitu proteinov, cez epigeneticku regulaciu, regulaciu transkripcie, translacie ci aktivity proteinu. Meranie DNA reparacnej kapacity teda predstavuje velmi komplexny pristup, ktory zahrna vsetky tieto faktory.

89
DNA DAMAGE IN INCIDENT CRC PATIENTS VS CONTROLS

90
NER CAPACITY IN INCIDENT SPORADIC CRC PATIENTS

91
NER CAPACITY IN CRC PATIENTS STRATIFIED FOR STAGING
OF THE TUMOR 14 15 24

92
NER GENE EXPRESSION LEVELS IN CRC PATIENTS AND CONTROLS

93
A D C B BER RATES IN VARIOUS TISSUES of CRC PATIENTS

94
NER RATES IN VARIOUS TISSUES of CRC PATIENTS
D C B NER RATES IN VARIOUS TISSUES of CRC PATIENTS

95
Expression profile of the genes involved in excision repair
Pathway Fold change p-value NEIL1 BER -1.26 0.0004 APEX1 1.15 0.0138 OGG1 -1.15 0.0467 PARP1 1.12 0.0210 LIG3 1.08 0.0922 MUTYH -1.08 0.4303 MPG -1.05 0.5807 NTHL1 1.00 0.9551 CSB NER -1.28 0.0002 CCNH -1.25 0.0081 XPA 0.0190 XPD 0.0408 LIG1 1.16 0.0708 MNAT1 0.1128 RPA3 0.1739 CDK7 1.07 0.4839 XPF 1.06 0.4089 RPA2 0.4898 XPB 1.05 0.3349 DDB1 1.02 0.6063 ERCC1 0.6803 DDB2 -1.03 0.6201 RAD23B 0.6661

96
DNA repair in relation to therapy
39 CRC patients 48 controls Peripheral blood ~ 64 years ~ 64 years Surgery Chemotherapy T0 T1 T2 6 months CRC patients at diagnosis, then another 6 months later months when they have undergone surgery and chemotherapy, and another six months when they were all considered to be cured, with no relapse. And we compared DNA repair.. Slyskova J et al, Molecular Carcinogenesis, in press

97
p=0.009 p=0.001 N=38 N=33 p=0.038 p=0.032 N=38 N=33

98
DNA repair in relation to therapy
Controls 40 DNA repair genes → + T2 CRC T0 + T1 Slyskova J et al, Molecular Carcinogenesis, in press 98

99
Introduction NER repair BER repair Oxaliplatin 5-fluorouracil
Source: Clinical Colorectal Cancer CIG Media LP 5-fluorouracil BER repair Source: Longley D. Natu 99

100
? Questions to be addressed
Mechanism? Cancer temporary suppression of DNA repair WHERE? Target, surrogate? tail DNA % NER Higher repair = Poor responders Worse prognosis Milder side effects Based on these results, there is a theory that there is a cancer-related temporary suppression of DNA repair that is reverted back to normal stage during the remission period. However, there still is certain inter-individual variability in repair capacity between patients. It will be interesting to explore further, whether this variability might have an effect on the treatment efficacy and long-term survival. We could not answer these questions in this study, since all patients that left in the study for whole year were good responders with no relapse of the disease within a year of follow-up. ? Lower repair = Good responders Better prognosis Advanced side effects

101
Department of Molecular Biology of Cancer
ACKNOWLEDGEMENTS I would like to express my deep gratitude to all co-authors, particularly for their friendship and valuable contribution Prague (CZ) P. Vodicka V. Vymetalkova L. Vodickova J. Slyskova P. Procházka M. Svoboda L.Bielik, M. Kroupa A. Rejhová S. Vodenkova K. Jiraskova Collaborating clinicians M. Schneiderová L. Lipská, M. Levý T. Büchler Pilsen (CZ) Václav Liška and collegues Hradec Kralove (CZ) Rudolf Stetina London, Oxford (UK) Ian Tomlinson Richard Houlston Pisa and Torino (I) Stefano Landi Federica Gemignani Barbara Pardini Alessio Naccarati Heidelberg (D) Kari Hemminki Asta Foersti Rajiv Kumar Justo L. Bermejo Federico Canzian Daniele Campa Öröbro, Sweden Torbjorn Nilsson Department of Molecular Biology of Cancer Institute of Experimental Medicine, Academy of Sciences of Czech Republic Website: This work is supported by GACR P304/10/1286, P304/12/1585 and IGA NR
 P. Vodicka. V. Vymetalkova. L. Vodickova. J. Slyskova. P. Procházka. M. Svoboda. L.Bielik, M. Kroupa. A. Rejhová. S. Vodenkova. K. Jiraskova. Collaborating clinicians. M. Schneiderová. L. Lipská, M. Levý. T. Büchler. Pilsen (CZ) Václav Liška and collegues. Hradec Kralove (CZ) Rudolf Stetina. London, Oxford (UK) Ian Tomlinson. Richard Houlston. Pisa and Torino (I) Stefano Landi. Federica Gemignani. Barbara Pardini. Alessio Naccarati. Heidelberg (D) Kari Hemminki. Asta Foersti. Rajiv Kumar. Justo L. Bermejo. Federico Canzian. Daniele Campa. Öröbro, Sweden. Torbjorn Nilsson. Department of Molecular Biology of Cancer. Institute of Experimental Medicine, Academy of Sciences of Czech Republic. Website: This work is supported by GACR P304/10/1286, P304/12/1585 and IGA NR")
102
Děkuji Vám za pozornost

103
Cytogenetic analysis in patients with newly diagnosed colorectal and breast cancer and healthy controls – results CRC Controls (n=298) Patients (n=100) P-value Age (years) 56,9±13,5 63,0±10,2 <0,0001 Aberrant cells (%) 1,82±1,32 2,14±1,44(*) 0,056 Total aberrations (%) 1,95±1,47 2,27±1,65(*) 0,087 CTA (%) 1,11±0,99 1,45±1,28* 0,030 CSA (%) 0,84±1,13 0,82±1,01 0,838 BREAST Controls (n=123) Patients (n=123) P-value Age (years) 65,2±16,2 59,7±10,2 0,015 Aberrant cells (%) 1,98±1,41 2,52±1,53** 0,002 Total aberrations (%) 2,08±1,52 2,64±1,60** CTA (%) 1,23±1,09 1,54±1,33(*) 0,082 CSA (%) 0,85±0,99 1,11±0,99* 0,023 statistically significant difference between patients and controls *p≤0,05 (*) the borderline of statistical significance statistically significant difference between patients and controls *p≤0,05, **p≤0,01 (*) the borderline of statistical significance
 Patients (n=100) P-value. Age (years) 56,9±13,5. 63,0±10,2. <0,0001. Aberrant cells (%) 1,82±1,32. 2,14±1,44(*) 0,056. Total aberrations (%) 1,95±1,47. 2,27±1,65(*) 0,087. CTA (%) 1,11±0,99. 1,45±1,28* 0,030. CSA (%) 0,84±1,13. 0,82±1,01. 0,838. BREAST. Controls (n=123) Patients (n=123) P-value. Age (years) 65,2±16,2. 59,7±10,2. 0,015. Aberrant cells (%) 1,98±1,41. 2,52±1,53** 0,002. Total aberrations (%) 2,08±1,52. 2,64±1,60** CTA (%) 1,23±1,09. 1,54±1,33(*) 0,082. CSA (%) 0,85±0,99. 1,11±0,99* 0,023. statistically significant difference between patients and controls *p≤0,05. (*) the borderline of statistical significance. statistically significant difference between patients and controls *p≤0,05, **p≤0,01. (*) the borderline of statistical significance.")
104
Chromosomal damage (the percentage of aberrant cells, total aberrations, CTA and CSA) in patients with newly diagnosed CRC and breast cancer and healthy controls
 in patients with newly diagnosed CRC and breast cancer and healthy controls")
105
Distribution of the frequency of aberrant cells in patients with newly diagnosed colorectal and breast cancer and healthy controls (expressed in %) 0 – 1% aberrant cells low frequency of aberrant cells 2% aberrant cells middle frequency of aberrant cells 3 a ≥4% aberrant cells high frequency of aberrant cells

106
Binary logistic regression analysis – determination of the cancer risk influenced by chromosomal damage and other factors (age and smoking) – aOR CRC Breast cancer 100 patients vs. 298 controls aOR 95%CI P-value Aberrant cells (%) 1,156 0, ,371 0,097 Age (years) 1,038 1, ,058 ≤0,0001 Smoking 1,065 1, ,131 0,038 Total aberrations (%) 1,117 0, ,298 0,149 1,066 1, ,132 0,035 CTA (%) 1,295 1, ,596 0,016 1,039 1, ,058 1,063 1, ,129 0,047 CSA (%) 0,952 0, ,184 0,656 1,040 1, ,059 1,067 1, ,132 0,034 123 patients vs. 123 controls aOR 95%CI P-value Aberrant cells (%) 1,335 1, ,600 0, 002 Age (years) 0,969 0, ,988 0,002 Smoking 1,104 1, ,198 0,018 Total aberrations (%) 1,315 1, ,562 0,968 0, ,987 0,001 1,102 1, ,196 0,020 CTA (%) 1,194 0, ,480 0,106 0,973 0, ,992 0,005 1,096 1, ,189 0,028 CSA (%) 1,567 1, ,083 0,964 0, ,984 ≤0,0001 1,118 1, ,215 0,008
 1,156. 0, ,371. 0,097. Age (years) 1,038. 1, ,058. ≤0,0001. Smoking. 1,065. 1, ,131. 0,038. Total aberrations (%) 1,117. 0, ,298. 0,149. 1,066. 1, ,132. 0,035. CTA (%) 1,295. 1, ,596. 0,016. 1,039. 1, ,058. 1,063. 1, ,129. 0,047. CSA (%) 0,952. 0, ,184. 0,656. 1,040. 1, ,059. 1,067. 1, ,132. 0, patients vs. 123 controls. aOR. 95%CI. P-value. Aberrant cells (%) 1,335. 1, ,600. 0, 002. Age (years) 0,969. 0, ,988. 0,002. Smoking. 1,104. 1, ,198. 0,018. Total aberrations (%) 1,315. 1, ,562. 0,968. 0, ,987. 0,001. 1,102. 1, ,196. 0,020. CTA (%) 1,194. 0, ,480. 0,106. 0,973. 0, ,992. 0,005. 1,096. 1, ,189. 0,028. CSA (%) 1,567. 1, ,083. 0,964. 0, ,984. ≤0, ,118. 1, ,215. 0,008.")
107
Opravy chybného párování bází řízené metylací
Opravy chybného párování bází řízené metylací (mismatch repair, MMR) hrají klíčovou roli v zachování genomické stability. MMR opravuje chyby v replikaci DNA, zahrnující nesprávně zařazané nukleotidy, inzerce a delece. A dále chyby v průběhu mitotické nebo meiotické rekombinace. Složky systému MMR jsou také zahrnuty v odpovědi na poškození DNA, buněčném cyklu, apoptóze a remodelaci chromatinu. Geny MMR jsou důležité jak v etiologii familiárního CRC, tak v souboru sporadicky se vyskytujících nádorů. Dysfunkce MMR vede k nárůstu nestability v mikrosatelitních sekvencích (MSI) napříč genomem.
 hrají klíčovou roli v zachování genomické stability. MMR opravuje chyby v replikaci DNA, zahrnující nesprávně zařazané nukleotidy, inzerce a delece. A dále chyby v průběhu mitotické nebo meiotické rekombinace. Složky systému MMR jsou také zahrnuty v odpovědi na poškození DNA, buněčném cyklu, apoptóze a remodelaci chromatinu. Geny MMR jsou důležité jak v etiologii familiárního CRC, tak v souboru sporadicky se vyskytujících nádorů. Dysfunkce MMR vede k nárůstu nestability v mikrosatelitních sekvencích (MSI) napříč genomem.")
108
Metylation of MLH1 in CRC patients
Crc7, 29 and 35 is MSI H 12 CRC samples showed higher level of methylation than 2.5% When setting „cutoff“ for methylation to 15% – strong association with MSI-H 7 healthy tissues showed methylation higher than 2.5% - often in pairs with methylated CRC sample 108

110
MLH1 protein expression - colon ascendens and flexura hepatica
[%] 110

111
MLH1 protein expression - colon ascendant and flexura hepatica
[%] 111

112
Expression of MLH1 in CRC patients
There is a association between promoter metylation of MLH1 and protein expression of MLH1 R = R2 = 0.52 P = Primes nenad bb Micro RNA regulace-uplatneni (48% efektu?) 112
 112.")
113
Long deletions – preliminary results
LOH (exony) predbezne
 predbezne.")
114
All changes in MLH1 gene:
MLH1- loss of the exon MLH1+ gain of the exon

115
DNA repair pathway DNA repair is a complex system of defenses evolved to protect the genomic integrity and involved in the process preventing carcinogenesis. Interindividual differences in DNA repair capacities are important determinants of cancer risk including CRC. CRC represents a complex disease where susceptibility may be influenced by genetic polymorphisms in the DNA repair system.

116
DNA damage in controls and CRC incident patients at the time
of diagnosis P < 0.001 N=69

117
P < 0.001 N=69 NER capacity in controls and CRC incident patients at the time of diagnosis

118
NER capacity in CRC patients stratified for staging of the tumor
14 15 24
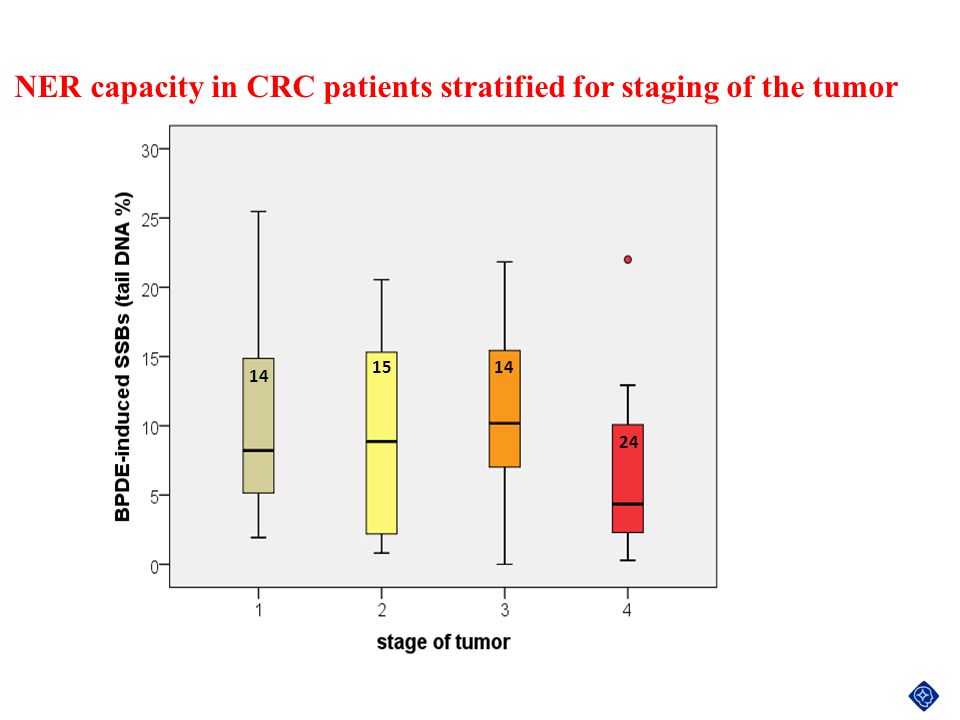
119
NER gene expression levels in CRC patients and controls

120
BER hladiny v různých tkáních CRC pacientů

121
NER hladiny v různých tkáních CRC pacientů

122
Vztah mezi NER a BER hodnotami
v nádorové tkáni R=0.32 p=0.007 R2= 0.10

123
Survival or Apoptosis, that’s the problem in cancer therapy and for individual health. The
determination either survival or apoptosis is due to the balance between DNA damage and the DNA repair levels in cells.

124
Děkuji Vám za pozornost

125
Arguments for genomic instability as the engine of tumorgenesis
Tumors harbour too many mutations to be explained by anything other than underlying genomic instability The probability of a tumour acquiring enough mutations for the full, malignant phenotype is too low unless the cells have an unstable genome Humans and model organisms with inherent genomic instability are prone to tumors In some tumours, there is direct evidence that some pathways that are involved in maintaining genomic integrity are defective

126
the studies ON MULTIPLE LOW-RISK VARIANTS
Sporadic forms of cancer are hallmarked by multigenic features in a complex interaction with environmental/life-style factors To assess the CRC risk in association with polymorphisms, and haplotypes in several pathways to identify relevant candidate genes. To participate on the alternative approach to identify relevant susceptibility loci using GWAS. To verify and refine GWAS (post-GWAS fine-mapping) data by candidate gene meta-analyses and by next generation sequencing Definition of both genetic and phenotypic landscape of CRC (e.g. DNA repair functional tests)
 data by candidate gene meta-analyses and by next generation sequencing. Definition of both genetic and phenotypic landscape of CRC (e.g. DNA repair functional tests)")
127
Simplified scheme showing a putative effect of low penetrance variants on CRC susceptibility
Individual Susceptibility 1 SNP / 2SNPs/…. Haplotypes Gene 1 1 SNP / 2SNPs/…. Haplotypes Gene 2 1 SNP / 2SNPs/…. Haplotypes Gene 3 1 SNP / 2SNPs/…. Haplotypes ……… 1 SNP / 2SNPs/…. Haplotypes Gene n +/- +/- +/- +/- +/- Internal environmental influence External environmental influence NER, BER, DSB, MMR, Rev. Repair Pathway 1 DNA Repair Pathway 3 Metabolism Pathway 2 Cell cycle Pathway 4 Immune response Other Pathways Cancer Risk

128
TP53 haplotype analysis Polakova et al., Human Mut 2009
TP53 Ins 11951_11966 Ex4 +119G>C IVS7 +72C>T Ex G>A Controls CRC OR (95% CI) A1 G C 763 819 Referent A2 106 159 1.40 ( ) 141 90 0.60 ( ) A 51 54 0.99 ( ) 63 36 0.53 ( ) T 32 34 0.99 ( ) 29 10 0.32 ( ) 24 5 0.19 ( ) Global P –value for haplotype effect <0.0001 Significant ORs for individual haplotypes are in bold (Yellow= increased risk, Blue= decreased risk) Polakova et al., Human Mut 2009
 A1. G. C Referent. A ( ) ( ) A ( ) ( ) T ( ) ( ) ( ) Global P –value for haplotype effect < Significant ORs for individual haplotypes are in bold (Yellow= increased risk, Blue= decreased risk) Polakova et al., Human Mut")
129
16 CRC susceptibility loci identified by GWAS
8q24.21 (rs ), 18q21 (rs , rs and rs ), 15q13.3 (rs and rs10318), 10p14 (rs ), 8q23.3 (rs )1 1q41 (rs , rs ), 3q26.2 (rs ), 12q13.13 (rs , rs ) and 20q13.33 (rs ), 14q22.2 (rs ), 16q22.1 (rs ), 19q13.11 (rs ) and 20p12.3 (rs and rs )2 11q23.1 (rs )3 1Tomlinson et al., Nature Genetics 2008 2Houlston et al., Nature Genetics 2010 3Pittman et al., Human Molecular Genetics 2008
, 18q21 (rs , rs and rs ), 15q13.3 (rs and rs10318), 10p14 (rs ), 8q23.3 (rs )1. 1q41 (rs , rs ), 3q26.2 (rs ), 12q13.13 (rs , rs ) and 20q13.33 (rs ), 14q22.2 (rs ), 16q22.1 (rs ), 19q13.11 (rs ) and 20p12.3 (rs and rs )2. 11q23.1 (rs )3. 1Tomlinson et al., Nature Genetics Houlston et al., Nature Genetics Pittman et al., Human Molecular Genetics")
130
SNPs at 16 genetic loci associated with CRC risk
rs * (1q41), rs (3q26.2), rs (8q23.3), rs (8q24.21), rs (10p14), rs * (11q23.1), rs (12q13.13), rs #, rs (14q22.2), rs † (15q13.3), rs (16q22.1), rs (18q21.1), rs (19q13.11), rs961253† and rs † (20p12.3) and rs # (20q13.33). Statisticaly significant associations with specific phenotypes are highlighted *microsatelite stable rectal disease †microsatelite stable colonic disease #microsatelite instability colonic disease Lubbe et al., Carcinogenesis 2011
, rs (3q26.2), rs (8q23.3), rs (8q24.21), rs (10p14), rs * (11q23.1), rs (12q13.13), rs #, rs (14q22.2), rs † (15q13.3), rs (16q22.1), rs (18q21.1), rs (19q13.11), rs961253† and rs † (20p12.3) and rs # (20q13.33). Statisticaly significant associations with specific phenotypes are highlighted. *microsatelite stable rectal disease. †microsatelite stable colonic disease. #microsatelite instability colonic disease. Lubbe et al., Carcinogenesis")
131
Gene promoter variant CASP N ins/del (rs ) verified on 13T European CRC cases and 14T controls: no association with sporadic cases (Peterlongo et al. In preparation) None of gene variants in APC D188V (rs459552), MLH1 I219V (rs ), MSH6 P92P (rs ) and D180D (rs ) and MUTYH V22M (rs ) and Q338H (rs ) was associated with sporadic CRC risk on the same population as above (Picelli et al. In preparation) Two pairs of tagging SNPs at 1q41 (rs and rs ) and 12q13.13 (rs and rs ) were associated with CRC. However, neither pair fully captured the association. The study shows intrinsic difficulties of post-GWAS fine-mapping studies (Spain et al. Human Molec Genetics, Epub Nov )
 None of gene variants in APC D188V (rs459552), MLH1 I219V (rs ), MSH6 P92P (rs ) and D180D (rs ) and MUTYH V22M (rs ) and Q338H (rs ) was associated with sporadic CRC risk on the same population as above (Picelli et al. In preparation) Two pairs of tagging SNPs at 1q41 (rs and rs ) and 12q13.13 (rs and rs ) were associated with CRC. However, neither pair fully captured the association. The study shows intrinsic difficulties of post-GWAS fine-mapping studies (Spain et al. Human Molec Genetics, Epub Nov )")
132
Recent research shows a pendulum swinging from GWAS to next generation candidate gene studies-capturing variants with low frequency and small effect Post-GWAS fine mapping-identification of candidate functional variants Meta-analyses of candidate functional variants Tomlinson et al. Mutagenesis 2012 in press Other approaches to identify functional consequences of relevant pathways (examples shown above) Slyskova et al. Mutagenesis 2012 in press
 Slyskova et al. Mutagenesis 2012 in press.")
133
Různé kombinace genotypů v souvislosti s chromosomálním poškozením
A= EPHX1 (high activity), GSTM1 (positive) vs. B= EPHX1 (low activity), GSTM1 (null) P=0.008 B A A= EPHX1 (high activity), GSTT1 (positive) vs. B= EPHX1 (low activity), GSTT1 (null) P=0.002 B A
, GSTM1 (positive) vs. B= EPHX1 (low activity), GSTM1 (null) P= B. A. A= EPHX1 (high activity), GSTT1 (positive) vs. B= EPHX1 (low activity), GSTT1 (null) P= B. A.")
134
A= XPD (AA), EPHX1 (low activity) B= XPD (CC), EPHX1 (high activity)
Různé kombinace genotypů v souvislosti s chromosomálním poškozením A=XPD (AA+AC), XPG (GG), XPC (CC), XRCC1 (GA+AA) B= XPD (CC), XPG (GC+CC), XPC (AA+AC), XRCC1 (GG+GA) * P=0.004 vs. A B A= XPD (AA), EPHX1 (low activity) vs. B= XPD (CC), EPHX1 (high activity) P=0.007 A B
, XPG (GG), XPC (CC), XRCC1 (GA+AA) B= XPD (CC), XPG (GC+CC), XPC (AA+AC), XRCC1 (GG+GA) * P= vs. A. B. A= XPD (AA), EPHX1 (low activity) vs. B= XPD (CC), EPHX1 (high activity) P= A. B.")
135
Associace mezi kapacitou opravy oxidativního DNA poškození, hOGG1 a APE1 genotypy
* A decrease in the capacity to repair DNA oxidative damage has been associated with variant alleles in hOGG1 Ser326Cys and APE1 Asn148Glu in combination (P=0.018). OGG1 Ser326Cys genotype Pavel Vodicka
. OGG1 Ser326Cys. genotype. Pavel Vodicka.")
136
Examples from humans: DNA damage and DNA repair according to exposure, Ist sampling

137
DNA repair kinetic: a lesson from animal study 2
γ-irradiation specific DNA repair activity in liver cells of mice, during and after the period of inhalation of 1,3-butadiene, and of unexposed control mice at particular time intervals The DNA repair activity in all animals is calculated as a difference between the initial levels of SSBs, measured immediately after γ-irradiation, and the levels of SSBs detected after 40 min. of incubation. Pavel Vodicka

138
Clonal nature of tumors – from a single cell
CARCINOGENESIS =multistep process – genetic + environmental factors Multiple mutations (growth controlling genes) Multiple causes and mechanisms Environmental factors: chemical carcinogens UV, ionizing radiation tumor viruses – RNA, DNA viruses Life-style Mutations – role in initiation of carcinogenesis
 Multiple causes and mechanisms. Environmental factors: chemical carcinogens. UV, ionizing radiation. tumor viruses – RNA, DNA viruses. Life-style. Mutations – role in initiation of carcinogenesis.")
Podobné prezentace










>")












